
中生代鞘翅目扁甲总科化石的研究进展
2020-09-14 11:50:48刘振华金建华
环境昆虫学报 2020年4期
刘振华,任 东,金建华,庞 虹*
(1. 中山大学生命科学学院/生态学院,中山大学生物博物馆,生物多样性演化与保护广东普通高校重点实验室,广州 510275; 2. 首都师范大学生命科学学院,昆虫演化与环境变迁重点实验室,北京 100048)

目前,结合分子系统学的研究结果,认为扁甲总科起源于晚三叠纪世至早侏罗纪世(McKennaetal., 2015; Zhangetal., 2018; McKennaetal., 2019),因此对中生代扁甲总科化石的研究不仅有助于研究扁甲总科在中生代的多样性及起源演化问题,也可以帮助我们了解扁甲总科中多样化的生活习性的起源。但是,由于化石仅保存了部分的形态特征,因此更增加扁甲总科化石研究的难度。长期以来关于扁甲总科化石的记录都十分稀少,直到21世纪后琥珀化石研究的兴起才逐渐改变了这一现状。本文对扁甲总科高级阶元的研究进展和中生代扁甲总科化石的研究进行概述。
1 扁甲总科高级阶元的研究
Latreille (1804) 最早以Cucujipes的名字描述了扁甲总科Cucujoidea,但那时还不具有总科、系等高级阶元的概念,也无完善的系统学思想,因此这一名称仅是用来指代一类有相似特征且在外型上比较相似的甲虫。19世纪后期,由于分类学研究方法的发展以及系统学思想的出现,包括外生殖器在内的特征都被用于甲虫分类,也出现了高级阶元的概念。Lameere (1900)、Kolbe (1901) 和Ganglbauer (1903a, 1903b) 均提出了亚目、总科或系的概念,并建立了锤角类Clavicornia这一分类单元,包含有大多数当时已知的扁甲总科类群。Sharp和Muir (1912) 首次基于雄性外生殖器对甲虫进行分类,将它们分成了包含扁甲总科在内的8个总科。直到现在郭公总科Cleroidea、扁甲总科、瓢虫总科Coccinelloidea、拟步甲总科Tenebrionoidea和长蠹总科Bostrichoidea这些类群的雄性外生殖器类型大都仍被称为“扁甲型”生殖器。Böving和Craighead (1931) 首次根据幼虫特征对甲虫进行了划分,将45个科归入到扁甲总科,一些研究者认为这是现代扁甲总科概念的首次出现(Pakaluketal., 1994)。但是仅依靠幼虫特征进行的分类系统存在缺陷,里面包括了部分现在不属于扁甲系的类群,例如扁股花蚤科Eucinetidae,伪郭公科Derodontidae和长蠹科Bostrichidae。