
防城港湾海域甲壳类群落结构及其与环境因子的关系
2020-09-16 07:21何思璇赖廷和
应用海洋学学报 2020年3期
何思璇,赖廷和
(1.中央民族大学生命与环境科学学院,北京100081; 2.广西壮族自治区海洋研究院,广西 南宁 530022)
海洋甲壳类是海洋食物网能量流动和物质循环中的重要环节,同时是生命周期短、经济价值较高的优质水产资源[1]。我国学者对海洋甲壳类的研究涉及区系、多样性、群落生态和资源经济利用等多个方面[1-10]。随着开发强度不断加大,近年来我国沿海河口海湾面临着日趋严重的资源过度利用和环境污染压力,海洋生物资源呈现低值化和小型化趋势[6-7],因此开展更多的海洋甲壳类群落结构动态监测,了解影响海洋渔业资源变化与环境因子的关系,为海洋资源可持续利用提供科学依据,具有重要意义。
防城港湾(21°32′30″ ~21°43′00″N,108°17′00″~108°29′00″E)位于广西防城港市江山半岛与企沙半岛之间,海域面积为126.95 km2,具有优良的区位优势和资源条件,是建设北部湾区域性国际航运中心的重要港区。遥感影像解译的结果表明:与1990年相比,大规模围填海使得该湾的湾口宽度变窄了约20%,湾内滨海湿地面积丧失了29.69%(3 769.40 hm2),原生红树林被毁灭了24.37%(203.84 hm2)。防城港湾是一个河口湾,防城河自港湾北部注入,多年平均径流量为1.786×109m3,流域面积为894.6 km2,2017年经防城河入海的污染物总量达15 030 t,防城港湾成为全市污染最严重的海域[11]。因此,防城港湾的海域开发利用与环境污染的影响应引起高度重视。游泳动物的群落结构与功能直接反映海洋生态系统状态,被视为表征海洋生态健康的关键指标[12]。但以往范围广、较系统的广西海域游泳甲壳类生物生态调查多集中在湾外较深海域,而对沿岸河口、海湾的调查则存在频次低、范围窄、站位少、重复性差等问题[13-15],难以查清海洋甲壳类渔业资源的趋势性变化及其受胁机制。因此本研究在防城港湾开展了甲壳类资源数量和结构及相关环境因子的季节动态调查,分析探讨该湾甲壳类种类组成、群落结构特征及环境因子的影响,以期为防城港湾海洋资源与环境的管理保护提供科学依据。
1 材料与方法
1.1 调查站位分布与采样方法
本研究在防城港湾设置了21个游泳甲壳类生物生态调查站位(图1),于2016年5、8、11月和2017年2月的大潮期,租用渔船开展单船型底拖网采样作业。所用底拖网的曳纲长度为30 m,网口宽度为3 m,囊网网目为1.0 cm。每站实施拖网15 min,下网期间将船速固定在3 kn。将所有渔获现场冷藏,带回实验室后按种计数和称量。可捕系数取0.8,综合可捕系数、网口宽度、拖网时间和船速计算渔获密度(尾/km2)和生物量(kg/km2)。同时,现场测定水温、水色、水深、透明度、溶解氧(DO)含量、pH、盐度等因子,并采集水样带回实验室,分析叶绿素a、悬浮物、生化需氧量(BOD)、硅酸盐、化学需氧量(COD)、总氮、氨氮、硝酸盐、亚硝酸盐、无机氮、总磷、无机磷等的含量,总共分析测定了19项环境因子。严格按照《海洋监测规范》[16]和《海洋渔业资源调查规范》[17]的要求进行环境因子现场测定、水质和动物样品采集保存、实验室分析测定及资料处理。
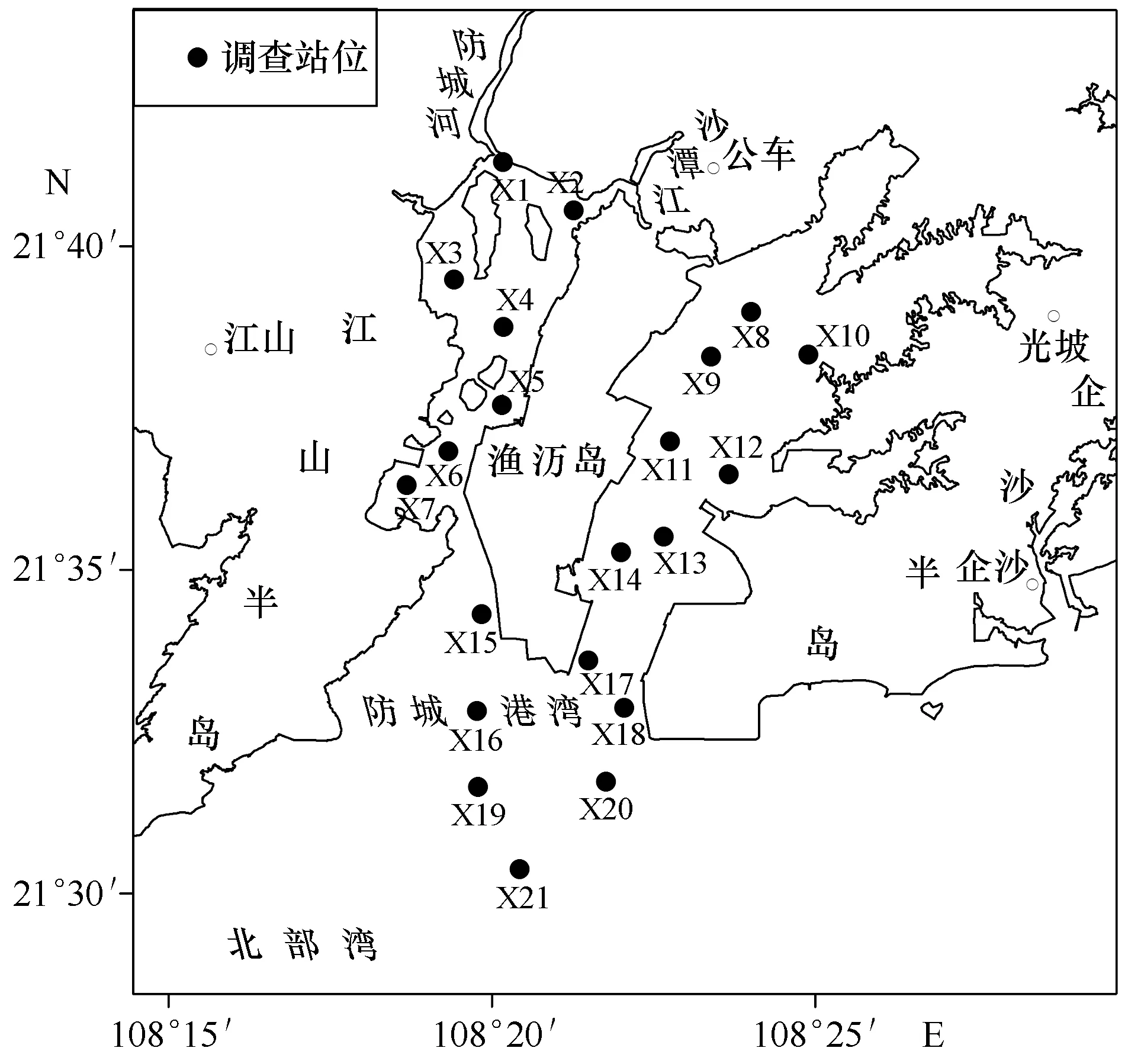
图1 采样站位分布Fig.1 Distribution of sampling stations
1.2 相关指数计算及数据分析
1.2.1 相关指数计算 采用如下公式计算甲壳类群落的相关指数:
(1)
d=(S-1)/log2N
