
布鲁氏菌BP26蛋白对鼠源树突状细胞分化及抗原提呈作用
2020-09-10 04:01曹小安刘永生周建华张广林孙晶晶李玲霞赵永婕胡永浩
江苏农业学报 2020年4期
徐 龙, 曹小安, 刘永生, 周建华, 张广林, 孙晶晶, 李玲霞, 赵永婕,胡永浩
(1.甘肃农业大学动物医学院,甘肃 兰州 730070; 2.中国农业科学院兰州兽医研究所/家畜疫病病原生物学国家重点实验室,甘肃 兰州 730070)
布鲁氏菌病(Brucellosis)是由布鲁氏菌(Brucella)感染引起的人畜共患传染病,具有流行广泛、危害极大的特点。近年来,布鲁氏菌给中国的公共卫生与安全带来了新挑战。由于布鲁氏菌没有明显的外毒素,其依靠外膜蛋白、脂多糖及Ⅳ型分泌系统发挥毒力并通过抑制溶酶体的相关功能抵抗细胞凋亡和逃离细胞死亡与降解。Brucella在宿主发挥细胞免疫和体液免疫时,具有逃避能力。Brucella在侵入机体后,主要侵染胚胎细胞及巨噬细胞,胞内生存和繁殖是其主要致病机理,其免疫机制在逃离宿主的免疫系统时发挥了重要作用[1]。
诸如OMP10、OMP19和BP26等外膜蛋白刺激BM-DCs(小鼠分离髓源树突状细胞)后,BM-DCs表面共刺激分子和MHC分子的表达及培养上清液中IL-12、IL-10、INF-γ、TNF-α的分泌均显著增加。而OMP25和OMP31刺激BM-DCs后会显著抑制部分表面分子的表达及炎性细胞因子IL-12、IL-6、INF-γ、TNF-α的分泌[2]。Dohmer等[3]也发现了其他种类的分泌蛋白质,但对这些蛋白质的功能仍处于探索阶段,具体机制也尚未研究透彻。刘景福等[4]发现BP26蛋白在BP26免疫模型上能够形成明显的迟发型变态反应。王勇等[5]发现重组蛋白BP26作为检测抗原建立的间接ELISA方法效果较为理想,可适用于临床布鲁氏菌感染的检测。王芳等[6]发现基于BP26建立的间接ELISA检测方法在检测Brucella时不能对细菌种属及其来源进行有效区分,通过检测BP26抗体不能对羊布鲁氏菌病进行有效区分和诊断。对小鼠接种OMP或L7/L12抗原的DNA疫苗或重组蛋白质可有效对抗布鲁氏菌有毒株的攻击感染。Saadi等[7]从OMP31、BP26、BLS、DnaK和L7/L12蛋白中筛选了5个新的T细胞表位,并基于上述蛋白质制成多表位肽疫苗使其稳定有效表达,在未来有可能用于预防或治疗布鲁氏菌病。
虽然BP26能够产生强力的抗体,但有效的抗原在细菌侵染宿主细胞时发挥的作用和分子机制尚不明确。本试验选取布鲁氏菌外膜蛋白基因BP26进行全基因扩增,并成功将其构建到pET-30a质粒中进行稳定表达,获得了重组质粒pET-30a-BP26。说明该蛋白质可在大肠杆菌内稳定表达。利用分子生物学的方法对BP26蛋白进行分析,获得了BP26蛋白的相关特性的基本信息,为未来研究布鲁氏菌侵袭宿主后BP26蛋白会通过何种信号通路对机体产生影响及相关调控机制奠定基础。
1 材料与方法
1.1 试验材料
1.1.1 菌株和质粒 布鲁氏菌阳性基因组、质粒pET-30a(+)为中国农业科学院兰州兽医研究所草食动物细菌病创新团队保存。大肠杆菌DH5α和BL21(DE3)感受态细胞购自北京全式金生物技术有限公司。
1.1.2 血清与抗体 羊布鲁氏菌阳性血清为中国农业科学院兰州兽医研究所草食动物细菌病创新团队保存,辣根过氧化物酶(HRP)标记的兔抗羊IgG购自赛默飞世尔科技(中国)有限公司。
