
pH 值对芽孢皮层裂解酶活性及结构的影响
2020-09-10 06:16郭洪伟马慧娇郭家俊曾朝玮
食品与生物技术学报 2020年6期
郭洪伟, 马慧娇, 郭家俊, 曾朝玮, 章 中
(宁夏大学 农学院,宁夏 银川750021)
芽孢是细菌营养体在缺乏营养的条件下形成的休眠态,对辐照、超高压、热、超声波、微波、化学物质等各类杀菌处理有极端抗性[1-2],是导致食品腐败变质的根本原因。 芽孢萌发后失去其极端抗性,有许多因素能诱导芽孢的萌发,如营养素、超高压、阳离子表面活性剂、机械擦伤、外源DPA(2,6-吡啶二羧酸)、外源裂解酶等[3]。 芽孢中最常见的皮层裂解酶是CwlJ 和SleB, 这2 种酶位于芽孢内膜和外膜上,它们对芽孢的萌发具有专一性。 芽孢被激活后,这2 种酶将芽孢皮层水解然后导致芽孢核心的完全水化。无论是CwlJ 或SleB 都可使皮层降解,缺乏这2 种酶其皮层便无法降解和无法完成萌发。Brown W C[4]等报道pH<7 的N-乙酰胞壁酸会抑制芽孢孢子的萌发,该抑制剂还抑制了2 种萌发相关酶的活性。Blocher J C[5]等报道在pH 值为6.5 时,添加山梨酸完全抑制了肉毒梭菌孢子的萌发, 在pH值为5.5 时, 未添加山梨酸的菌株孢子的萌发并没有受到抑制作用。 Sheng S 等[6]报道枯草芽孢杆菌孢子CotA 的最大活性保持在pH 值为4~5 之间,可以用于固定孢子的惰性基质中, 在制备生物催化剂中,产率可以大大提高。 Porebska I 等[7]报道芽孢杆菌孢子在酸性果汁中的萌发受到了抑制。 但是萌发程度取决于菌种和时间、 孵化和使用的营养化合物,并不全取决于pH。 由此可知,酸性条件会对菌体的生长和活性有影响,对细菌芽孢的萌发有一定的抑制作用,但其抑制机理是什么,是不是因为酸抑制了芽孢皮层裂解酶的活性,这尚不清楚,有待于深入研究。 目前国内外研究芽孢皮层裂解酶的相关报道极少, 且未见关于pH 处理对芽孢皮层裂解酶活性和结构影响的相关报道,故推断存在2 种可能性:其一是酸性pH 值对皮层裂解酶有抑制作用,这种抑制作用导致细菌芽孢不萌发; 其二是酸性pH 值对皮层裂解酶无抑制作用,酸抑制芽孢的萌发另有原因。