(2)
J=H′/Hmax
(3)
Jc=c/(a+b-c)×100
(4)
A=C/(C+B)×100
(5)
IRI=(ni/N+wi/W)×F×10 000
(6)
式(1~3)中:H′为Shannon-Wiener种类多样性指数,Pi为某样品中第i种的个体数(ni)占该样品总个数(N)的比值(ni/N),S为该样品鉴定出的物种总数,Hmax为log2S,表示多样性指数的最大值;d为Margalef丰富度指数,J为Pielou均匀度指数[18]。
式(4)中:Jc为物种相似性指数(%),a、b为相比较的两个海区的动物种数(种),c为两个海区的共有种数(种)[18]。
式(5)中:A为某季节与其前一季节比较的种类更替率(%),C为相邻两个季节间减少及增加的物种数(种),B为相邻两个季节间相同的物种数(种)[19]。
式(6)中:IRI为相对重要值,ni为某季或全年第i个种群的个体数,N为某季或全年的个体总数,wi为某季或全年第i个种群的生物量,W为某季或全年所有种群的生物量,F为某季或全年第i个种群的捕获频率[20]。根据陈国宝等(2007)的分级标准,IRI≥500为优势种,100≤IRI<500为主要种,10≤IRI<100为一般种,IRI<10为少见种[20]。
1.2.2 数据分析 在SPSS Statistics 23.0软件中,采用单因素方差分析检验甲壳类群落的密度、生物量和多样性指数的差异显著性。采用PRIMER v6软件开展群落相似性分析,先将不同季度、站位的甲壳类种群密度进行4次方根转换,筛选贡献率大于5%种群的密度数据计算Bray-Curtis相似性矩阵,采用等级聚类分析进行群落分组,采用相似性分析检验不同分组之间的差异显著性,采用相似性百分比分析找出导致各分组内部群落结构相似的典型种,采用生物-环境分析找出与甲壳类群落结构相关性最大的环境因子[21-23]。
2 结果与讨论
2.1 群落种类组成
4个季度底拖网调查共采集到甲壳类60种,隶属于2目19科35属,包括口足目虾蛄类3种,十足目57种(含虾类25种,蟹类32种)。以对虾科种数最多,共17种,其次为梭子蟹科,有11种。种类组成以暖水性种为主,有36种,占60.0%;暖温性种24种,占40.0%,具有明显的亚热带性质。经济型种类较多,对虾科的日本囊对虾(Marsupenaeusjaponicus)、墨吉明对虾(Fenneropenaeusmerguiensis)、长毛明对虾(Fenneropenaeuspenicillatus)、宽沟对虾(Melicertuslatisulcatus)、刀额新对虾(Metapenaeusensis)和近缘新对虾(Metapenaeusaffinis)等,梭子蟹科的梭子蟹属(Portunus)和蟳属(Charybdis)种类,及虾蛄科的口虾蛄(Oratosquillaoratoria),是北部湾海域中价值较高的甲壳类海产品。
在春季共捕获51种,夏季为34种,秋季为35种,冬季为26种。季节出现频次为1、2、3、4季的种数分别有21、12、8、19种,分别占总种数的35.0%、20.0%、13.3%、31.7%。季节更替率依次为38.6%(春—夏)、3.8%(夏—秋)、28.1%(秋—冬)、51.0%(冬—春)。夏季与秋季之间更替率最低,可能与这两季水温差距小有关。
全年在各站位采集到的甲壳类种数差异较大,X1站种数最少,仅3种;X17站种数最多,有27种,X20站次之,有26种(图2)。从图2可以看出,在湾口的东部海域存在1个种数高值区域,包括了X17、X18和X20等3个站位。
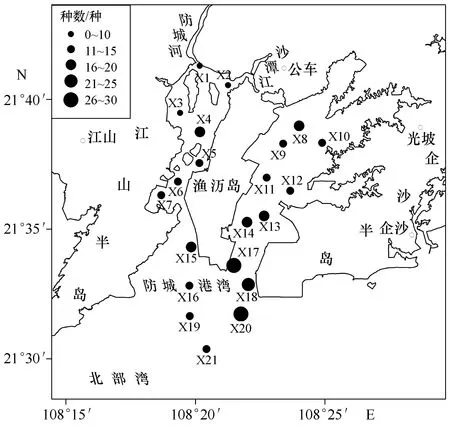