1.1.3 主要生化试剂 2×EasyTaq© PCR SuperMix (+dye )、限制性内切酶EcoRⅠ、SalⅠ、T4 DNA Ligase、胶回收试剂盒、质粒小提试剂盒、IPTG购自宝生物工程(大连)有限公司,卡那霉素、30% Acr制胶液、RPMI1640细胞培养基、PBS(pH=7.2)购自北京索莱宝科技有限公司,High Affinity Ni-Charged Resin FF购自南京金斯瑞生物科技有限公司,Tryptone、Yeast Extract购自OXOID公司。
1.1.4 试验动物、病毒及试剂 28~42日龄BALB/C小鼠由中国农业科学院兰州兽医研究所动物中心提供,所有动物试验按照单位相关伦理要求进行。
胎牛血清购自美国Glbco公司。重组小鼠粒细胞-巨噬细胞集落刺激因子(rmGM-CSF)和重组小鼠白介素-4(rmIL-4)购自美国PeproTch公司。单克隆抗体为兔抗鼠的PE-CDllC、FITC-MHCⅡ、FITC-CD86、APC-CD80、APC-CD40,均购自美国BD Biosciences公司。阳性对照为布鲁氏菌脂多糖(LPS),购自Sigma公司。
1.2 方法
1.2.1 引物合成 使用Primer Premier5.0筛选出最优引物,上下游引物两端添加保护性碱基并分别引入EcoR Ⅰ和SalⅠ 2个酶切位点,并对密码子进行适当优化,引物:BP26-F:5′-CCGGAATTCGCCACCATGCAGGAGAATCAGATGACG-3′,BP26-R:5′-CCGGTCGACTTAATGATGATGATGATGATGCTTGATTTCAAAAACGAC-3′),酶切位点以下划线标出。设计完成的引物由北京擎科新业生物技术有限公司合成,扩增片段长度为666 bp。
1.2.2 布鲁氏菌BP26基因的PCR扩增及克隆 以布鲁氏菌基因组作DNA模板,扩增体系参照说明书进行配置。采用2×EasyTaq©PCR SuperMix(+dye)和合成的引物在56 ℃退火温度下扩增获得BP26目的片段,将扩增后的产物在1.15%琼脂糖凝胶上电泳,参照胶回收试剂盒说明书对切取的目的条带进行回收。用EcoRⅠ和SalⅠ对回收后的目的片段与pET-30a载体质粒在37 ℃水浴锅中双酶切4~5 h后回收。 用T4连接酶体系16 ℃过夜连接线性化酶切的质粒和目的基因产物。将全部连接产物转入DH5α中,冰浴30 min后42 ℃ 热激90 s,迅速转移至冰上放置3 min。在含200 mg/L的Kan+抗性固体培养基上培养,37 ℃ 倒置过夜培养后挑取阳性克隆的菌落小摇后进行PCR扩增及酶切鉴定。将鉴定成功的质粒送至北京擎科新业生物技术有限公司测序。
1.2.3 布鲁氏菌BP26重组蛋白的表达及纯化 将构建成功的质粒转入BL21中保存,挑取单个阳性克隆的菌落接种至5 ml含Kan抗性液体LB中37 ℃过夜培养。吸取2 ml培养的菌液至200 ml含Kan抗性液体LB中,37 ℃,200 r/min培养。当菌液OD600值达到0.6~0.8时,设置诱导温度为16 ℃、25 ℃、37 ℃ 3个梯度,设置IPTG的终浓度分别为1.00×10-3mol/L、5.00×10-4mol/L、2.50×10-4mol/L、1.25×10-4mol/L、1.00×10-4mmol/L 5个梯度,设置诱导时间为3 h、4 h、5 h、6 h 4个梯度。各设置条件诱导结束后收获菌液,8 000 r/min,4 ℃,离心30 min,弃上清液,用少许0.01 mol/L的PBS再次悬浮沉淀后用同等条件离心,收集沉淀。用Lysis equilibration buffer (LE Buffer)对沉淀进行悬浮后,超声破碎仪裂解,超声5 s,间歇2 s,工作时间为1 h。破碎结束后以上述同等条件离心分离上清液和沉淀,分别取上清液与沉淀40 μl和10 μl, 5×SDS-PAGE Loading Buffer充分混匀煮沸10 min后进行SDS-PAGE,完成后使用考马斯亮蓝染色液对蛋白质胶室温染色4 h后,再以脱色液充分脱色,分析重组蛋白质BP26的表达水平。
