
亚热带主要造林树种土壤氮保留及相关功能的微生物特征*
2020-09-16 04:54梁艺凡杨军钱张冰冰施秀珍胡行伟黄志群
林业科学 2020年8期
王 磊 梁艺凡 杨军钱 张冰冰 王 涛 施秀珍 胡行伟 黄志群
(1.福建师范大学地理科学学院 福建师范大学湿润亚热带山地生态国家重点实验室培育基地 福州 350007)
全球许多地区氮沉降已超过植物和微生物的需求,出现氮饱和现象。我国亚热带地区是全球氮饱和最严重区域之一,引起国内外广泛关注(Moetal., 2008)。氮素流失将会导致水体富营养化(Howarthetal., 1988)、地下水污染(Peterjohnetal., 1996)、土壤酸化(Vitouseketal., 1997)、生态系统养分失衡(Schulzeetal., 1989)和全球气候变化(Huetal., 2015)等一系列问题。因此,如何减少我国亚热带森林土壤氮流失成为亟待解决的科学问题。
土壤氮循环过程(包括固氮、硝化和反硝化作用等)主要由相关功能微生物驱动。固氮作用是土壤固氮微生物将大气中N2转化成NH3的过程,对增加氮素输入、提高土壤肥力有重要作用。编码固氮酶nifH基因是研究固氮微生物最常用的标记物(Mirzaetal., 2014),但是目前对森林土壤中nifH基因丰度的研究还很缺乏(Levy-Boothetal., 2014)。氨氧化作用是土壤硝化作用的限速步骤,主要由氨氧化古菌(ammonia oxidizing archaea, AOA)和氨氧化细菌(ammonia oxidizing bacteria, AOB)驱动(贺纪正等, 2009)。反硝化作用是反硝化微生物将NO3--N依次还原为N2O或N2的过程(贺纪正等, 2013)。土壤硝酸盐淋溶及氮氧化物温室气体的排放是亚热带森林生态系统氮损失的主要途径(Zhangetal., 2014)。充分认识树种如何影响土壤固氮、硝化和反硝化微生物功能基因丰度及群落结构,对于提高土壤氮保留能力具有重要意义。
树种通过作用于多个氮循环过程影响土壤氮保留,不同树种土壤氮保留能力显著差异(Templaretal., 2005)。Morales等(2010)研究发现森林土壤比农业土壤中nifH基因丰度更高。Levy-Booth等(2010)发现nifH基因丰度与花旗松(Pseudotsugamenziesii)土壤有机碳含量显著正相关。森林土壤pH值、含水量、C/N、有机碳及黏粒含量是影响AOA和AOB生态位分化的重要非生物因子(Luetal., 2015;Shietal., 2016;Wessénetal., 2011)。李永春等(2014)发现由于杉木(Cunninghamialanceolata)根系分泌物及枯枝落叶分解导致土壤pH值降低,可能利于AOA生长。