目前,傅立叶变换红外吸收光谱仪(FT-IR)有高灵敏度、高分辨率、高效扫描及高度计算机化等长处, 是研究蛋白质分子内氢键的一种有效方法,常用来估测多肽和蛋白质二级结构[8-10]。荧光分光光度计能够扫描液相荧光标记物发射出的荧光光谱,近年来经常被用来分析蛋白质的三级结构。 酶有最适的pH 值范围,并且对酸碱度的调节极其敏感,pH值能导致盐桥的变化,并加强其三级结构,故利用这些仪器研究不同pH 处理后皮层裂解酶(CwlJ)的结构变化非常有必要。
为研究酸性pH 值对芽孢萌发的抑制机理,本文作者从研究pH 处理对皮层裂解酶(CwlJ)的活性的影响规律着手,结合酶结构变化的研究,分析皮层裂解酶 (CwlJ) 的活性和结构是否受pH 值的影响,进而揭示酸性pH 值对芽孢萌发的抑制机理,这对研究芽孢的萌发有重要意义。
1 材料与方法
1.1 材料与试剂
皮层裂解酶CwlJ(纯度>90%):南京钟鼎生物技术有限公司产品;脱芽孢衣溶液,作者所在实验室制备;Tris-HCl 缓冲液: 北京索莱宝科技有限公司产品;溴化钾(色谱纯):美国Sigma-Aldrich 公司产品。
1.2 仪器与设备
Spectrum Two 型红外光谱仪:美国PerkinElmer公司产品;Scientz-1LS 真空冷冻干燥离心浓缩仪:宁波新芝生物科技股份有限公司产品;DHG-9245A电热鼓风干燥箱: 上海一恒科学仪器有限公司产品;XMTD-6000 型电子恒温不锈钢水浴锅:上海宜昌仪器纱筛厂制造;UV-2450 型紫外可见分光光度计: 日本岛津公司产品;970CRT 荧光分光光度计:上海仪电分析仪器有限公司产品。
1.3 方法
1.3.1 CwlJ 冻干粉的前处理将CwlJ 冻干粉混入摩尔浓度为0.02 mol/L 的Tris-HCl 缓冲液中, 配成质量浓度为0.1 mg/mL 的酶溶液,为pH 的处理,测酶活和酶的二三级结构提供原材料。
1.3.2 pH 处理CwlJ 酶溶液用稀HCl 和稀H2SO4调节其溶液体系的pH 值,将CwlJ 溶液分别处理成pH 3、5、7、9、11 的酶溶液体系。
1.3.3 酶活性的测定CwlJ 的酶活计算方法可用Makino[11]的方法。在32 ℃下,通过测出1 cm 光路内脱芽孢衣的芽孢悬浮液的OD600值的减少量来分析酶活。 反应终产物体积为5 mL,包含1 mL 脱芽孢衣芽孢混合液和4 mL 酶液, 酶液包含Tris-HCl 缓冲液和皮层裂解酶CwlJ,酶活性单位(U/L)定义为:每分钟OD600值降低0.001 所需的酶量。 每10 min测一次OD600值, 根据80 min 内OD600值变化量计算酶活。
计算公式为:

式中:X 为酶活(U/L);t 为反应时间(min);V 为参与反应的酶量(mL)。
1.3.4 傅立叶红外光谱分析采用KBr 压片法制样[12]:溴化钾达到色谱纯纯度,溴化钾取200 mg,样品取2 mg,在玛瑙研钵中研磨,在170 ℃烘箱中烘24 h 以上,干燥密封保存。 用无水乙醇酒精棉球对傅立叶红外变换光谱仪以及研钵等器皿进行消毒。使用波长为400~4000 nm 的傅立叶红外变换光谱仪对CwlJ 进行全波段扫描,扫描次数为32次,分辨率为4 cm-1,每个样品测量重复3次,取平均值。
1.3.5 FT-IR 谱图处理选用Omnic 8.0 软件处理, 采集酰胺I 带的波段, 进行FT-IR 去卷积,标峰,做基础红外光谱图分析。 利用Peakfit 4.12 软件对谱峰进行标准化,平滑,去基线,求二阶导数,寻找峰, 在二阶导数原谱和酰胺I 带红外原谱的基础上进行Gauss Amp 连续拟合,进行多次拟合使残差值最小化(R2≥0.999),直至R2值不变为止[13]。 得到子峰数目为9~13 之间, 确定各个子峰的峰面积与CwlJ 的二级结构之间的比例关系,根据峰面积计算各种条件下峰的二级结构归属[14]。
1.3.6 荧光光谱分析配制质量浓度为0.1 mg/mL的CwlJ 溶液,扫描其荧光值。 设定荧光分光光度计条件: 扫描范围为200~500 nm, 扫描速度为200 nm/min,激发狭缝和发射狭缝宽均为10 nm,响应时间为0.1 s,记录数据波长间隔为1 nm。
1.3.7 数据统计分析所有实验最少重复3次,再用统计软件Origin 9.0 作图和分析数据。
2 结果与分析
2.1 不同pH 处理对CwlJ 的活性的影响
在最适反应温度32 ℃的条件下, 分别测定不同pH 值(3、5、7、9、11)的体系中CwlJ 的活力,确定最适反应pH 值[15]。 CwlJ 会造成脱芽孢衣芽孢的萌发,芽孢萌发时,其核心水化,造成折光性发生变化,表现为脱芽孢衣溶液的OD600值明显下降,下降水平越大,芽孢的萌发率就越高,皮层裂解酶活性也就越高[16-17]。