图2 甲壳类种数空间分布Fig.2 Distribution of crustacean species numbers
将所有21个站分为西湾(X1~X7)、东湾(X8~X14)和湾口(X15~X21)3个海区,统计表明湾口有46种,东湾有41种,西湾有36种。东湾与湾口共有种为32种,相似性指数为58.2%;东湾与西湾共有种为27种,相似性指数为54.0%;西湾与湾口共有种为27种,相似性指数为49.1%。3个海区的共有种为24种,包括刀额新对虾、亨氏仿对虾(Parapenaeopsishungerfordi)、近缘新对虾、日本囊对虾、钝齿蟳(Charybdishellerii)和银光梭子蟹(Portunusargentatus)这些广布种,占总种数(60种)的40.0%。可见区域之间的种类组成存在一定程度异化。在东湾和西湾盐度较低的站位捕获到细螯沼虾(Macrobrachiumsuperbum)、脊尾白虾(Exopalaemoncarinicauda)等河口种及养殖逃逸种凡纳滨对虾(Litopenaeusvannamei),但这些种类不出现在湾口海域;而哈氏仿对虾(Parapenaeopsisharbwickii)和锈斑蟳(Charybdisferiatus)仅在湾口海域捕获到少量个体。
2.2 群落优势种
由表1可知,优势种和主要种共18种,其中虾类11种,蟹类5种,虾蛄类2种。不同季节中甲壳类群落优势种组成有所差异,春季优势种和主要种均为6种;夏季优势种有5种,主要种有3种;秋季优势种有6种,主要种有7种;冬季优势种有7种,无主要种。周年优势种有亨氏仿对虾、日本囊对虾、钝齿蟳、刀额新对虾和近缘新对虾等5种,主要种有6种:须赤虾(Metapenaeopsisbarbata)、长毛明对虾、中型新对虾(Metapenaeusintermedius)、口虾蛄、周氏新对虾(Metapenaeusjoyneri)和远海梭子蟹(Portunuspelagicus),一般种有12种,少见种有37种。优势度最大的优势种为:亨氏仿对虾(春季)、刀额新对虾(夏季)、近缘新对虾(秋季)、钝齿蟳(冬季)。各个季节所有优势种的相对重要值之和的占比分别为85.8%(春季)、83.6%(夏季)、78.8%(秋季)和97.0%(冬季),冬季优势种集中度最为明显,秋季则相对较弱。
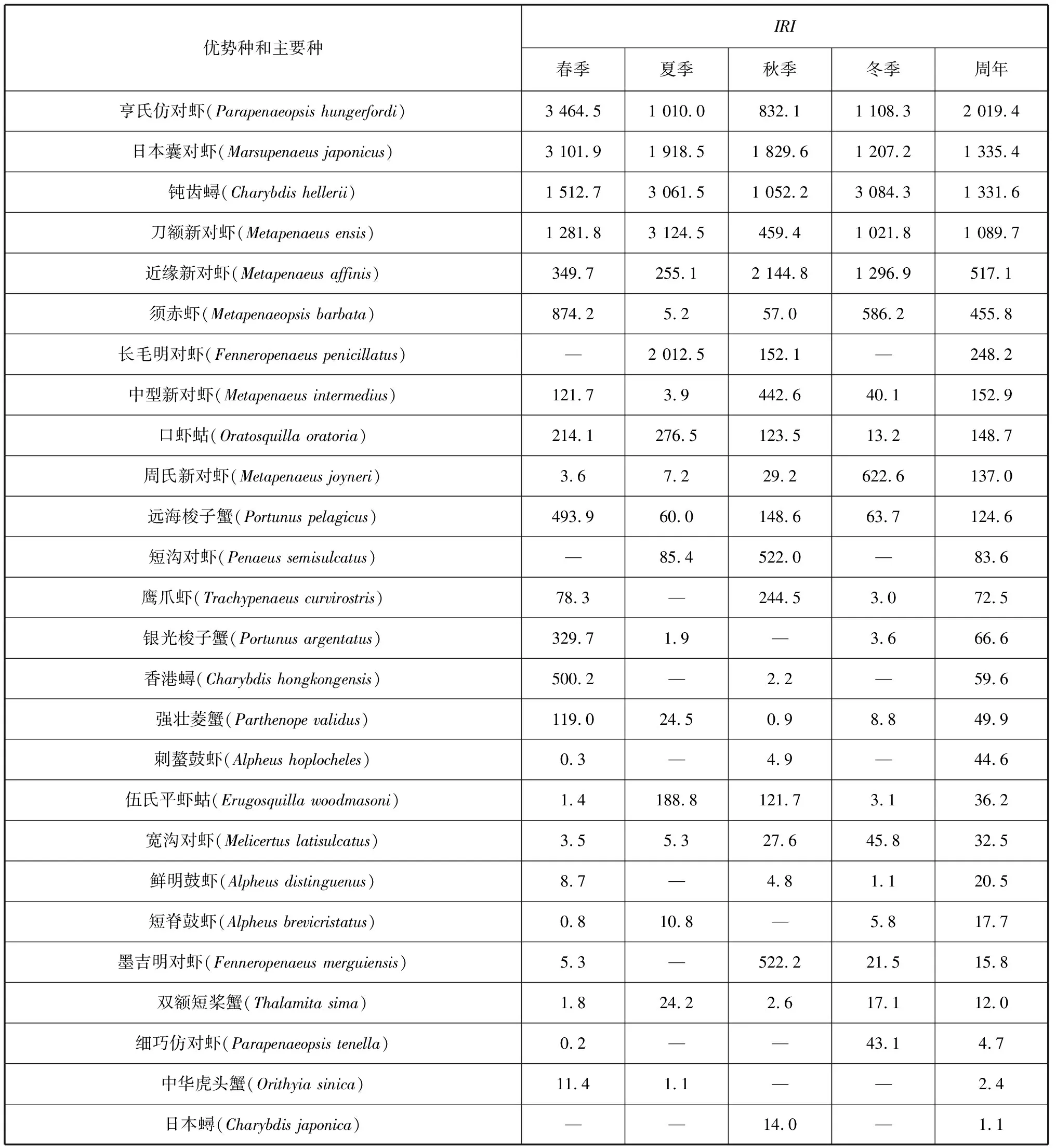
表1 防城港湾甲壳类优势种、主要种及其相对重要值
2.3 密度与生物量
防城港湾甲壳类的年均密度为32 201尾/km2,全湾现存量为4.1×106尾,虾类、蟹类和虾蛄类分别占81.3%、15.6%和3.1%。从图3可以看出,4个季节均出现高密度区,春季高密度区出现在湾口,夏季在湾口东部密度最大,秋季的高密度区位于东湾和湾口相连的海区,冬季高密度区分别位于西湾中部和湾口东北部海域。春季的X17站密度最高,达323 889尾/km2;秋季的X4站密度最低,仅有1 578 尾/km2。甲壳类密度的季节变化非常明显,春季为67 391尾/km2,夏季为25 502尾/km2,秋季为18 589尾/km2,冬季为17 321尾/km2;4个季节之间的密度差异极显著(F3,80=5.430,p<0.01)。不同海区密度大小顺序为:湾口(59 963尾/km2)>西湾(19 245尾/km2)>东湾(17 395尾/km2),海区之间的密度差异极显著(F2,81=7.404,p<0.001)。但站位间的密度差异不显著(F20,63=1.324,p>0.05),主要是由于组内(即季节)密度差异较大,均方数值大,导致F值较低。
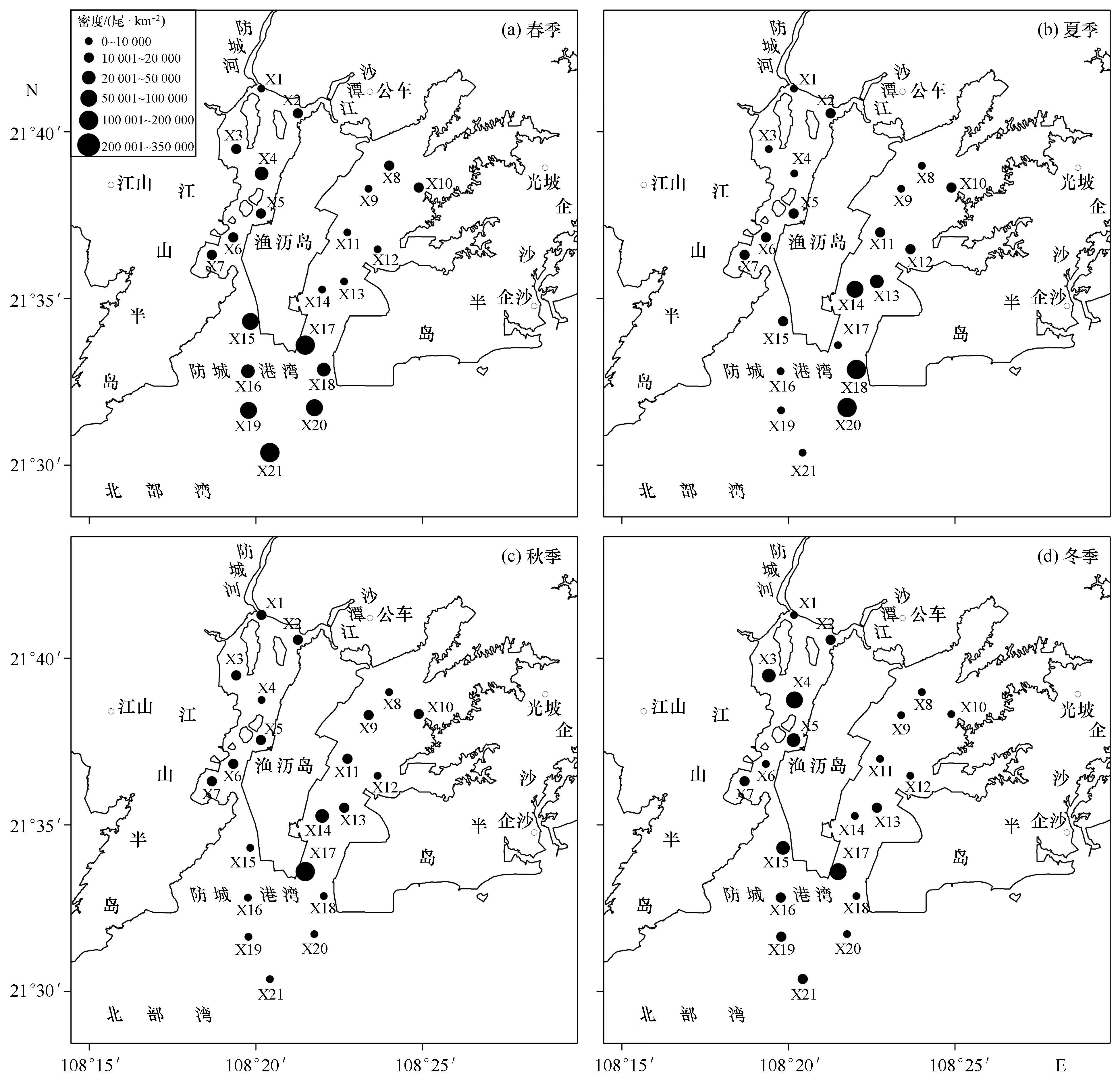