将成功鉴定的布鲁氏菌BP26蛋白表达液装入经LE Buffer平衡后的His-Tag Ni柱中,收集流穿液后以LE Buffer洗脱4个柱体积后,以含20 mmol/L浓度咪唑LE Buffer为起点对柱内的沉淀进行洗脱,柱底端口液体流速控制在1.0~1.5 ml/min。含20 mmol/L浓度咪唑LE Buffer洗涤5个柱体积, 含40 mmol/L浓度咪唑LE Buffer洗涤3个柱体积,含60 mmol/L浓度咪唑LE Buffer、含160 mmol/L浓度咪唑LE Buffer各洗涤1个柱体积,含250 mmol/L浓度咪唑LE Buffer洗脱2个柱体积,含20 mmol/L、40 mmol/L、60 mmol/L、160 mmol/L浓度咪唑LE Buffer流出液各收集2 ml,含250 mmol/L浓度咪唑LE Buffer流出液全部收集,纯化获得试验所需的目的蛋白质。
1.2.4 布鲁氏菌BP26重组蛋白的Western-Blot分析 参考文献[8]中的方法,将稀释液调整为10倍稀释的Casein封闭液,优化一抗孵育时间为1 h,二抗孵育时间为45 min,对BP26蛋白进行Western-Blot分析。
1.2.5 小鼠髓源树突状细胞的体外诱导培养 选取4只SPF级的42~56日龄BALB/c雌性小鼠,参照文献[9]分离小鼠髓源树突状细胞(BM-DCs)。参考文献[10]中的方法用rmGM-CSF和rmIL-4刺激,使分离制备的骨髓原代细胞分化为未成熟DCs。DCs培养至第6 d,调整BP26蛋白终浓度为100 μg/ml刺激DCs细胞;调整LPS 终浓度为100 ng/ml刺激DCs细胞,并将该组选作阳性对照,开展后续试验。
1.2.6 布鲁氏菌BP26蛋白刺激鼠源树突状细胞 设置LPS与重组 BP26蛋白分别刺激BM-DCs 24 h后,收集DCs,1 500 r/min离心7 min,收集上清液进行与介导细胞炎症相关细胞因子的检测。BM-DCs沉淀用含有5%胎牛血清、pH为7.2的PBS悬浮,对DCs活性以PE-CDllC抗体4 ℃ 避光染色30 min后经流式细胞仪进行分析。DCs共刺激分子表达分析: 设置PE-CD11C、APC-CD40、FITC-MHCⅡ三色共染为试验组一,设置PE-CD11C、APC-CD80、FITC-CD86为试验组二。同时设置同型对照和单染管,抗体4 ℃染色30 min,加入1 ml含有5%胎牛血清的PBS,1 500 r/min,7 min离心收集DCs。上述PBS重悬细胞,用FAC SC alibur流式细胞仪进行检测,FlowJo软件进行数据分析。
1.2.7 参与炎症反应的细胞因子检测 将BM-DCs沉淀以Trzol法提取细胞RNA并测定其浓度,使用Qiagen Omniscript RT Kit将RNA反转录为cDNA。采用2×SYBR Green Abstart PCR Mix,以cDNA为模板,参考文献[11]设计需要检测的细胞因子引物,各体系与扩增条件参照说明书。试验设置3个独立重复,扩增结束后记录Ct值 。
1.3 统计学数据分析
采用FlowJo V10软件对经流式细胞仪检测获得的结果进行分析,采用统计软件Graphpad Prism 8进行绘图,采用SPSS V25.0软件对试验组和对照组的流式数据与Ct值进行差异显著性分析,以P<0.05为差异显著,P<0.01为差异极显著。
2 结果与分析
2.1 表达质粒的鉴定