Crowson (1955)在其《鞘翅目科的自然分类》一书中提出的扁甲总科的概念已具有系统发育学的思想,虽然在组成上仍与Böving和Craighead (1931) 提出的观念有许多相似,但不包括上述的现有分类系统不属于扁甲系的类群。另外,Crowson将扁甲总科分为锤角类Clavicornia和异跗类Heteromera,其中锤角类包含了广义上扁甲总科Cucujoidea s.l.的大部分类群,因此也被McElrathetal. (2015) 视为现在扁甲总科分类阶元的基础,而后者则为现在的拟步甲总科。但同时Crowson也指出由于缺乏强有力的形态学证据,应将扁甲总科与相近的类群如郭公总科、叶甲总科Chrysomeloidea和象甲总科Cuculionoidea相区分。Crowson关注到总科内各科之间的划分存在许多问题,提出了厘清扁甲总科与其它总科之间关系、扁甲总科内各科间的关系的艰巨性。Crowson (1955) 进一步指出在锤角类中存在包含皮坚甲科Cerylonidae在内的一个亚群,它们都具有4-4-4或3-3-3的跗式以及特别的后翅翅脉模式等一些特征,被称为皮坚甲系Cerylonid series。另外宽颚扁甲科Prostomidae在Crowson (1955, 1967) 的研究中仍被视为扁甲科Cucujidae中的一个亚科,但Crowson (1967) 指出基于后翅翅脉、外生殖器以及幼虫形态,该类群应该与异跗类更近而非扁甲科,这一观点也被之后的研究人员所采用(Abdullah, 1974; Lawrence, 1977)。圆蕈甲科Ciidae和长酪甲科Phycosecidae也分别从锤角类移至异跗类(Crowson, 1960)和郭公总科(Crowson, 1964)。毛蕈甲科Biphyllidae和小花甲科Byturidae的分类学地位尤为不确定,大多数情况下它们被视为扁甲总科中的锤角类(Crowson, 1955, 1967, 1981; Lawrence, 1982),但外生殖器的形态以及转节和腿节相接的模式却更接近异跗类(Crowson, 1960; Watt, 1974; Lawrence, 1977),同时它们与郭公总科的相似也时常被提及(Crowson, 1955)。Lawrence和Newton (1982) 首次以拟步甲总科来指代异跗类,而扁甲总科则专指锤角类,这一观点被许多学者接受(Pakaluketal., 1994; Bouchardetal., 2011)。此后,Sen Gupta和Crowson (1966, 1969) 对南半球的扁甲总科进行了一系列研究,设立了澳洲蕈甲科、鸟巢甲科和皮扁甲科Phloeostichidae。而Lawrence (1991) 基于幼虫特征将霍巴特甲科Hobartiidae从澳洲蕈甲科中独立出来,之后Gimmeletal. (2009) 基于前人的研究首次确立新西兰特有类群伪姬花甲科Cylaxyridae有效的科级地位。