亚热带森林土壤的反硝化作用主要受氧化还原电位、土层深度、土壤有机碳含量等因素调控(Xuetal., 2007;Zhangetal., 2014)。Reverchon等(2015)发现柚木(Tectonagrandis)和白饭树(Flueggeavirosa)混栽,能显著增加编码硝酸盐还原酶narG、编码亚硝酸盐还原酶nirS和编码一氧化二氮还原酶nosZ基因的丰度;大多数反硝化基因丰度与土壤全氮含量、土壤C/N显著相关。
本文以亚热带5种人工纯林,闽楠(Phoebebournei)、火力楠(Micheliamacclurei)、木荷(Schimasuperba)、杉木和福建柏(Fokieniahodginsii)为对象,探讨主要造林树种对土壤氮循环相关功能微生物基因丰度和群落结构的影响,阐明树种对土壤氮保留影响的微生物驱动机制,为我国调整人工林树种、提高人工林生产力和可持续性、减少氮流失等提供科学依据。
1 研究区概况
样地位于福建厦门市汀溪国有防护林场(24°49′N,118°07′E),属南亚热带海洋性季风气候,光热充足、雨量丰沛、夏秋季节多台风,年均温20.6 ℃,年均降水量1 568 mm,降水主要发生在3—9月。主要地貌类型为低山丘陵,平均海拔202~390 m,土壤以酸性红壤为主。选取立地基本一致的闽楠、火力楠、木荷、杉木和福建柏5种人工林(基本情况见表1),其前茬树种分别为杉木、马尾松(Pinusmassoniana)、木荷、杉木和马尾松。5种人工林的林下植被相似,主要有铁线蕨(Adiantumcapillus-veneris)、乌毛蕨(Blechnumorientale)、藤黄檀(Dalbergiahancei)、寄生藤(Dendrotrophefrutescens)、酸叶胶藤(Ecdysantherarosea)、羊角藤(Morindaumbellata)、五节芒(Miscanthusfloridulus)、蒲公英(Taraxacummongolicum)、芒箕(GleichenialinearisClarke)、金毛狗(Cibotiumbarometz)等。
2 研究方法
2.1 样地设置
2017年11月,在5种人工林中各设置3个20 m×20 m的样地,共15个。同一树种林分的3个重复之间距离1~3 km。
2.2 土壤样品采集
2017年11月在各样地内,采用内径3.5 cm的土钻沿样地对角线上的10个点采集表层(0~10 cm)土样,采集点距离树干0.5 m以上,将同一样地内的土样混匀后装入无菌自封袋,放入保鲜盒,尽快运回实验室。在室内去除土样中的砂石、可见根和动物等,过2 mm孔径无菌筛网,充分混匀后分为3份: 1份储存在4 ℃冰箱备用,用于测定土壤微生物生物量、矿质氮等; 1份放入2 mL的无菌离心管中,储存在-80 ℃冰箱用于土壤微生物DNA提取和功能基因丰度测定; 1份在室温下自然风干,用于测定土壤基本理化性质。