图3 不同季节防城港湾甲壳类密度分布Fig.3 Distribution of crustacean density in four seasons in Fangchenggang Bay
研究海域的甲壳类年均生物量为212.290 kg/km2,全湾现存量是4 088 t,虾类、蟹类和虾蛄类分别占68.0%、27.1%和4.9%。各季节均出现生物量高值区,春季集中在湾口海域,夏、秋季高值区分布在连接东湾和湾口的深水海域,冬季高值区则在湾口的北部区域(图4)。最高生物量出现在夏季的X20站,达1 097.43 kg/km2;最小值出现在秋季的X4站,仅7.65 kg/km2。不同海域分区之间的生物量差异极显著(F2,81=6.292,p<0.01),表现为:湾口(331.22 kg/m2)>东湾(176.36 kg/m2)>西湾(129.28 kg/m2)。各季节的甲壳类生物量大小顺序分别为:春季(346.68 kg/km2)>夏季(199.91 kg/km2)>秋季(168.45 kg/km2)>冬季(134.11 kg/km2),季节之间差异显著(F3,80=3.587,p<0.05)。站位间生物量差异不显著(F20,63=1.389,p>0.05)。
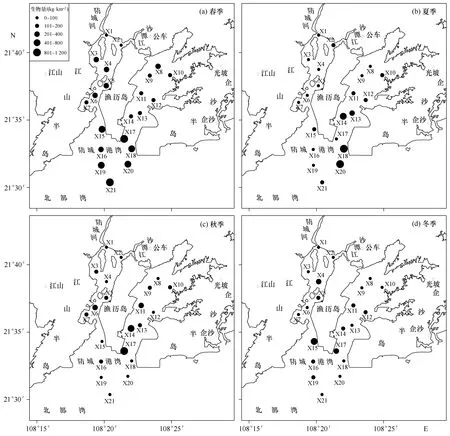
图4 防城港湾4个季节甲壳类生物量分布Fig.4 Distribution of crustacean biomass in four seasons in Fangchenggang Bay
2.4 多样性指数
从图5(a)可以看出,防城港湾甲壳类群落的各站种数最多达17种,最少仅2种。6~7种的站位较多,占29.8%;10种以上的站位占22.6%。4个季节的甲壳类种数多少依次为:春季(9.0)>秋季(7.1)>夏季(6.2)>冬季(5.2),季节之间差异显著(F3,80=3.493,p<0.05)。不同海域分区之间的种数差异极显著(F2,81=5.415,p<0.01),表现为:湾口(8.5种)>东湾(7.1种)>西湾(5.1种)。但站位间的种数差异不显著(F20,63=1.970,p>0.05)。
从图5(b)可以看出,H′范围在0.200~3.160之间。H′的平均值为1.847,小于2.000的站位占54.8%,大于3.000的站位仅占2.4%,较多区域的甲壳类群落受到扰动,大多数站位种数少且优势种很突出。各分区的H′均值分别为:东湾(2.210)>湾口(1.799)>西湾(1.531),差异极显著(F2,81=7.657,p<0.001)。站位间H′值的差异极显著(F20,63=3.087,p<0.001)。秋季H′值高于其他3个季节,但季节之间差异不显著(F3,80=1.550,p>0.05)。
从图5(c)可以看出,d值范围在0.200~2.230。d的均值为1.056,大于1.000的站位占54.8%。各分区的d值分别为:湾口(1.396)>东湾(1.356)>西湾(0.975),差异极显著(F2,81=5.776,p<0.01)。站位间差异极显著(F20,63=2.576,p<0.01)。各季节的d值高低依次为:秋季(1.210)>春季(1.189)>夏季(0.928)>冬季(0.897),季节间差异不显著(F3,80=2.276,p>0.05)。
从图5(d)可以看出,J值范围在0.160~0.988之间。J的均值为0.734,高于0.800的站位占41.7%。季节间J值差异极显著(F3,80=4.067,p<0.01)。3个分区的J均值分别为:东湾(0.808)>西湾(0.720)>湾口(0.674),差异显著(F2,81=4.265,p<0.05),湾口群落的优势种更为突出。但站位间差异不显著(F20,63=0.912,p>0.05)。