Leschenetal. (2005) 利用基于形态特征的系统发育学研究方法,主要针对当时的皮扁甲科进行了形态系统发育学研究,把蚧蕈甲科Agapythidae,皮蕈甲科Priasilphidae,澳洲扁甲科Myraboliidae和伪角甲科Tasmosalpingidae 4个科独立出来,并且在该研究中将扁甲总科中除皮坚甲系之外的类群称为“Basal Cucujoid”。
自21世纪以来,分子生物学技术对生物系统发育学研究产生了巨大的影响,尤其是在高级阶元的分类研究上。Huntetal. (2007) 关于鞘翅目的分子系统学研究显示小花甲科和毛蕈甲科都应属于郭公总科,得到了许多研究者的支持(Bocaketal., 2014; McElrathetal., 2015; Robertsonetal., 2015)。该研究显示“皮坚甲科系”并不属于扁甲总科,但由于系统发育树中整个扁甲总科的位置并未得到很好的解释,因此并没有对此作出进一步的分析。Clineetal. (2014) 的研究将Cybocephalinae亚科从露尾甲科Nitidulidae中移出,独立成方头甲科。而McElrathetal. (2015) 和Robertsonetal. (2015) 的研究均支持将皮跳甲科Propalticidae降为扁谷盗科Laemophloeidae中一亚科,而后者则首次将原先的“皮坚甲系”提升至瓢虫总科,得到多项研究结果的支持(McKennaetal., 2015; Zhangetal., 2018)。最近,McKennaetal. (2019) 主要基于转录组数据的研究再一次对扁甲总科的系统提出了挑战,他们的研究显示扁甲总科并非单系,而是分为3个单独的支系,其中澳洲蕈甲和大蕈甲为一支系,蜡斑甲科Helotidae、姬蕈甲科Sphindidae、原扁甲科Protocucujidae、出尾扁甲科、短甲科Smicripidae、小露尾甲科和露尾甲科为一支系,剩余其它科则被还原为叶甲总科和象甲总科的姐妹群。该研究结果由于其基部节点的支持率并不高,因此还有待进一步探讨。综上所述,目前扁甲总科共包含了25个现生科(表1),但该分类系统随着未来研究的深入仍有可能发生变化。
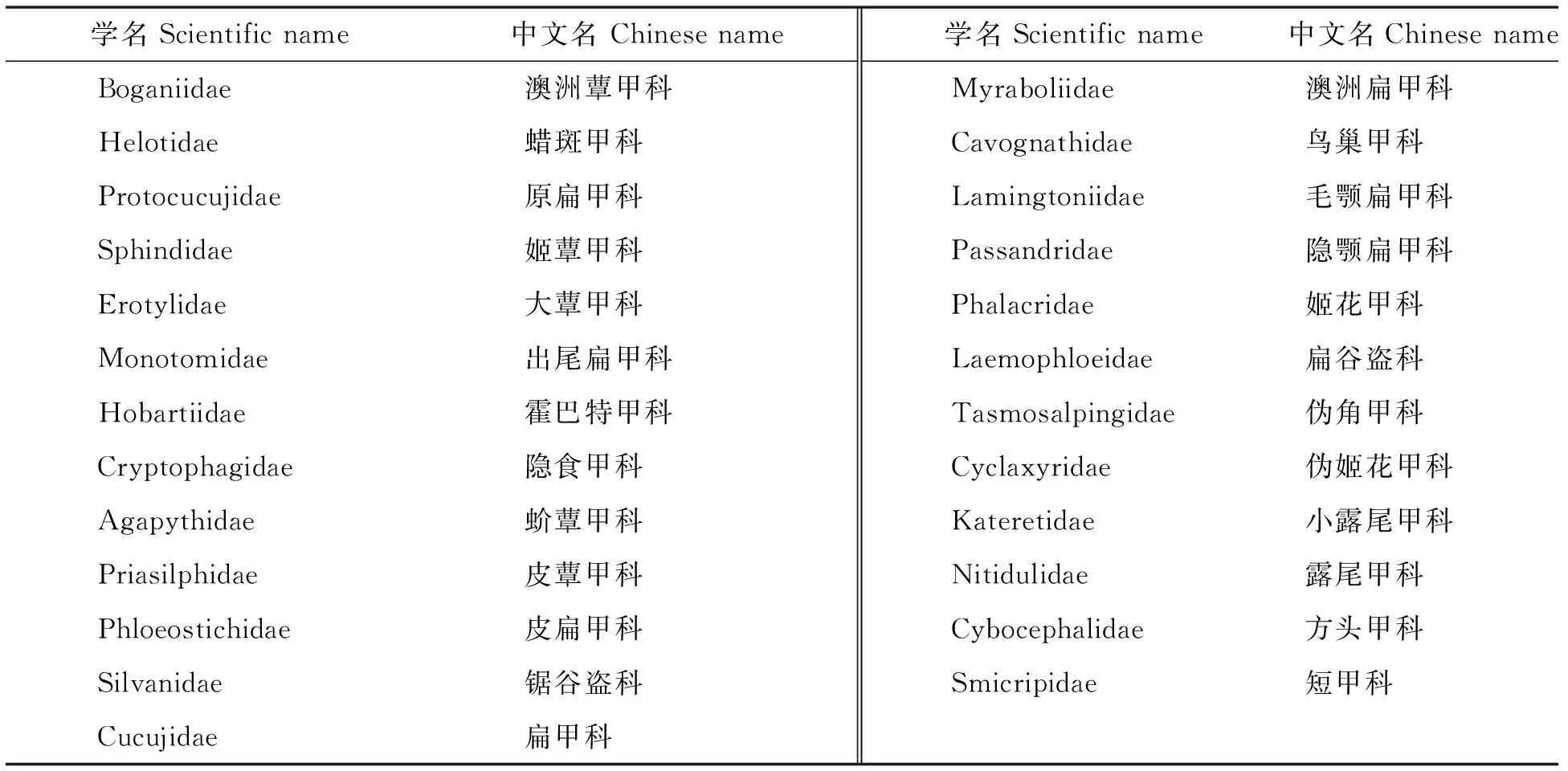
表1 扁甲总科各科学名-中文名对照表Table 1 Contrast of Scientific-Chinese family names in Cucujoidea