基于挑取的阳性克隆菌落进行PCR扩增,并以挑取的菌落培养的菌液作为DNA模板再次进行PCR扩增,2次均扩增出666 bp大小的条带。参照质粒小提试剂盒使用说明书提取菌液质粒,并用EcoRⅠ和SalⅠ将重组表达质粒pET-30a-BP26双酶切后琼脂糖电泳,分别得到大小正确的目的片段和载体片段(图1)。

M:2 000 DNA marker ;1:质粒pET-30a-BP26酶切产物。图1 表达载体双酶切图谱Fig.1 Restriction enzyme map of expression vector
2.2 BP26蛋白的重组表达
对纯化后的BP26蛋白进行SDS-PAGE电泳后经卡马斯亮蓝染色后脱色,最佳诱导条件为:温度37 ℃,IPTG浓度5×10-4mol/L,时间5 h时。诱导后可在30 000左右出现条带(图2),与预期条带大小最符合,条带最为清晰,证明该条件下BP26蛋白在BL21(DE3)中的表达量最高。

M:蛋白质marker;1:pET-30a-BP26上清液;2:pET-30a-BP26沉淀。图2 SDS-PAGE鉴定pET-30a-BP26表达结果Fig.2 Expression of pET-30a-BP26 by SDS-PAGE
将纯化诱导表达上清液中的BP26蛋白的各浓度咪唑洗脱液经SDS-PAGE鉴定分析,发现在泳道1有BP26蛋白和大量杂蛋白质出现,证明诱导表达上清中的蛋白质并未完全与High Affinity Ni-Charged Resin FF结合;泳道2出现了与泳道1相同的情况,证明LE Buffer据有将蛋白质从高亲和力树脂上洗脱下来的能力,泳道3~6并未出现明显的蛋白质条带,但是从泳道7、泳道8可以观察出BP26蛋白的表达量较多,而杂蛋白质条带几乎没有。说明20~160 mmol/L浓度咪唑洗脱液可以充分将杂蛋白质从亲和树脂上洗脱下来,洗脱效果最佳的咪唑浓度为40 mmol/L(图3)。
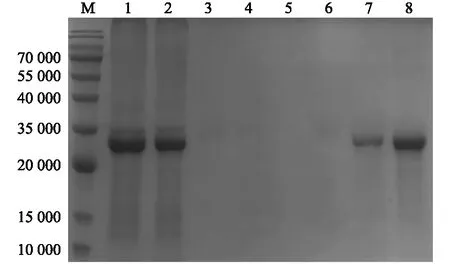
M:蛋白质marker;1:BP26流穿液;2:LE Buffer洗涤后结果;3:含20 mmol/L 咪唑LE Buffer洗涤结果;4:含40 mmol/L 咪唑LE Buffer洗涤结果 5:含60 mmol/L咪唑LE Buffer洗涤结果;6:含160 mmol/L咪唑LE Buffer洗涤结果 ;7、8:含250 mmol/L咪唑LE Buffer洗脱结果。图3 BP26蛋白纯化电泳检测Fig.3 Electrophoresis detection of BP26 protein
2.3 BP26蛋白Western blotting检测结果
表达产物经SDS-PAGE电泳后转印到0.45 μm PVDF膜上,封闭液作用后、经一抗、二抗孵育后ECL显色,在30 000左右出现目的条带(图4),说明表达产物的反应原性良好,符合后续试验要求。

M:蛋白marker;1、2: pET-30a- BP26 (+)。图4 布鲁氏菌BP26蛋白Western-Blot结果Fig.4 Western-blot results of Brucella BP26 protein
2.4 BP26蛋白基本性质分析
ExpASy在线软件分析结果显示,BP26蛋白总长约为250个氨基酸,相对分子质量大小为26 550,等电点为6.39;带正、负电荷氨基酸残基数均为24。BP26蛋白在280 nm处的摩尔消光系数为5 960 mol/L;BP26蛋白N端为Met,在哺乳动物体内半衰期约为30 h,在酵母体内半衰期大于20 h,在大肠杆菌体内半衰期大于10 h。