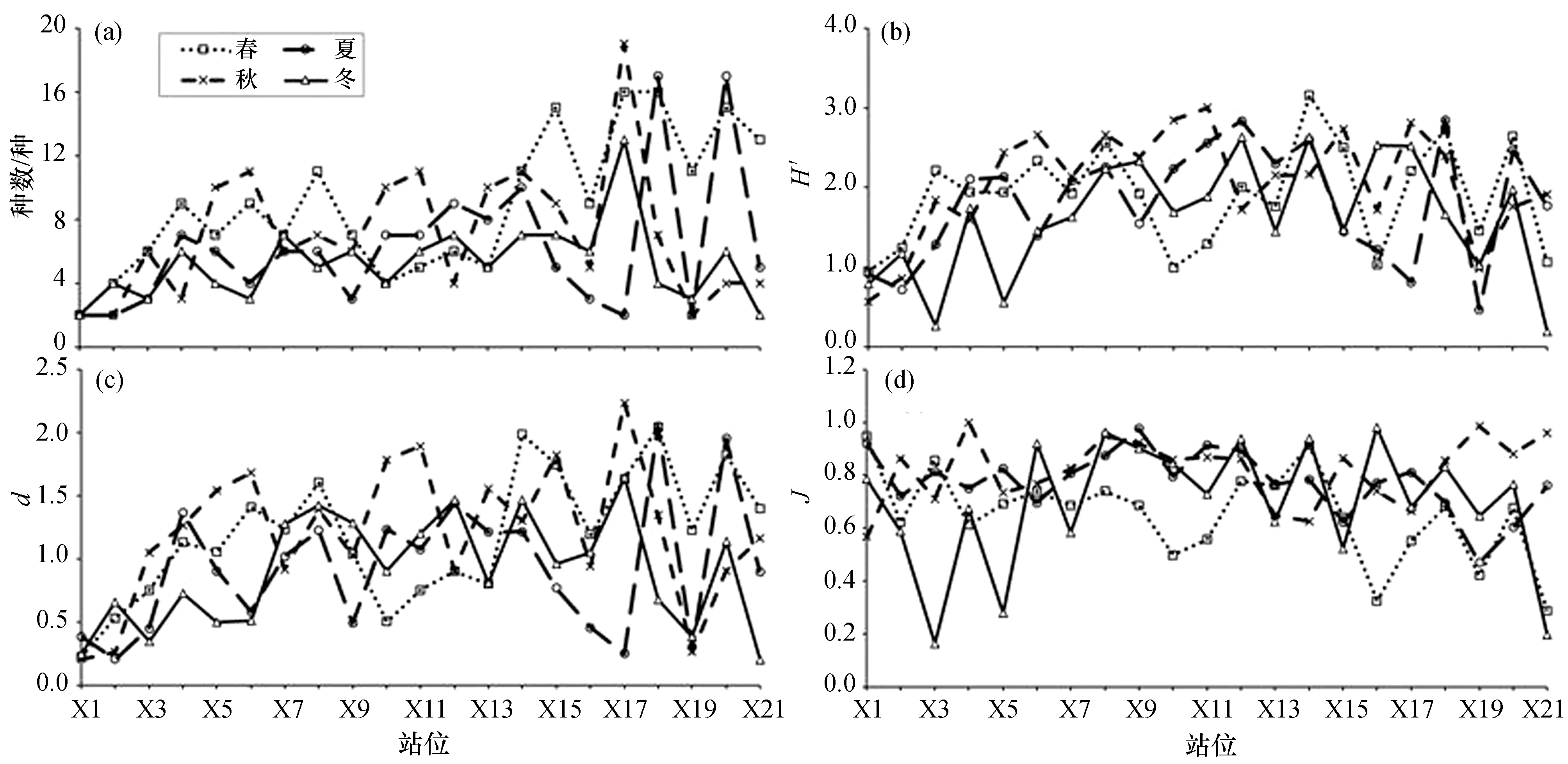
图5 防城港湾甲壳类多样性指数季节变化Fig.5 Seasonal change of diversity indexes of crustacean in Fangchenggang Bay
2.5 群落结构相似性分析
不同季节群落分组结果明显不同(图6),等级聚类分析显示春季群落主要分为两个站位组(R=0.785,p<0.001),a1组由西湾和东湾的所有14个站位组成,a2组由湾口站位组成,站位X17独立于上述两组,如图6(a)所示。从图6(b)可以看出,夏季群落可分为两个组(R= 0.509,p<0.01),第一组包括X18和X 20这两个深水站,第二组包括其余19个站。秋季群落分为4个组(R=0.901,p<0.001),c1组由西湾顶部站位组成,c2组由湾口站位组成,c3组由两个相邻的深水站位X14和X17组成,c4组由西湾中部与东湾顶部的站位组成,如图6(c)所示。冬季群落可分为两个组(R= 0.541,p<0.001),d1组主要由湾顶水浅的站位组成,d2组主要由位于航道及湾口的站位组成,如图6(d)所示。总体上秋季甲壳类群落结构异质性高于春、夏、冬季。
相似性百分比分析表明,春季两组的组内平均相似性分别为56.1%和61.3%。a1组的典型种包括日本囊对虾、刀额新对虾和钝齿蟳,对组内相似性的累积贡献率达81.8%。a2组的主要典型种为亨氏仿对虾、银光梭子蟹、日本囊对虾和刀额新对虾等,贡献率达83.7%。
夏季两组的组内平均相似性分别为43.8%和71.9%。b1组的典型种为日本囊对虾、刀额新对虾、亨氏仿对虾和长毛明对虾,贡献率达81.5%;b2组的主要典型有刀额新对虾、钝齿蟳、亨氏仿对虾和伍氏平虾蛄等,累积贡献率达80.4%。
秋季c1~c4组的组内平均相似性分别为48.4%、53.0%、62.9%和53.2%。c1组典型种是刀额新对虾,贡献率达100%;c2组主要典型种包括日本囊对虾和鹰爪虾,贡献率分为42.3%和40.7%;c3组主要典型种包括近缘新对虾、亨氏仿对虾、伍氏平虾蛄、墨吉明对虾、钝齿蟳等,累积贡献率为80.2%;c4组的主要典型种有短沟对虾(Penaeussemisulcatus)、亨氏仿对虾、日本囊对虾、中型新对虾、长毛明对虾等,累积贡献率为80.2%。
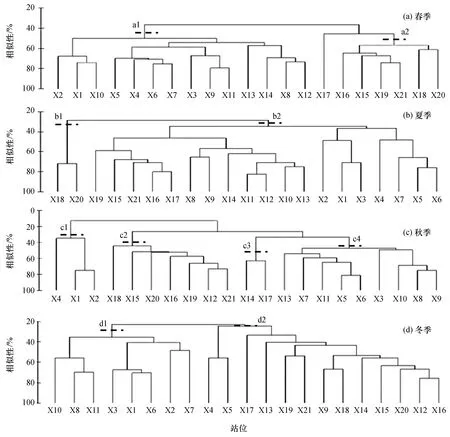
图6 防城港湾甲壳类群落等级聚类图Fig.6 Hierarchical cluster dendrogram of crustacean communities in Fangchenggang Bay
冬季两组的组内平均相似性分别为43.1%和40.6%。d1组的典型种有刀额新对虾、日本囊对虾和钝齿蟳,累积贡献率达80.5%;d2组主要典型种有亨氏仿对虾、钝齿蟳、周氏新对虾、须赤虾和近缘新对虾等,贡献率为85.4%。
2.6 群落结构与环境因子的相关性
考虑到季节间群落结构差异较大而环境因子较相近,本研究将数据分为4个季节单独进行群落结构与环境因子的相关性分析。生物-环境分析表明群落结构与环境因子的关系具有复杂性和综合性(表2)。在春季,甲壳类群落结构与水深、pH、盐度、氨氮和总氮等5个指标组合的相关系数值最高(R=0.467,p=0.003)。夏季相关性最高的是水深、水色、水温、COD、亚硝酸盐、总磷、悬浮物和叶绿素a等8个指标的组合(R=0.493,p=0.005)。在秋季,相关性最高的环境因子是盐度(R=0.505,p=0.005)。冬季相关性最高的是DO、pH、透明度、水色、BOD、氨氮、硅酸盐和叶绿素a等8个指标的组合(R=0.619,p=0.005)。