2.2 土壤理化性质及微生物生物量
采用玻璃电极pH计测定土壤pH值,土水比1∶2.5。烘干法测定土壤质量含水量。2 mol·L-1KCL溶液浸提新鲜土样后,利用连续流动分析仪(Skalar San++,荷兰)测定矿质氮含量。元素分析仪(Elemental EL MAX CNS analyzer,德国)测定土壤全碳和全氮含量并计算碳氮比。环刀法测定0~10 cm土层土壤密度。英国马尔文仪器公司的MasterSize2000型激光粒度仪测土壤质地。氯仿熏蒸浸提法测定土壤微生物生物量(Vanceetal., 1987)。
2.3 土壤DNA提取及实时荧光定量PCR
土壤微生物DNA提取: 称取0.3 g土壤,使用MOBIO POWERSOIL土壤提取试剂盒(MOBIO laboratories, Carlsbad, California, USA),提取土壤总DNA。使用Nano Drop 2000荧光分光光度计测定土壤DNA,将符合要求的土壤DNA样品保存在-80 ℃冰箱中备用。土壤氮循环各功能基因的定量PCR引物参见表2。
2.4 末端限制性片段长度多态性(T-RFLP)
AOA的群落结构通过末端限制性片段多态性技术(T-RFLP)进行分析。AOB由于丰度较低,未能通过PCR扩增获得足量的PCR产物用于T-RFLP群落结构分析。AOA的扩增引物为CrenamoA23f/CrenamoA616 r。使用Bio-Rad公司的S-1000 PCR仪对土壤总DNA进行扩增。具体试验参照胡行伟(2013)的方法。
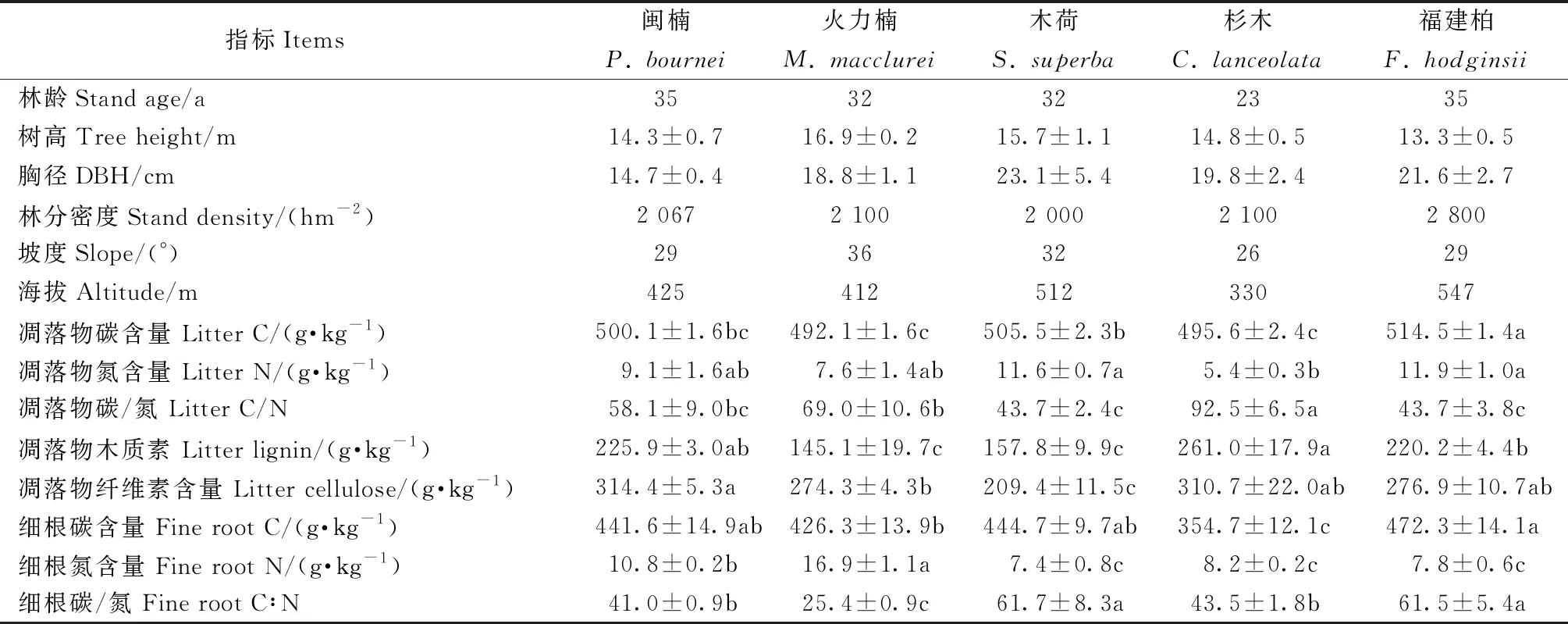
表1 不同树种人工林基本概况(均值±标准误,n=3)①
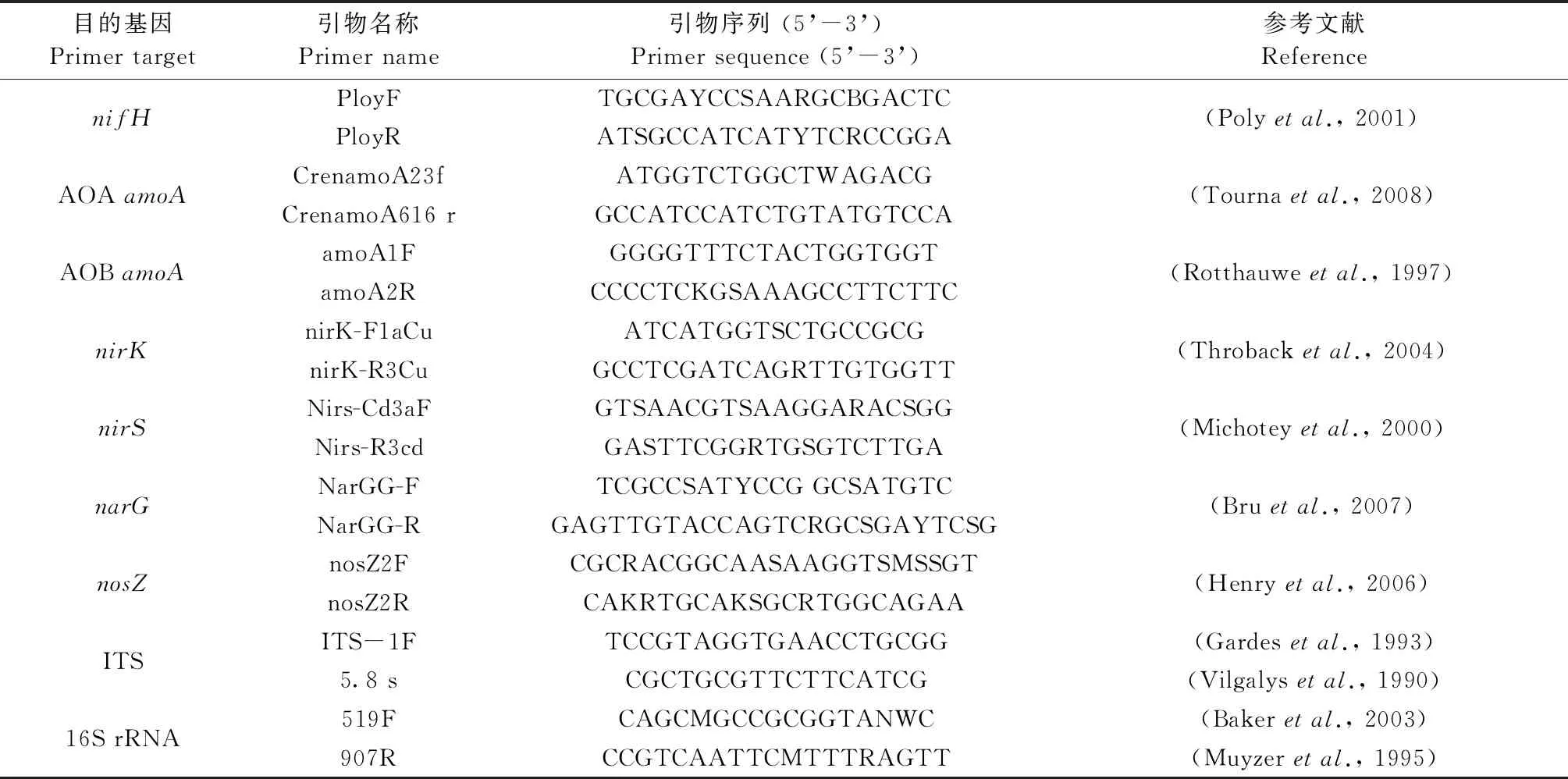
表2 氮循环微生物定量PCR引物
2.5 克隆文库和测序
AOAamoA基因的 PCR 扩增使用和T-RFLP分析相同的引物,引物不添加荧光标记。扩增产物回收后,连接到pGEM-T EasyVector 上,然后将载体转化入大肠杆菌TOP10感受态细胞。从每个克隆文库中随机选择50个克隆子进行测序,具体的克隆子选择方法参考(Singhetal., 2011)。所获得的目的基因序列通过MEGA 6.0 建立 neighbor-joining tree 进行系统发育分析(Tamuraetal., 2013)。本研究所获得的序列信息全部上传到GenBank数据库,其中AOAamoA基因序列的编号从MH141588到MH141591。
2.6 数据处理