BP26蛋白在溶液中的不稳定指数为27.93,推测BP26蛋白性质稳定。BP26蛋白的脂肪族指数为92.16,亲水性平均系数为-0.030,BP26属跨膜蛋白且其外膜蛋白质具有较高的亲水性,属亲水蛋白质。对BP26蛋白的二级结构进行预测,发现α螺旋比例为41.20%,β转角比例为4.80%,无规则卷曲比例为36.80%,延长链比例为17.20%,可见BP26蛋白的二级结构以α螺旋为主(图5)。BP26蛋白的信号肽区域为第1~28位氨基酸,信号肽剪切位点在28~29位氨基酸之间。对BP26蛋白的氨基酸序列中的抗原表位进一步进行分析,数据显示序列中存在8个潜在的抗原表位,平均抗原倾向性系数为1.016 4。

图5 布鲁氏菌BP26蛋白二级结构预测Fig.5 Secondary structure prediction of Brucella BP26 protein
2.5 BM-DCs体外诱导培养及纯度检测
在细胞因子rmGM-CSF和rmIL-4的共同作用下,4只BALB/C鼠分离的BM-DCs平均含量可达到1 ml 1.3×106细胞,符合试验要求。倒置显微镜观察DCs的形态特征及生长状况。在培养24 h后,可观察到DCs体积较小,为均一的圆形,部分贴壁生长。至第3 d可观察到DCs集落群并且DCs体积较第1 d的体积大。培养至第6 d,部分DCs脱离集落群,呈现不规则形态并形成类似于树杈状的突起(图6)。将培养第6 d的DCs用PE- CD11C染色标记后用流式细胞仪分析DCs活性,结果显示:分离制备的DCs活性可达到85%以上(图7),符合试验要求,可用于后续试验。

A:骨髓原代细胞培养24 h;B:诱导第3 d;C,D:诱导第6 d ;A,C,D:放大×20;B:放大×40。图6 用含rmGM-SF和rmIL-4完全培养基不同时间诱导的细胞形态Fig.6 Induction of cell morphology at different time with complete medium containing rmGM-SF and rmIL-4

图7 DCs活性Fig.7 Activity detection of dendritic cells
2.6 布鲁氏菌BP26蛋白对DCs成熟及细胞因子表达
如图8所示,LPS组与DC对照组相比,经LPS刺激作用后,DCs的成熟呈促进趋势。同时发现在BP26刺激BM-DCs后,试验组设置的CD40、MHC-Ⅱ、CD86和CD80的双染区域比值与空白DC组对比后,有明显的上升趋势,故能判定布鲁氏菌BP26蛋白对BM-DCs的成熟具有促进作用。SPSS软件分析结果显示,LPS与BP26虽然对BM-DCs的成熟均有促进作用,但APC-CD40、APC-CD80、FITC-CD86组中差异不明显,而在FITC-MHCⅡ组中呈现出较明显的差异。

图8 流式细胞术检测BP26和LPS处理后DC表面共刺激分子的表达Fig.8 The expression of costimulatory molecules on dendritic cells treated with BP26 and LPS by flow cytometry
2.7 参与DCs炎症反应细胞因子的检测
如图9所示,LPS组与DC对照组相比,经LPS刺激作用后,TNF-α的表达量差异显著(P<0.05)。同时发现在BP26刺激BM-DCs后,IL-10的表达量与DC对照组相比呈下降趋势且差异极显著(P<0.01),IL-12的表达量与DC对照组相比呈现下降趋势且差异显著(P<0.05)。分析结果说明,布鲁氏菌BP26蛋白能够引起细胞的炎症反应,促进炎性细胞因子的释放。