2.7 讨论
2.7.1 群落种类组成 北部湾海域潮下带甲壳类动物种类非常丰富。根据相关资料记载,1984年连续每月在北部湾涠洲岛以北海域开展的底拖网和阿氏拖网调查渔获中鉴定出甲壳类动物196种,包括虾类31种,蟹类153种,虾蛄类12种[13]。贾晓平等(2003)于2001年秋、冬季在北部湾涠洲岛以南海域开展了48个站位的底拖网调查,共获甲壳类64种,包括虾类24种,蟹类31种,虾蛄类9种[24]。谢旭等(2017)于2001年11月至2002年1月在琼州海峡海域调查发现虾类19种和蟹类31种[9,25]。
本研究全年在防城港湾海域采用底拖网采集到甲壳类共计60种,包括虾类25种,蟹类32种,虾蛄类3种。暖水性种和暖温性种分别占60.0%和40.0%,该湾甲壳类群落具有明显的亚热带性质,这是由它位于北回归线的南部边缘这一地理区位决定的。防城港湾中4个季度均出现的甲壳类有19种,仅占总种数的31.7%,可见很多种类只在某些季节出现,季节性洄游种类较多,地方性种类相对较少。从图2可看出水深大的湾口站位种数比东湾、西湾站位更多,表明了河口湾有不少甲壳类动物来自湾口以外海域,同时还有某些种类如脊尾白虾、中华绒螯蟹(Eriocheirsinensis)等来自河口上游。据资料记载,在防城港湾的东湾和湾口开展的各1个站位的2个航次(1989年10月和1990年1月)拖网调查中获得包括日本囊对虾、须赤虾、红点黎明蟹(Matutalunaris)和锈斑蟳等7种甲壳类[14]。范航清等(1998)使用定置拦网采集广西英罗港红树林缘潮水中的游泳动物,仅获脊尾白虾、长毛明对虾和三疣梭子蟹(Portunustrituberculatus)等3种甲壳类[26]。何斌源等(2002)使用定置拦网采集广西英罗港红树林潮沟中的游泳动物,捕获11种甲壳类[27]。上述研究所获甲壳类种数远少于本次研究,可能与研究范围相对较窄,设置站位少,生境较单一,温度、盐度范围不够宽广有关。另外,定置拦网方式可能不合适于采集甲壳类,很多甲壳类在退潮后就地潜伏在大拦网范围内的滩涂上。
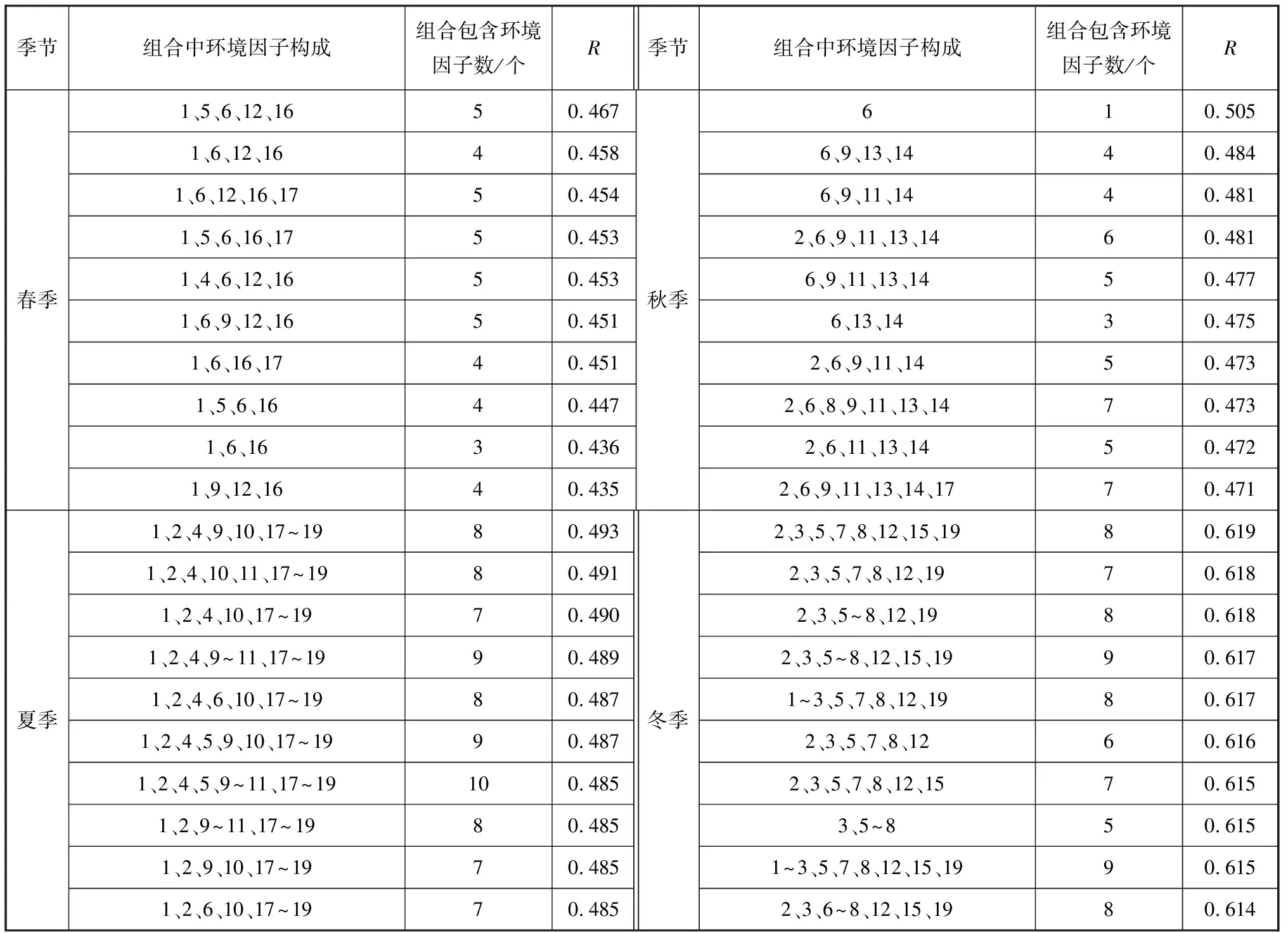
表2 与甲壳类群落结构的相关系数居前10位的环境因子组合
2.7.2 群落优势种结构与资源变化 本研究表明防城港湾甲壳类年均密度和生物量分别为32 201尾/km2和212.290 kg/km2,数量呈现显著的季节变化和空间差异,渔汛期明显以春季为主,湾口高、内湾低,表现出典型的近岸生态类型结构,与本研究调查海域位于河口湾、实测水深不超过15 m的自然条件相符。不同季节的第一优势种不同,但主要优势种基本是表1中的前5位,即亨氏仿对虾、日本囊对虾、钝齿蟳、刀额新对虾和近缘新对虾,相似性百分比分析表明这些种在聚类分组的组内相似性贡献率往往居前,是主要典型种。
防城港湾甲壳类资源特征从属于北部湾总体,但有其独特性,本湾甲壳类优势种结构与北部湾较深海域及琼州海峡明显不同。贾晓平等研究表明北部湾涠洲岛以南海域的秋、冬季生物量接近,分别为12.41、13.76 kg/km2,虾类优势种有中华管鞭虾(Solenoceracrassicornis)、长足鹰爪虾(Trachypenaeuslongipes)、吐露赤虾(Metapenaeopsistoloensis)、刀额仿对虾(Parapenaeopsiscultrirostris)和宽突赤虾(Metapenaeopsispalmensis)等;蟹类优势种有锈斑蟳、逍遥馒头蟹(Calappaphilargius)、日本蟳、香港蟳、纤手梭子蟹(Portunusgracilimanus)和银光梭子蟹等[24]。