利用Excel 2016和SPSS19.0软件进行统计分析。利用单因素方差分析(ANOVA)和多重比较法(LSD)计算不同树种植物特性、土壤基本理化性质、微生物生物量和氮循环功能微生物基因拷贝数的差异显著性。通过Pearson相关系数检验不同树种植物特性与土壤理化性质之间的关系。对各树种森林土壤氮循环功能微生物基因拷贝数与土壤理化性质进行Pearson相关分析。计算各树种森林土壤之间AOA TRFs相对丰度的Bray-Curtis指数,然后利用非度量多维测度法(NMDS)观察样品之间AOA 群落结构的差异,通过Per MANOVA来检测不同树种森林土壤之间AOA群落结构差异的显著性。
3 结果与分析
3.1 土壤理化性质
5种人工林土壤基本理化性质(黏粒和粉粒含量、密度、pH值及全氮、碳氮比、铵态氮含量)均无显著差异(表3)。木荷林土壤砂粒含量显著高于杉木和闽楠林(P<0.05); 杉木林、闽楠和木荷林土壤含水量显著高于福建柏林; 福建柏林土壤全碳含量显著高于杉木林; 木荷林和火力楠林的土壤硝态氮含量显著高于闽楠林和福建柏林; 火力楠林土壤矿质氮含量显著高于闽楠林。
3.2 土壤微生物生物量
火力楠和木荷林的土壤微生物生物量碳(MBC)显著高于闽楠和杉木林(P<0.05)(图1)。火力楠、木荷和福建柏林的土壤微生物生物量氮(MBN)显著高于闽楠和杉木林(P<0.05)。火力楠和木荷林的微生物生物量碳/氮(MBC/MBN)显著高于杉木林(P<0.05)。在5个人工林土壤微生物生物量(MBC、MBN)中,火力楠和木荷林的均最高,而闽楠和杉木林的均最低。

表3 各树种人工林土壤基本理化性质(均值±标准误,n=3)
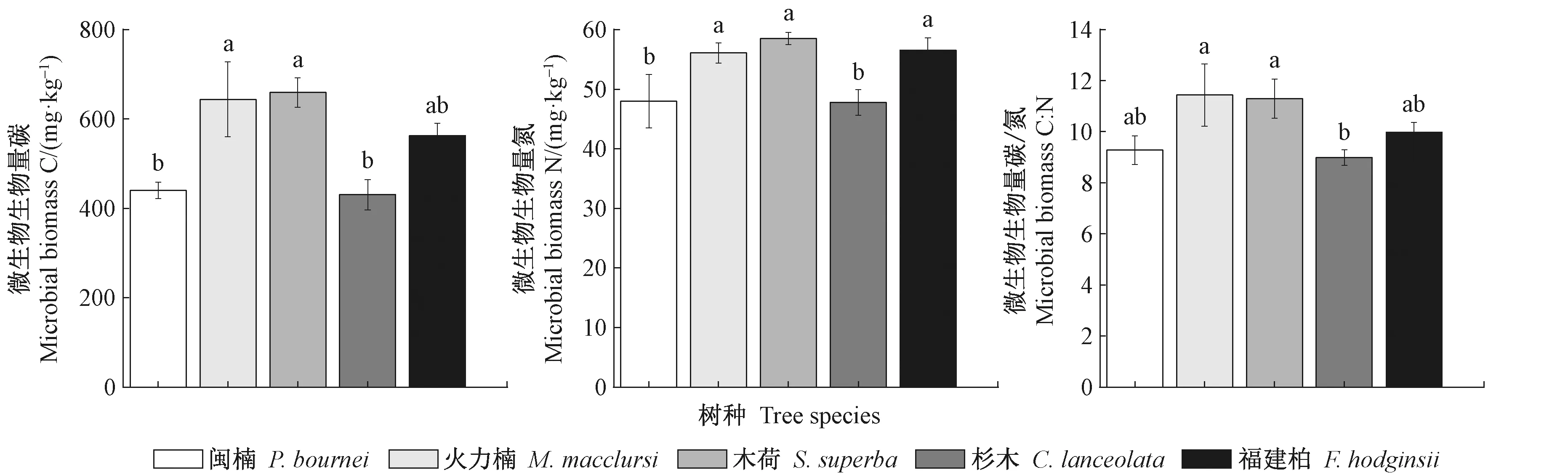
下同。 The same below.
3.3 土壤理化性质与微生物生物量的相关性
对5种人工林的Pearson相关分析表明(表3),土壤全碳含量与植物细根碳含量极显著正相关(P<0.01); 土壤全氮含量与细根碳含量显著正相关(P<0.05); 土壤碳氮比与凋落物碳、细根碳含量显著正相关(P<0.01); 土壤NO3--N含量与凋落物木质素、纤维素含量显著负相关(P<0.05); MBC与凋落物木质素、纤维素含量极显著负相关(P<0.01); MBN与凋落物木质素、纤维素含量显著负相关,而与细根碳含量显著正相关(P<0.05)。
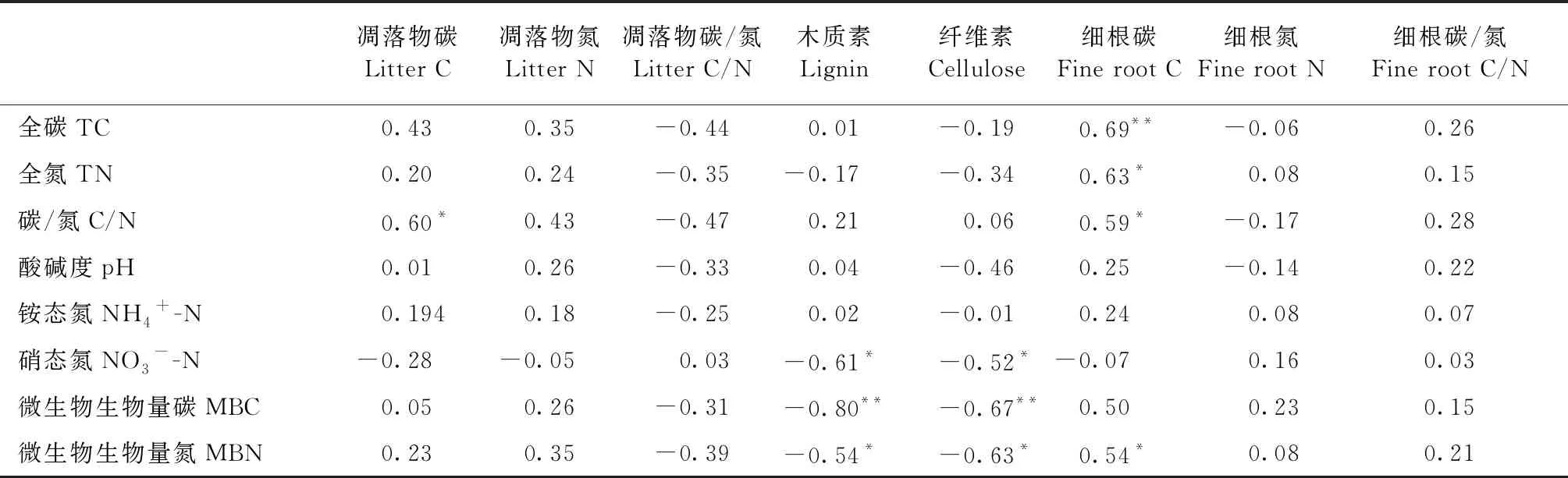
表3 各树种森林土壤理化性质及微生物生物量的Pearson相关分析①
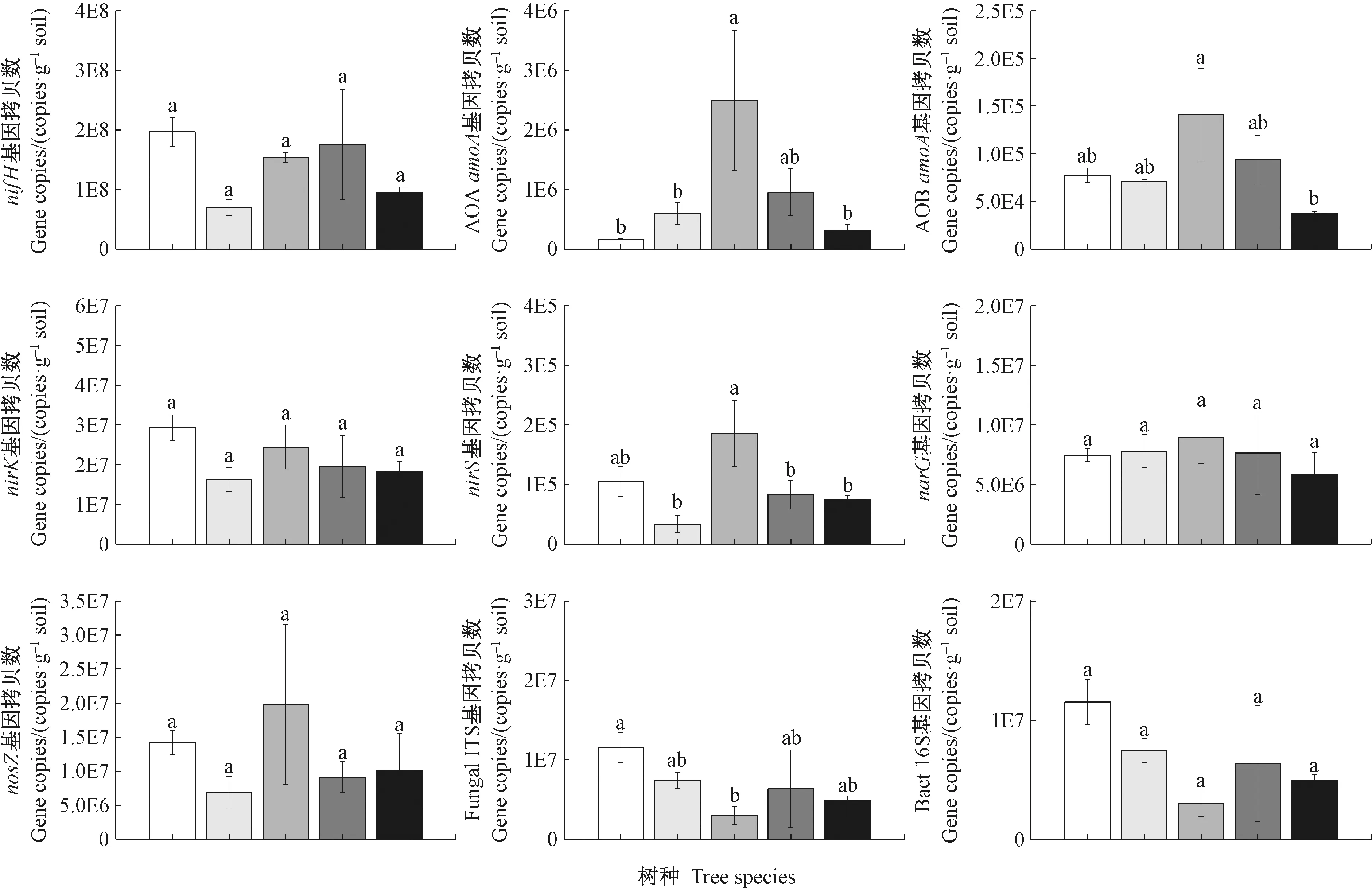
图2 各树种土壤氮循环相关功能微生物基因丰度(均值±标准误,n=3)