CwlJ 对pH 值的变化极其敏感见图1。 当对照是无菌水和脱芽孢衣溶液的混合液时,随着时间的变化,对照组的OD600值基本不变;pH 值为3 和pH值为5 的OD600值下降最明显,说明CwlJ 的活性保持较好,pH 值为5 是活性最高点;pH 值为7,pH 值为9 和pH 值为11 的OD600值的变化基本相同,都有显著下降,但下降幅度较低,说明当溶液中酸碱度开始趋近于中性和碱性时,酶活已经开始受到一定的抑制作用,随着处理时间的增长,酶活的降低更显著。 Miyata 等[18]研究发现:当pH 值在7.2~8.4时,皮层裂解酶活力最佳,当pH 值为4.2 时,皮层裂解酶基本失活,这一研究报道也是关于皮层裂解酶的,但实验结果与本文有一定差异,可能因为菌种不同,本实验是从枯草芽孢杆菌中分离纯化得出的皮层裂解酶,而皮层裂解酶中有CwlJ 和SleB,皮层裂解酶的相对分子质量,分子结构均不同,对pH变化的耐受程度也不同, 而本实验研究的是CwlJ,所以推测皮层裂解酶中的SleB 可能会在酸性条件下失活,而CwlJ 的活性在pH 值为3~5 左右最高,非常适宜在弱酸性条件下水解芽孢皮层而导致芽孢萌发。 可以推测,在酸性pH 条件下芽孢不萌发的原因不全是因为皮层裂解酶活受到影响[19],有可能和营养素受体、DPA 释放等原因有关, 原因有待进一步研究。
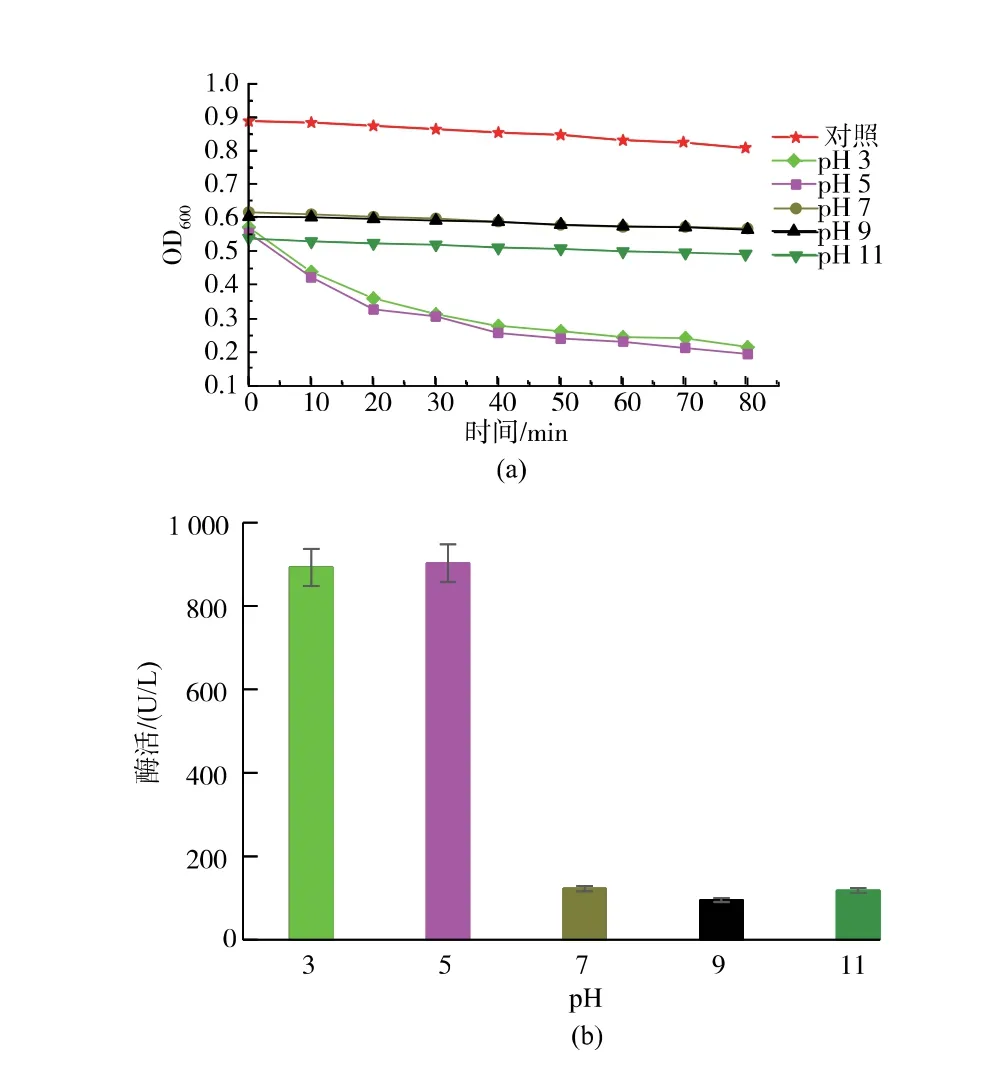
图1 不同pH 处理条件下皮层裂解酶CwlJ 活力的变化趋势Fig. 1 Change trend of CwlJ activity of skin lyase under different pH treatment conditions
2.2 不同pH 处理下CwlJ 的FT-IR 谱图的变化
利用傅立叶红外光谱仪对不同pH 处理下的CwlJ 进行全波段扫描,得到如图2 的FT-IR 谱图以及各谱带特性(表1),从图2 中能够看到全波段的FT-IR 谱图的结构变化。 如图3 可看到不同pH 处理条件下的CwlJ 的谱型基本相同, 但在3600~3300 cm-1的宽吸收峰强度不断加强,表明不同pH处理能引起CwlJ 的分子内和分子间的氢键增强。最常用的分析蛋白质的二级结构即是在1700~1600 cm-1, 酰胺I 带的震动频率主要取决于C=O之间氢键的性质,随着pH 的增大,羰基(C=O)的伸缩震动越来越微弱, 故可观察1700~1600 cm-1的波长范围的变化来研究CwlJ 的二级结构。

图2 不同pH 处理条件下皮层裂解酶CwlJ 的FT-IR 谱图Fig. 2 FT -IR spectra of cortical lyase CwlJ under different pH treatment conditions