谢旭等的研究表明,在琼州海峡海域春季优势种仅有葛氏长臂虾(Palaemongravieri),秋季优势种仅有哈氏仿对虾,常见种有周氏新对虾、斑节对虾(Penaeusmonodon)、鹰爪虾和长毛明对虾等4 种[9];该海域的蟹类春季优势种仅日本矶蟹(Pugettianipponensis)1种,秋季种间分布均匀而无优势种,常见种有远海梭子蟹、矛形梭子蟹(Portunushastatoides)、日本蟳、红星梭子蟹(Portunussanguinolentus)、红线黎明蟹(Matutaplanipes)、锈斑蟳和绵蟹(Lauridromiadehaani)等7种[25]。中国毛虾(Aceteschinensis)、哈氏仿对虾、中华管鞭虾等是椒江口海域虾类群落优势种[28],但这些虾类在防城港湾海域仅为偶见。
本研究调查海域甲壳类的平均体重随季节变化明显,春季为5.14 g,夏季为7.74 g,秋季为9.06 g,冬季为7.84 g,先升后降的变化受到动物繁殖生长规律和渔业捕捞活动的综合影响:多数种类在春、夏季繁殖,同时一些种类因索饵需要在一定的海域范围内洄游;由于伏季休渔管控,主要捕捞季节在夏末至冬初,渔汛期间个体较大者被捕捞。渔业生产的相关因素可导致群落结构和数量的年际变化,2010年8月在湾口区布设6个站位的调查结果表明:甲壳类生物量为252.52 kg/km2,虾类、虾蛄类、蟹类的每尾质量分别为3.71、7.03、14.00 g[29]。本研究2016年8月调查表明湾口区的甲壳类生物量为270.61 g/km2,虾类、虾蛄类、蟹类的每尾质量分别为4.66、12.90、17.72 g。2010—2016年,湾口区甲壳类生物量提高了7.2%,虾类、虾蛄类、蟹类的每尾质量分别提高25.6%、83.5%、26.6%,优势种向较大型化发展,越来越严格的休渔管控促使资源在一定程度上得到恢复。
2.7.3 群落结构与环境因子的关系 河口海湾是海陆作用最为剧烈、生态结构最为复杂的区域,同时也是人为干扰最频繁、污染最严重的区域。影响海洋甲壳类群落结构和数量的自然因素复杂,包括水深、盐度、温度、地形、水团甚至全球气候变化[8,28, 30-37]。同时捕捞、围填海、环境污染等人为因素的影响也非常显著[8,38-42],往往导致甲壳类群落结构发生明显演替,资源结构呈现小型化和低值化。
生物-环境分析表明防城港湾甲壳类群落结构与本研究测定的19个环境因子存在或大或小的显著相关关系,不同季节的主要环境因子差异较大,大多数情况下多因子组合具有更高的显著性,反映了环境因子的复杂性和综合性。这些环境因子之中既有不可控的宏观自然因子,也有人类活动导致的污染因子。生物-环境分析中的最大相关系数意味着发现了所测环境因子与生物群落的最佳匹配,环境因子组合对所观察群落结构做出最好的解释;同时,因子少且影响显著的组合显然比较容易解释群落结构[19]。在春季,与群落结构相关性居前的单个环境因子是盐度和水深,这两个因子组合对甲壳类群落结构影响达到显著水平(R=0.434,p<0.05)。春季是防城港湾甲壳类主要种群的繁殖盛期,此时沿岸水团盐度、温度条件满足大多数甲壳类产卵繁殖要求,甲壳类渔获多为幼体,同时春季甲壳类密度远高于其他季节。从密度分布来看,98.2%的亨氏仿对虾、98.2%的须赤虾、82.7%的近缘新对虾集中分布在水较深、盐度高的湾口区,日本囊对虾、刀额新对虾等虽然分布范围广,但也以湾口区相对较多。盐度和水深协同作用可以解释春季甲壳类群落主要优势种的分布格局。