3.4 氮循环功能微生物基因丰度
采用定量Q-PCR 检测5种人工林土壤固氮、硝化和反硝化微生物基因丰度,根据标准曲线计算出土壤氮循环功能微生物基因拷贝数量(图2)。5种人工林土壤固氮微生物nifH基因拷贝数无显著差异(P>0.05)。5种人工林土壤AOAamoA基因拷贝数变化范围为1.56×105~2.5×106copies·g-1soil,AOBamoA基因拷贝数变化范围为3.70×104~1.4×105copies·g-1soil。5种人工林土壤AOAamoA基因拷贝数差异显著,其中木荷林的显著高于闽楠、火力楠和福建柏林(P<0.05)。5种人工林土壤AOBamoA基因拷贝数仅木荷显著高于福建柏(P<0.05)。5个树种之间,木荷的AOA和AOBamoA基因拷贝数均最高。5个树种土壤AOA/AOB由高到低排序依次为木荷(17.75)、杉木(10.13)、火力楠(8.52)、福建柏(8.38)和闽楠(2.01)。nirS基因拷贝数变化范围为3.39×104~1.86×105copies·g-1soil,木荷林显著高于火力楠、杉木和福建柏林(P<0.05)。5种人工林土壤narG、nirK和nosZ基因的拷贝数均无显著差异。闽楠林真菌ITS基因拷贝数显著高于木荷(P<0.05)。5种人工林土壤细菌16S rRNA基因拷贝数均无显著差异(P<0.05)。
3.5 土壤氮循环功能微生物基因丰度与土壤理化性质的关系
将5个树种森林土壤氮循环微生物基因拷贝数和土壤理化性质进行Pearson相关分析(表4),表明土壤全碳和土壤AOAamoA显著负相关(P<0.05); 土壤碳氮比与土壤AOAamoA极显著负相关(P<0.01); 土壤pH值与土壤AOBamoA、narG、nosZ显著正相关(P<0.05); 土壤NH4+-N含量与土壤AOAamoA显著负相关(P<0.05); 土壤NO3--N含量与土壤AOAamoA极显著正相关(P<0.01)。
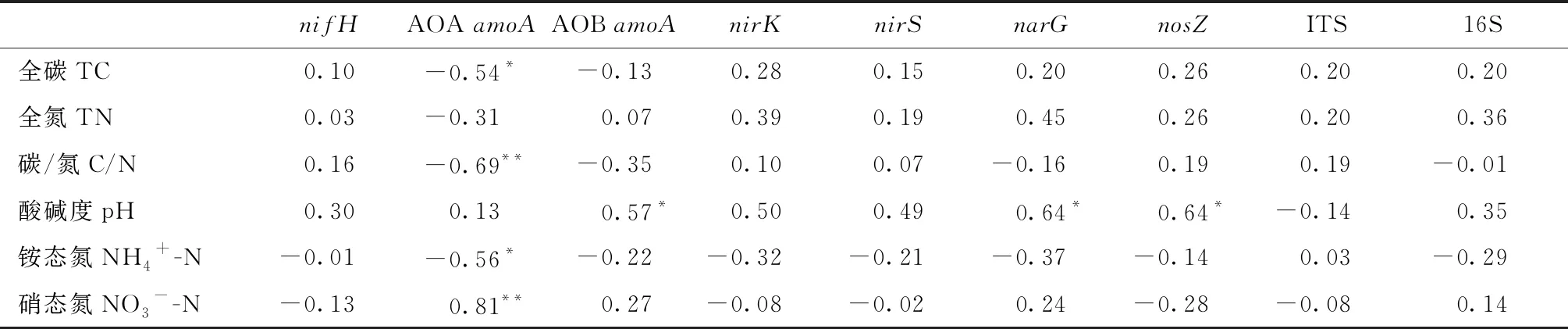
表4 各树种土壤氮循环功能微生物基因拷贝数与土壤理化性质的Pearson相关分析
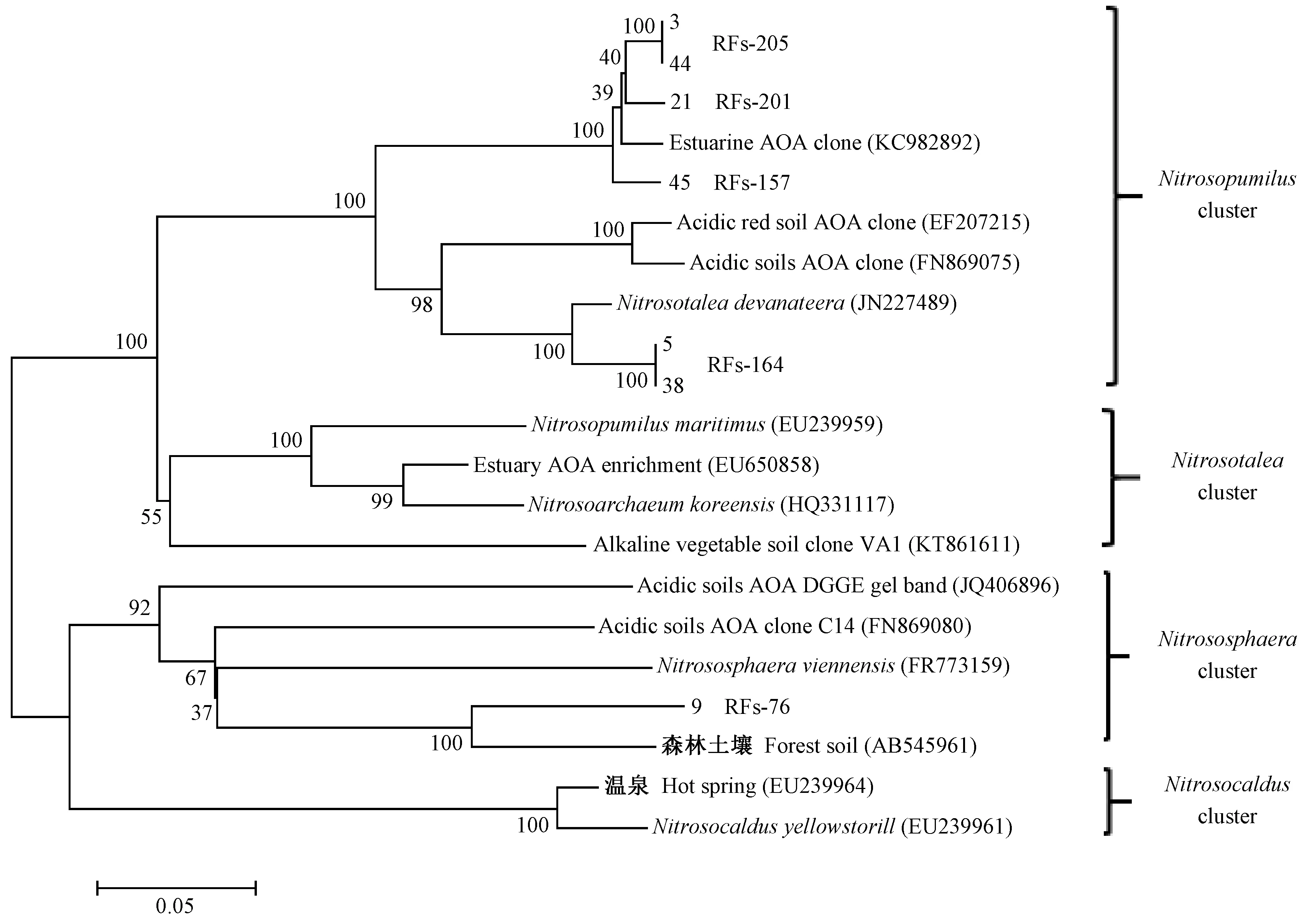
图3 各树种土壤氨氧化古菌AOA amoA群落结构的系统发育分析
3.6 氨氧化古菌群落结构的系统发育和T-RFLP分析