表1 FT-IR 谱图的各谱带特性[20-21]Table 1 Characteristics of FT-IR spectra
随着pH 的增大,从峰形变化分析,峰形由大的波动逐渐变为小的波动, 见图3。 从峰位变化分析,pH 值为3 的CwlJ 的酰胺I 带为1629.5 cm-1, 随着pH 值的逐渐增大, 在对其进行酸碱度的调节过程中,酸碱度由pH 值为3 到pH 值为5 时,CwlJ 的酰胺I 带向低波数方向进行不断的平移, 红移约0.5 cm-1,即由1629.5 cm-1移至1629.0 cm-1;当pH 值为7 时,酰胺I 带的波数保持不变趋于稳定状态。随着pH 调节为碱性状态的过程中,从pH 值为7 调节到pH 值为9 时,CwlJ 的酰胺I 带向高波数方向不断的平移, 红移约35 cm-1, 即由1629.0 cm-1移至1664.0 cm-1,红移波数值比较大;当pH 值为9 调节到pH 值为11 时, 酰胺I 带向高波数不断的平移, 红移约0.5 cm-1。 这表明CwlJ 的结构会随着pH 的改变而改变,具有酸碱不稳定性,这是因为蛋白质中氨基酸肽链与外围介质构成氢键,当氢键作用较强时,C=O 的电子云密度下降, 吸收带向低波数红移; 当氢键作用较弱时,C=O 的电子云密度加强,吸收带向高波数红移[22-23]。 说明pH 处理会直接影响CwlJ 的原空间结构, 在调节酸碱度的过程中CwlJ 的分子内氢键会受到影响, 对CwlJ 的活性也会产生抑制作用。
2.3 不同pH 处理下CwlJ 酰胺I 带的拟合
酰胺I 带是评价与分析蛋白质二级结构的重要分析手段。研究表明[24],FT-IR 中酰胺Ⅰ带的峰形受蛋白质主链结构即二级结构的影响,酰胺Ⅱ带和酰胺Ⅲ带与二级结构只存在间接的影响。 在蛋白质的FT-IR 图谱中, 蛋白质的α-螺旋、β-折叠、β-转角和无规则卷曲等二级结构对应的振动吸收峰互相折叠形成一宽峰,即酰胺Ⅰ带[25]。在对酶进行傅立叶红外光谱分析其二级结构时,酶的二级结构全部表现在酰胺I 带图谱中, 酰胺I 带的二阶导数表明其各个波数所对应的不同的积分面积[26],在蛋白质的二级结构分析中, 一般采用曲线拟合的方法,对CwlJ 酰胺I 带进行基线修正,高斯去卷积,求二阶导数,曲线拟合,求出峰面积,确定分峰数量是否正确,依据峰面积估算各二级结构的相对含量[27]。其中酰胺Ⅰ带各峰位为:1615~1637 cm-1和1682~1700 cm-1为β-折叠特征峰;1646~1664 cm-1为α-螺旋特征峰;1637~1645 cm-1为无规卷曲特征峰;1664~1681 cm-1为β-转角特征峰[28]。