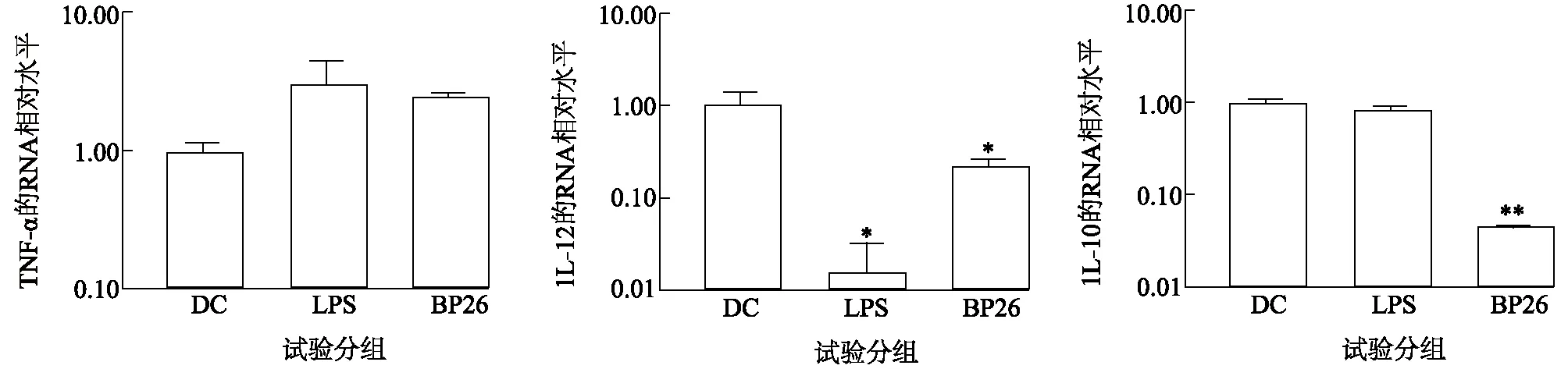
图9 参与BM-DCs炎症反应细胞因子显著差异性分析Fig.9 Analysis of significant differences in cytokines involved in BM-DCs inflammatory response
3 讨 论
本研究获得了高效可溶的BP26重组蛋白,与预测结果相一致。纯化后刺激BM-DCs的研究结果表明,BP26蛋白能促进BM-DCs分化成熟及诱导细胞炎症的发生,发现其呈现MHCⅡ抗原提呈作用,并能对IL-10与IL-12的分泌产生抑制作用。
BP26蛋白作为布鲁氏菌外膜蛋白质家族重要的毒力因子[12],对该蛋白质的研究集中于利用BP26蛋白或其家族成员与其他毒力因子相连进而制成多表位肽疫苗对动物进行相关免疫,从而预防布鲁氏菌病的发生[13]。但是对于BP26在刺激细胞后激活的信号通路和由此产生的活化信号的研究较少。此外,本试验通过生物信息学工具进行分析发现该蛋白质有大量亲水性抗原区域暴露,亲水性高,这与试验获得高效可溶性表达蛋白质的结果一致。通过表达获得的可溶性活性高的重组蛋白质,为蛋白质与宿主互作功能研究奠定了可靠的基础。
树突状细胞(DCs)是在抗原提呈中扮演着重要角色,目前研究认为,DCs为体内提呈功能最强大的专职抗原提呈细胞(APC)[14],是机体免疫应答的始动者 。将此次试验构建的重组蛋白BP26作为抗原刺激BM-DCs,通过流式细胞术分析BM-DCs表面标志物的变化,发现布鲁氏菌BP26蛋白能促进BM-DCs成熟分化,表明该蛋白质参与细胞炎症反应,在细菌对宿主细胞的侵入过程中发挥重要作用。
CD86是抗原呈递细胞上的表达分子,可提供T细胞活化和存活所必需的共刺激信号[15]。它是T细胞表面上2种蛋白质CD28抗原和CTLA-4的配体,其主要与CD28结合[16],该蛋白质与CD28抗原的结合是激活T细胞的共刺激信号。这些分子与CD80抗原一起提供必要的刺激,以引发T细胞抵抗抗原呈递细胞呈递的抗原[17]。本研究发现重组BP26蛋白刺激BM-DCs后这些表面分子表达不同程度升高,表明该蛋白能够活化T细胞反应,而MHCⅡ表面分子表达显著升高表明该蛋白质通过MHCⅡ途径提呈抗原。
CD80是CD86活化T淋巴细胞时的协同刺激因子,在自身免疫监控、体液免疫应答及移植反应中发挥重要作用[18]。CD80属于免疫球蛋白超家族,其受体是CD28和CD152(CTLA4)[19-20]。CD80在活化B淋巴细胞、活化T淋巴细胞、巨噬细胞、外周血单核细胞及树突状细胞中均可表达;在非活化B淋巴细胞、红细胞、粒细胞及核细胞上不表达[21-22],而多个研究结果表明,布鲁氏菌感染宿主后能够产生强烈的BP26蛋白相关表位的抗体[23-24],细胞因子检测结果证明BP26能够激发细胞炎症反应,刺激免疫细胞活化。本研究通过重组BP26蛋白抗原刺激BM-DCs后CD80表面分子升高,说明BP26在细菌侵入宿主引起机体产生免疫效应的过程中发挥着重要的作用。
猜你喜欢
传染病信息(2022年6期)2023-01-12
现代畜牧科技(2021年4期)2021-07-21
现代检验医学杂志(2016年4期)2016-11-15
西安工程大学学报(2016年3期)2016-06-05
中国民族医药杂志(2016年9期)2016-05-09
中国卫生标准管理(2015年2期)2016-01-14
西南医科大学学报(2015年1期)2015-08-22
医学研究杂志(2015年6期)2015-07-01
癌变·畸变·突变(2015年3期)2015-02-27
西南军医(2015年2期)2015-01-22