通过对5种森林土壤AOAamoA基因PCR产物进行酶切,分析其群落结构(图3)。限制性酶切片段主要由RFs-205、RFs-202、RFs-157、RFs-164和RFs-76所组成。各树种森林土壤中的AOAamoA属于Nitrosopumilus和Nitrososphaera类群。使用HpyCH4V限制性内切酶对AOAamoA基因PCR产物进行酶切,能够产生4个TRFs,其中TRF-76和TRF-165 是最主要的2个片段类型,分别占总片段的54.88%~100%、0~45.12%(图4),其中TRF-76属于Nitrososphaera类群,TRF-165属于Nitrosopumilus类群。TRF-76和TRF-165的相对丰度对不同树种的响应非常明显,闽楠的TRF-76相对丰度为100%,木荷、杉木、福建柏的TRF-76显著降低; TRF-165的相对丰度在杉木土壤中最高,在闽楠中未检出TRF-165。TRF-86只在福建柏人工林中检出。TRF-156只在木荷人工林中检出。
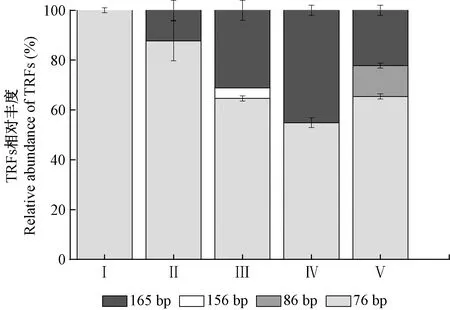
图4 各树种土壤氨氧化古菌AOA amoA基因T-RFs的相对丰度
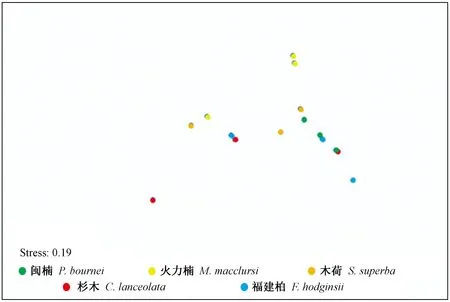
图5 各树种土壤AOA群落结构的非度量多维测度(NMDS)分析
通过非度量多维测度法(NMDS)分析两两样品之间的Bray-Curtis相似性指数(图5)。AOAamoA的群落结构在树种水平上未出现明显分离。进一步PerMANOVA分析发现,不同树种土壤AOAamoA的群落结构差异未达到显著性水平(P>0.05)。
4 讨论
4.1 各树种森林土壤理化性质及微生物生物量的特征
不同树种凋落物质量不同,决定了凋落物分解速率的差异(Zhangetal., 2015),从而影响土壤碳、氮库(万晓华等, 2014)。本研究发现福建柏林的土壤全碳含量显著高于杉木林,这可能由于福建柏凋落物碳氮比、木质素含量显著低于杉木,细根碳含量显著高于杉木(表1),其凋落物及细根分解速率较快。木荷和火力楠林的土壤硝态氮含量显著高于闽楠和福建柏林,土壤硝态氮含量与凋落物木质素、纤维素含量显著负相关(表3)。这可能由于闽楠和福建柏林凋落物的难分解物质(木质素、纤维素)含量较高,微生物难以分解利用进而将氮素释放出来。在森林生态系统中,硝态氮的淋失和反硝化是土壤氮素损失的重要途径(Huygensetal., 2008)。木荷和火力楠林的土壤硝态氮库较大,这表明该树种可能不利于土壤氮保留。
不同森林类型土壤微生物生物量存在显著差异。杉木连栽导致土壤中酚类物质累积,从而抑制土壤微生物生物量和活性(李秋玲等, 2012)。本研究发现,火力楠和木荷林的MBC显著高于闽楠和杉木林,火力楠、木荷和福建柏林的MBN显著高于闽楠和杉木林。Pearson相关分析可知,MBC与木质素、纤维素含量极显著负相关; MBN与木质素、纤维素含量显著负相关,而与细根碳含量显著正相关。凋落物的木质素和纤维素含量显著影响土壤微生物生物量。
4.2 不同树种对氮循环相关功能微生物基因丰度的影响