分子系统学研究的发展同时也推动了另外一项研究的进行,即基于分子系统发育树和化石证据的分歧时间估算。其中大多数的研究显示扁甲总科的起源时间应该为早侏罗世至中侏罗世(Huntetal., 2007; McKennaetal., 2015; Zhangetal., 2018),而Toussaintetal. (2017) 利用McKennaetal. (2015) 的分子数据和不同的化石证据的研究结果将这一时间推前至早三叠世,McKennaetal. (2019) 的研究虽然也将扁甲总科的起源时间往前推进,晚于Toussaintetal. (2017) 的结果,为晚三叠世。由此也可以看出化石的研究对分歧时间的估算有着极其关键的作用,化石鉴定的准确与否决定着起源时间推测的可靠性。
2 中生代扁甲总科化石的研究
甲虫作为最古老的全变态发育昆虫之一,其化石的研究长期吸引了众多研究人员的注意。但甲虫的许多鉴定特征,比如跗节数量及形态、各足基节窝的形态等在许多化石中都难以观察,有些化石仅保存有鞘翅。因此其分类鉴定工作存在着很大的难度,特别是对于如扁甲总科这类在形态上相似的类群。目前最古老的甲虫是描述自德国早二叠世地层的ColeopsisarchaicaKirejtshuk, Poschmann and Nel (2014),然而最古老的扁甲总科化石发现自早侏罗世。
2.1 侏罗纪的扁甲总科化石

图1 Nitidulites argoviensis Heer, 1865线条图(引自Heer, 1865)Fig. 1 Illustration of Nitidulites argoviensis Heer, 1865 (cited from Heer, 1865)
2.2 白垩纪的扁甲总科化石

表2 中生代扁甲总科化石名录Table 2 Checklist of Mesozoic Cucujoidea

图2 A,Palaeoboganium jurassicum Liu, , Lawrence, Ren and Pang, 2018模式标本(引自Liu et al., 2018);B,Metahelotella monochromata Liu, , Ren and Pang, 2019模式标本(引自Liu, , Ren et al., 2019);C,Lenax karenae Liu, Tihelka, McElrath and Yamamoto, 2020模式标本(Liu et al., 2020);D-E,Pleuroceratos burmiticus Poinar and Kirejtshuk, 2008模式标本(引自Liu, , Wang et al., 2019)

续表2 Continued table 2