图3 不同pH 处理条件下皮层裂解酶CwlJ 的酰胺I 带红外原谱Fig. 3 IR spectra of the amide I band of CwlJ in the skin under different treatment conditions
图4 为pH 值为3、5、7、9、11 的CwlJ 的酰胺I带经去卷积和二阶导数处理后获得的叠加峰分解为各个子峰的高斯曲线拟合峰图。 由表2 和表3 的峰面积比例求出其结构归属。CwlJ 的二级结构主要组成为α-螺旋,其次是β-折叠、β-转角、无规则卷曲; 大小为α-螺旋>β-折叠>β-转角>无规则卷曲。经过pH 处理后的CwlJ 的二级结构发生如下变化:1) 随着pH 的增大,α-螺旋先增大后减小,pH 值从3 到9,α-螺旋逐渐增大;pH 值从9 到11,α-螺旋不断的减小。α-螺旋是蛋白质的二级结构中最稳定的,随着pH 的增大,蛋白质的二级结构发生重新排列[29],蛋白质的结构稳定性下降,直接导致CwlJ 的α-螺旋中的C=O 双键电子云密度不稳定, 氢键直接断裂并与其他极性基团发生氢键键合,发生解螺旋,影响皮层裂解酶CwlJ 活性中心的构象,活性变低;2)随着pH 的增大,无规则卷曲状态先增大后减小,pH 值从3 到7,无规则卷曲状态由15.13%增加到17.18%,pH 值从7 到9, 无规则卷曲状态由17.18%降低到15.46%, 表明随着pH 的增加,CwlJ的二级结构先从有序状态逐渐向无规则状态转化,当pH 值达到7 以上时,CwlJ 的二级结构即从无规则状态逐渐向有序状态转化[30]。 由此可证明pH 处理可能会导致CwlJ 发生水解, 空间结构呈无序状态,造成酶活降低;3)随着pH 的增大,β-转角先减小后增大,pH 值从3 到5,β-转角由22.43%降低到19.37%,当pH 值从5 到11,β-转角由19.37%增加到21.38%,β-转角本身属于无序结构, 经处理后,β-转角结构很有可能会向无规则卷曲状态转化[31-32]。 