沈秋兰等(2016)研究发现,毛竹(Phyllostachyedulis)林土壤固氮微生物nifH基因拷贝数显著高于阔叶林,土壤氮素水平明显提高。但本研究5种森林土壤的固氮微生物nifH基因拷贝数并无显著差异,这与Reverchon等(2015)的研究结果一致。这可能和各树种森林土壤pH值无显著差异有关,大部分土壤固氮细菌数量受土壤pH值影响较大,有其最适合的土壤pH值范围(周移国等, 2013)。
He等(2007)对中国南方不同施肥处理下酸性红壤的调查发现,AOA 在数量上占绝对优势,AOA与 AOB比值的范围从1~12。本研究中5种森林土壤中AOA基因丰度显著高于AOB丰度,且AOA/AOB>2,与AOB相比,AOA在酸性森林土壤中更有优势,这与李永春等(2014)研究结论相符合。土壤pH值是影响氨氧化微生物分布的最主要因素,Nicol等(2008)发现在pH值范围4.9~7.5内,随土壤pH值降低,AOA基因拷贝数及表达活性升高。本研究相关分析发现AOB丰度与土壤pH值显著正相关,而AOA与土壤pH值并无显著相关性。这可能与5种人工林土壤pH值无显著差异且样本数量少有关。Zhang等(2012) 通过稳定性同位素探针技术(DNA-SIP)研究发现,AOA而非AOB主导了酸性土壤中的硝化作用。本研究土壤NO3--N含量与AOA极显著负相关,而与AOB无显著相关性,这可能是因主要由AOA驱动了酸性森林土壤氨氧化过程。木荷的AOA 丰度显著高于闽楠、火力楠和福建柏,AOB丰度显著高于福建柏。这可能意味着木荷土壤的硝化作用在5种人工林中表现最强,加剧硝态氮淋溶和经反硝化作用气态氮损失的风险,不利于氮保留。
反硝化功能微生物中narG、nirK和nosZ的基因拷贝数在5种森林土壤间均无显著差异,这可能由于5种森林土壤的基本理化性质差异不显著。木荷林nirS的基因拷贝数显著高于火力楠、杉木和福建柏林,这可能和木荷林土壤的AOA、AOB丰度最高有关。Ribbons等(2016)研究发现,AOA和AOB基因拷贝数的总和与反硝化功能基因nirK与nirS的基因拷贝数的总和之间显著正相关,氨氧化微生物通过硝化作用将铵态氮转化成硝态氮,为反硝化作用提供底物,从而增加反硝化微生物基因丰度。Zhang等(2018)研究发现,潜在的反硝化速率和nirK、nirS基因丰度密切相关。反硝化作用是氮素损失的重要途经,木荷林土壤nirS的基因拷贝数显著高于火力楠、杉木和福建柏林,这可能使木荷林土壤的温室气体(N2O、NO)排放量较高,不利于森林生态系统的氮保留。
5种森林土壤真菌ITS基因丰度只有闽楠林显著高于木荷林,这可能因为闽楠林的凋落物,难以被微生物分解利用。与之相反,Ribbons等(2016) 研究发现,ITS基因丰度在不同树种和样地间均无显著差异。Petersen等(2012)发现细菌16S rRNA基因丰度和总氮矿化速率显著相关,但本研究中5种森林土壤的16S rRNA基因丰度没有显著差异。