进入新生代后,有更多扁甲总科的化石在世界各地的不同地层之中被发现,其中更有两个新科被描述,分别为发现于中国始新世抚顺琥珀的中国锯谷盗科Sinisilvanidae Hong(洪友崇,2002)和来自始新世波罗的海琥珀的Wabbelidae Alekseev(Alekseev, 2017)。
2.3 中生代扁甲总科化石的系统学研究
3 中生代扁甲总科化石研究中存在的问题与展望
3.1 中生代扁甲总科化石研究中存在的问题
(1)扁甲总科化石鉴定困难。正如之前提到的,扁甲总科形态特征鉴定的困难给化石的研究带来了困难。一方面是中生代扁甲总科的化石记录较为稀少,特别是印痕化石所提供的形态学特征信息较少;另一方面部分化石鉴定的准确性存在一定问题。根据中生代扁甲总科化石的统计数据(图3)可以看到,印痕化石不仅物种记录少于琥珀化石,其科属的丰富度更是远不及后者。而化石鉴定的不准确不仅是由于化石保存下来的特征较少,也与化石研究的阶段相关。比如Heer (1865) 在描述NitidulitesargoviensisHeer和Nitidulitesbellus(Heer) 时将后者放在了吉丁科内,然而根据原始的手绘图我们甚至无法对它们的系统地位作出任何的推测(图1)。但即使在琥珀化石中也出现鉴定错误现象,比如Poinar and Kirejtshuk (2008) 在描述缅甸琥珀中的PleuroceratosburmiticusPoinar and Kirejtshuk(图2-D)时根据其与锯谷盗科OryzaephilusGanglbauer外形的相似性把它放在了锯谷盗科中的锯谷盗亚科。根据该种甲虫前足基节窝基转片外露(图2-E)这一特征不同于任何锯谷盗科,Liuetal. (2019) 将其从锯谷盗科中移除,视为一科级地位不确定的甲虫。事实上,我们在研究过程中检视到不少扁甲总科的印痕化石或者琥珀化石材料,但其中一部分无法确定它们的科级地位。这其中一部分原因是化石中能观察到的特征有限,时常会缺失比较关键的特征;另外现生扁甲总科中很多科都没有很好的独有衍征,并且部分重要特征存在于外生殖器这类难以在化石中保存下来的部位,这就加大了化石材料鉴定的难度。
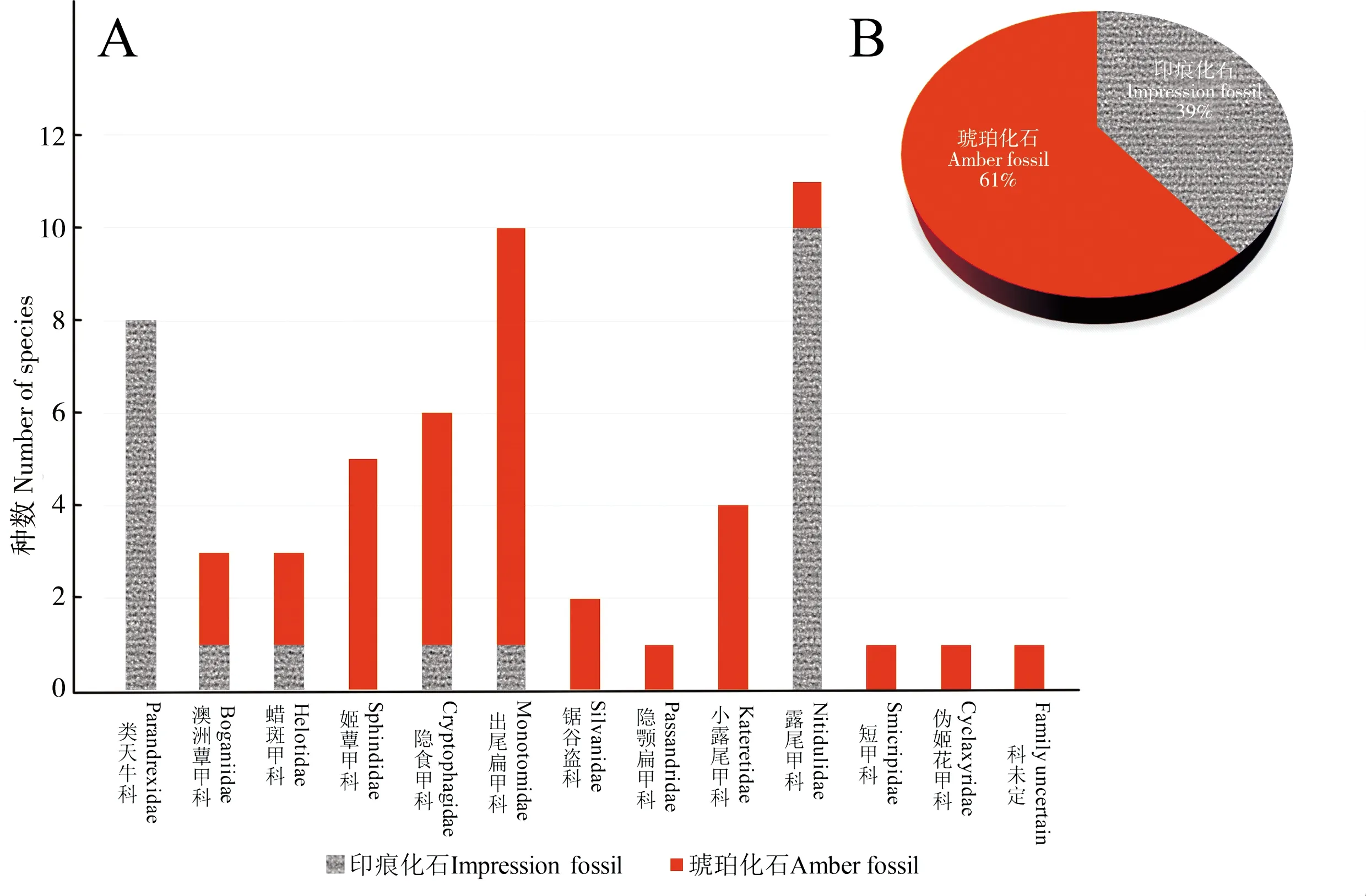
图3 中生代扁甲总科化石统计图Fig.3 Diagram of Mesozoic Cucujoidea注:A,示各科中琥珀化石及印痕化石的组成;B,示总体琥珀化石及印痕化石物种数的比例。Note: A, composition of amber fossils and impression fossils in each family; B, ratio of amber fossils and impression fossils in total.