这种转化趋势与高pH 处理下CwlJ 的变性和逐渐形成分散体的形态有关,外界对芽孢的一些处理条件下,芽孢的不萌发和保持休眠态的特性可能也与CwlJ 的变性以及CwlJ 的内部结构分散有紧密的联系;4)随着pH 的增大,β-折叠先增大后减小,pH值从3 到7,β-折叠相对含量不断增加,β-折叠由26.29%增加到26.31%,pH 值从7 到11,β-折叠由26.31%增加到26.76%, 由于β-折叠比较常见于蛋白质的内部结构区域,pH 增加使得蛋白质发生解聚现象是β-折叠先增加的原因, 随着体系完全成为碱性时,蛋白质大分子因为碱性而聚集的现象导致β-折叠结构减少, 由此可推断β-折叠结构在碱性聚集体形成的过程中起到重要作用,碱性聚集体的形成使CwlJ 的活性中心被包埋,进而导致活性降低。
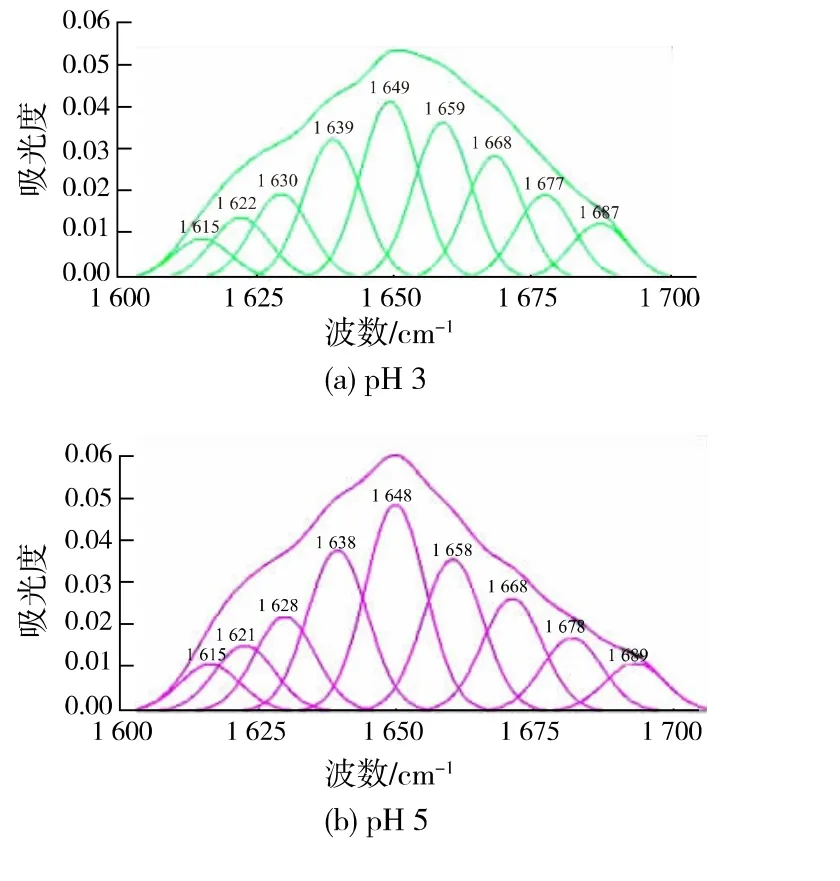

图4 不同pH 处理条件下皮层裂解酶CwlJ 酰胺I 带高斯曲线拟合图Fig. 4 Gauss curve fitting of I band of CwlJ cleavage enzyme under different pH treatment conditions

表2 不同pH 处理条件下皮层裂解酶CwlJ 酰胺I 带的不同波数的二级结构相对含量Table 2 Two level structure of different bands of CwlJ amide I band of the skin lyase under different pH treatment conditions
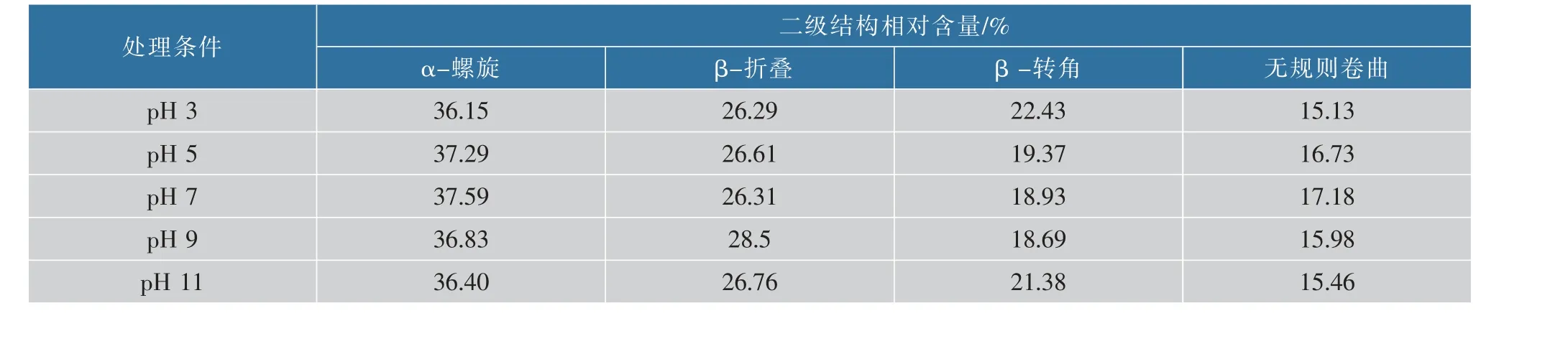
表3 不同pH 处理条件下皮层裂解酶CwlJ 酰胺I 带二级结构相对含量Table 3 Secondary structure of I band of CwlJ amide I under different treatment conditions
2.4 pH 处理对CwlJ 的荧光光谱分析
由图5 可知,样品在波长300~400 nm 范围内,各个样品的峰位,峰形都基本没有发生变化,λmax没有发生红移,说明pH 处理会导致CwlJ 的Trp 残基暴露于亲水环境中, 所以推测酶并没有发生变性,相对来讲,λmax也没有发生蓝移,说明Trp 残基自身的微环境并没有发生改变,荧光基团在亲水环境中以相对稳态的形式存在。 有出现特征峰,所有出现特征峰的最大激发波长均为348 nm,但是出现最大激发波长的峰强度显著不同。