4.3 不同树种对土壤氨氧化古菌群落结构的影响
通过克隆和测序发现AOA类群属于Nitrosopumilus和Nitrososphaera,其中TRF-76占总片段的54.88%~100%,属于Nitrososphaera,占绝对优势。本研究结果与前人结果相似,Zhang等(2012)发现,Nitrososphaera在强酸性土壤(pH<4.5)的硝化作用中发挥着重要作用。早有研究报道酸性土壤中存在属于Nitrosopumilus的AOA类群(Heetal., 2007)。
本研究用HpyCH4V限制性内切酶对AOAamoA基因PCR产物进行酶切,仅得到4个TRFs。胡行伟(2013)对澳大利亚桉树(Eucalypusspp.)林土壤中AOA使用RsaI限制性内切酶得到12个TRFs。TRFs数量不同可能是由于生态系统类型不同所致,也可能是内切酶的不同。李永春等(2014)发现不同林分通过土壤速效钾、pH值和有机质含量引起土壤AOA群落结构显著差异。但本研究发现不同树种的AOA群落结构无显著差异,可能是由于不同树种的土壤pH值、TN、C/N、 NH4+-N含量没有显著差异,且微生物群落结构对环境因子改变的响应滞后造成的。Prescott等(2013)认为,树种对土壤的影响具有较强环境依赖性,树种效应可能在肥沃或中等肥力水平的土壤中更易检测到。Ribbons等(2016)认为,树种的影响小于立地,因此需要未来在同质园或更严格试验条件下研究树种对土壤氮保留的影响。
5 结论
在5种亚热带森林的酸性土壤中,氨氧化古菌AOA在数量上占据优势地位,在土壤硝化作用中可能发挥着主导作用,木荷林的AOA和AOB丰度最高,nirS的基因丰度显著高于火力楠、杉木和福建柏林,木荷林的AOA丰度显著高于闽楠林,可能加剧氮素从森林生态系统损失的风险。5种森林土壤的AOA类群属于Nitrosopumilus和Nitrososphaera,AOA群落结构无显著差异。
猜你喜欢
北京大学学报(自然科学版)(2022年4期)2022-08-18
中国林副特产(2022年4期)2022-08-08
散文诗世界(2022年5期)2022-05-23
右江民族医学院学报(2022年2期)2022-05-19
河北医学(2021年10期)2021-10-27
花卉(2021年2期)2021-01-21
林业与环境科学(2020年3期)2020-07-21
农家科技下旬刊(2020年2期)2020-04-07
中国临床医学影像杂志(2019年6期)2019-08-27
中华实验和临床病毒学杂志(2016年3期)2016-08-09