(2)缺少现生形态系统发育研究。对于高级阶元位置不明的化石种类,使用基于形态特征的系统发育学研究方法是一个有效的解决途径。然而到目前为止,仅有Leschenetal. (2005) 和Lawrenceetal. (2011) 对扁甲总科进行了较为全面的形态学系统发育研究。但前者的研究重点在于解决当时Phloeostichidae的分类学问题,后者则为整个鞘翅目的系统发育学研究,并且结果和最近的分子系统学研究的结果存在较大的差异。关于形态系统发育方法用于化石的研究,目前可见于Liuetal. (2018) 和Caietal. (2018), Jinetal. (2019) 以及Gimmeletal. (2019) 分别对澳洲蕈甲科、隐颚扁甲科和伪姬花甲科的化石进行的系统发育学分析。
(3)部分类群化石记录地层的单一性。从化石记录上可以看到,扁甲总科中许多科在中生代的化石记录比较稀少,不少都仅发现于一个地层。比如姬蕈甲科仅发现于黎巴嫩琥珀中,锯谷盗科、隐颚扁甲科、短甲科和伪姬花甲科则仅发现于缅甸琥珀中,而这样的局限性可能影响相关的古生物地理学推测。Wuetal. (2018) 描述了缅甸琥珀中的Electroxyracretacea(Wu),首次在新西兰之外发现伪姬花甲科,推测该科甲虫在白垩纪时期很可能广泛分布于冈瓦纳古陆。然而Gimmeletal. (2019) 将描述于波罗的海琥珀的姬花甲科化石属NeolitochropusLyubarsky and Perkovsky (Lyubarsky and Perkovsky, 2016) 移至伪姬花甲科,推测伪姬花甲科曾经可能在盘古大陆广泛分布,并在欧洲地区至少存活至新生代。
3.2 中生代扁甲总科化石研究的展望
长期以来,甲虫化石的研究主要集中在被认为是较为原始的类群原鞘亚目和肉食亚目(Crowson, 1962; Kirejtshuketal., 2010; Tanetal., 2012; Beuteletal., 2013; Kirejtshuketal., 2016; Jarzembowskietal., 2017)以及二叠系比较古老的地层(Ponomarenko, 2004; Ponomarenkoetal., 2014; Yanetal., 2018)。扁甲总科化石的研究相对来说比较稀少,直到21世纪之后才有越来越多的中生代扁甲总科化石被描述。从化石记录中可以看到,超过70%的中生代扁甲总科化石是在21世纪甚至是近10年才被发现,只有3个科在20世纪便有了化石记录。再加上扁甲总科中还有超过一半的类群无化石记录,因此对于中生代扁甲总科化石的研究一方面仍需要大量的基础分类学研究,通过新的化石记录来了解中生代扁甲总科的多样性起源;另一方面还需要更多地使用系统发育学的研究方法来进行研究,这不仅有助于更准确的鉴定化石,也可以帮助探究化石种与现生类群的系统关系,进而为扁甲总科的起源与演化以及生物地理学研究提供依据。
前面提到扁甲总科中有大量仅发现分布在南半球澳大利亚、南美以及新西兰的类群,最新的研究显示扁甲总科的起源时间为晚三叠世至早侏罗世(Zhangetal., 2018; McKennaetal., 2019);而地质学研究则显示盘古大陆的分离发生自 1.8~1.6亿年前,冈瓦纳古陆最早的分离大概发生在侏罗纪约1.65~1.6亿年前,并在之后发生了一系列的板块运动形成现在的局势(Sanmartin and Ronquist, 2004)。因此扁甲总科目前的分布格局很可能与板块运动有着密切的联系。更多更古老的化石记录将有助于准确的推测出扁甲总科中各个科乃至亚科的起源时间,结合古地质学的研究推测扁甲总科的演化历史,以及解释其中南半球类群分布模式的成因。
猜你喜欢
少年博览·小学低年级(2023年9期)2023-10-09 09:51:46
幼儿画刊(2023年3期)2023-05-26 05:39:28
小哥白尼(野生动物)(2021年4期)2021-07-29 08:15:46
小哥白尼(野生动物)(2021年1期)2021-07-16 08:02:50
银潮(2020年11期)2020-12-04 12:42:52
江苏农业科学(2019年5期)2019-09-02 14:01:46
广东农业科学(2017年5期)2017-08-29 10:37:31
童话世界(2017年8期)2017-05-04 04:08:36
健康必读(2016年7期)2016-08-30 04:49:04
中国火炬(2015年3期)2015-07-31 17:39:19