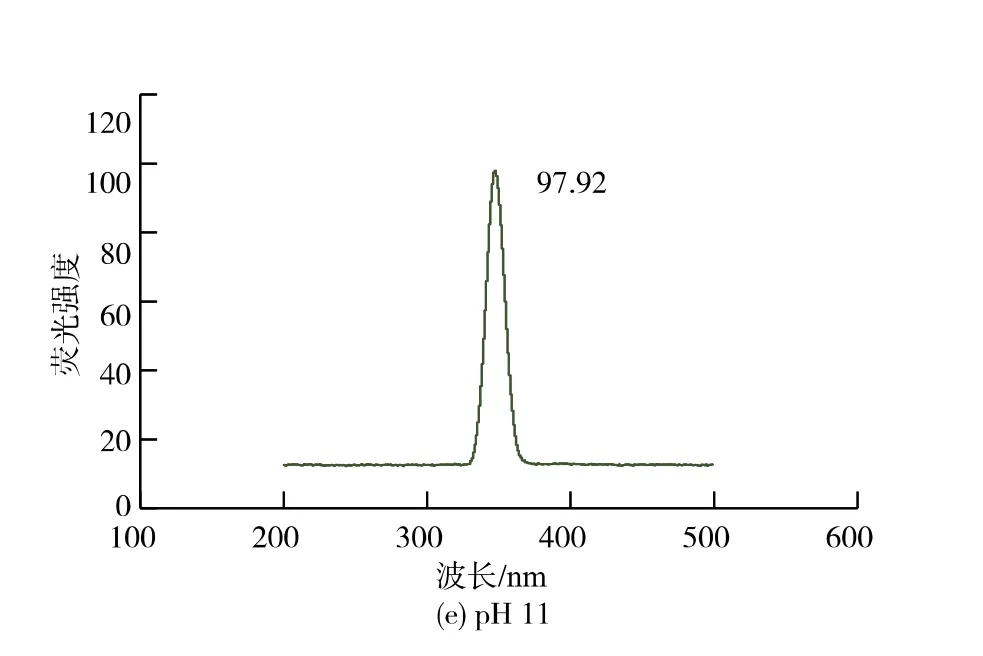
图5 不同pH 处理条件下皮层裂解酶CwlJ 的荧光光谱Fig. 5 Fluorescence spectra of cortical lyase CwlJ under different pH treatment conditions
结合CwlJ 的活性实验得出,pH 值为5 的荧光强度最强,则说明在pH 5 的条件下,CwlJ 的内部蛋白质完全展开,空间结构更加松散扩张,环境相对处于稳定状态,CwlJ 的活性较高, 其活性在这个处理条件下被完全激活, 活性中心完全暴露, 引起CwlJ 的激活效应;pH 值为11 的条件下,CwlJ 的荧光强度较强,内部结构基本完全展开,空间结构比较疏松, 其活性减少;pH 值为7 的条件下,CwlJ 荧光强度不高,蛋白质展开了一部分,空间结构比较紧密,活性减少的较强;在pH 值为3 和pH 值为9的条件下,CwlJ 的荧光强度非常低, 有可能是荧光基团发生过度的暴露,引起荧光猝灭,导致空间结构极其密集[33],在pH 值为9 的条件下活性最低,但是在pH 值为3 的条件下CwlJ 的活性比较高,推测这有可能与Trp 残基暴露于亲水环境中, 肽链过度伸展有关。
3 结 语
实验表明不同pH 处理会改变芽孢皮层裂解酶(CwlJ)的活性、二级结构和三级结构,CwlJ 的活性与结构呈正相关关系,结构越稳定,活性越强。 当pH 值为3 和pH 值为5 时,CwlJ 的活性高, 当pH值为9 时,CwlJ 活性最低, 由此可推测酸对芽孢萌发的抑制作用并不是因为其抑制了CwlJ 的活性,这证明酸性pH 值抑制芽孢萌发的原因与其对皮层裂解酶(CwlJ)活性的作用无关。 本实验推进了对细菌芽孢萌发机理的研究, 为芽孢深层机理的进一步的探究夯实了基础。
猜你喜欢
中华实验眼科杂志(2022年6期)2022-11-15
分子催化(2022年1期)2022-11-02
医学概论(2022年4期)2022-04-24
湖南饲料(2021年4期)2021-10-13
磷肥与复肥(2021年5期)2021-06-19
当代水产(2021年2期)2021-03-29
当代水产(2020年3期)2020-06-15
深圳大学学报(理工版)(2020年1期)2020-01-06
首都体育学院学报(2019年5期)2019-10-18
湖南饲料(2019年4期)2019-10-17