夏季对群落结构影响显著的最少因子组合是水深、水温和总磷组合(R=0.455,p<0.05)。夏季大量淡水进入河口湾导致水体盐度普遍下降,防城河入海最先流经的西湾北部几个站位盐度很低,不利于多数甲壳类生存。河口种刀额新对虾分布范围向外扩展,67.9%的个体集中分布在湾口区;75.4%的钝齿蟳和74.9%的亨氏仿对虾也集中于湾口区;防城港湾夏季实测水温平均达32.8 ℃,水较深、温度较低的湾口区更为适合甲壳类生存。同时,随着夏季生产生活日趋活跃,总磷等污染物更多地从西湾顶部排入,人为因素改变甲壳类生境条件进而影响其群落结构与数量。
盐度单个因子极显著影响秋季甲壳类群落结构(R=0.505,p=0.005),相关系数比其他因子或因子组合高出较多,这种环境因子格局是4个季节仅有的(表2)。盐度呈明显梯度是河口湾的特征,决定了不同渗透压调节能力的甲壳类种群分布格局,河口性强的刀额新对虾96.6%的个体集中分布在盐度较低的西湾中上部;海洋性较强的钝齿蟳和鹰爪虾分别有73.1%和81.4%分布在湾口区高盐站位;亨氏仿对虾、近缘新对虾、日本囊对虾则广布在中等盐度水体且较分散;秋季甲壳类各主要种群表现出符合其生理适应特点的规律性分布,反映了盐度主导的环境因子格局。
冬季透明度、盐度、DO等3因子组合的相关系数较高(R=0.611,p<0.01),很接近该季节最大相关系数0.619(表2);同时相关性最高的单个环境因子DO远高于其他单因子(R=0.536,p<0.05)。适宜的DO范围是海洋生物生存的基本条件,透明度较高有利于甲壳类饵料浮游生物的生长;盐度的梯度分布细化了不同渗透压调节能力的甲壳类种群的适宜生境,河口性种刀额新对虾集中在盐度较低的西湾中上部,亨氏仿对虾和须赤虾多生活在高盐度的湾口区,钝齿蟳则多出现在中等盐度站位。
此外,虽无法定量人为因素的影响,但秋、冬季甲壳类群落种数少、数量低、多样性指数不高,明显地与渔业活动管控程度同频共振,由于伏季休渔管控,捕捞主要集中在夏末至冬初,导致秋、冬季甲壳类群落结构及数量发生较大变化。同时氮磷营养盐、COD等指标一般起着消极作用。因此,必须加强渔业活动的管理和污染物排放控制,积极开展海洋生态修复,才能保证防城港湾渔业资源的可持续利用。
3 结论
在防城港湾调查共获甲壳类60种,其中虾蛄类3种,虾类25种,蟹类32种。暖水性种占60.0%,暖温性种占40.0%。周年优势种有亨氏仿对虾、日本囊对虾、钝齿蟳、刀额新对虾和近缘新对虾等5种,主要种、一般种和少见种分别有6、12、37种。全湾甲壳类年均密度为32 201尾/km2,年均生物量为212.29 kg/km2,季节变化和空间差异显著,在春季形成渔汛。每站种数为2~17种;H′在0.200~3.160之间,平均值为1.847,H′大于3.000的站位仅占2.4%;d在0.200~2.230之间,平均值为1.056;J在0.160~0.988之间,平均值为0.734。聚类分析表明春、夏、冬季群落均分为2组,秋季分为4组,甲壳类群落表现出一定程度的时空异质性。影响群落结构的主要环境因子随季节变化。
猜你喜欢
广西电业(2022年7期)2022-10-13
百科探秘·海底世界(2022年2期)2022-01-26
中国设备工程(2020年7期)2020-06-28
人物画报(2020年1期)2020-01-07
温州大学学报(自然科学版)(2019年2期)2019-06-04
东坡赤壁诗词(2018年6期)2018-12-22
中国-东盟博览(旅游版)(2018年7期)2018-05-14
百科知识(2017年21期)2017-12-05
源流(2015年10期)2015-11-17
医学综述(2015年11期)2015-09-12
