
天山野苹果林立地条件和林分结构对苹果小吉丁种群数量的影响
2020-09-10 02:16:22马志龙卡德艳卡德尔张元明阿地力沙塔尔
新疆农业科学 2020年7期
马志龙,彭 彬, 卡德艳·卡德尔,张元明,阿地力·沙塔尔
(1.新疆农业大学林学与园艺学院,乌鲁木齐 830052;2.中国科学院新疆生态与地理研究所,乌鲁木齐 830011)
0 引 言
【研究意义】苹果小吉丁(AgrilusmaliMatsumura)又名串皮虫、金蛀甲,属于鞘翅目(Coleoptera)吉丁虫科(Buprestidea),1年发生1代,主要以幼虫钻蛀野苹果[Malussieversii(Led.)Roem]枝干的韧皮部和木质部进行危害,导致营养和水分输送被切断,常造成枝条干枯或整树枯死[1],是新疆巩留县乃至伊犁地区野果林的重要蛀干性害虫。自1993年在新疆发现以来,已经在野果林中大面积扩散蔓延[2],巩留县野果林苹果小吉丁发生面积甚至已经与野果林分布面积相当[3]。伊犁天山野果林是我国唯一的野果林资源分布区,也是我国唯一的世界野苹果基因库组成部分[4]。研究影响苹果小吉丁种群数量的重要环境因子,为有效控制苹果小吉丁的危害及传播有重要意义。【前人研究进展】影响苹果小吉丁发生的影响因素如寄主植物、天敌寄生等已有相关报道[5-6]。由于昆虫本身的生物学和生态学特性,其发生和分布在一定程度上受到环境因子的影响。影响昆虫发生的环境因子从类型上可以分为林分结构因子(包括林分密度、林分郁闭度、树种组成等)和立地条件因子(包括坡度、坡位、海拔和坡向等)[7]。与靠近山区的野苹果树受害情况相比,靠近河谷的野苹果树虫口密度更大、受害更严重[4]。苹果小吉丁成虫具有喜热、喜光习性,活跃程度随着温度的升高而增强,并且喜欢将卵产于树冠向光处,其在林间的分布明显受到温度、光照等条件变化的影响[8-9]。多样性-稳定性假说和天敌假说中提出,随着林间植物种类的增加,昆虫的种类随之增多,各类昆虫的发生期互相弥补及重叠,容易形成稳定的食物链,提高天敌控制能力[10-11]。与多个树种组成的混交林分相比,树种较为单一的纯林中植物性昆虫更容易爆发[13-14]。【本研究切入点】有关野果林林分结构和立地因子对苹果小吉丁产生影响的相关研究依然较少。研究伊犁地区野苹果林苹果小吉丁种群数量与林分结构和立地条件的关系分析各调查因子对苹果小吉丁种群数量的影响作用及影响程度。【拟解决的关键问题】调查对比分析不同立地条件、林分结构条件下苹果小吉丁种群发生规律,研究具有联合抗性作用的立地条件、林分结构中的关键因子对苹果小吉丁种群数量变动的影响。
1 材料与方法
1.1 材 料
研究区位于新疆巩留县库尔德宁镇野苹果林和新源县哈拉不拉乡萨哈野苹果林,地理坐标为E82°49′40.15″~82°51′58.41″,N43°14′11.56″~43°15′35.36″,主要垂直分布于海拔1 200~1 500 m的山沟中、山坡上及低山丘陵带,分布状态为不连续的条块状。样地林分气候为温带大陆性相对较为湿润的气候,立地土壤为山地黑棕色野苹果林土。野苹果林系统中生物资源丰富,除了野苹果以外,林间还自然分布核桃JuglansregiaLinn.、野杏ArmeniacavulgaisLam.、野生欧洲李PrunusdomesticaLinn.、准噶尔山楂CrataegussongoricaKoch.、蔷薇RosaacicularisLindl.等乔、灌木。在巩留县库尔德宁镇的阿勒玛塞、金吉克塞以及哈拉不拉乡萨哈等野果林分布区各选取3块调查样地;在库尔德宁镇阔克塔勒村和72团8连各选取2块调查样地;在库尔德宁镇角勒塞选取4块调查样地。每块调查样地面积50 m×50 m。表1

表1 样地林分指标信息Table 1 Stand index information of sample land
SYNTEK电子数显坡度仪(400MM标准型)、镊子、手锯、修枝剪、解剖镜、解剖刀、手持GPS(EXPLORIST 500)、放大镜、标本夹、自喷漆、记号笔、记录本、皮尺、游标卡尺、标签、罗盘仪(DQY-1型)和测高仪(CGQ-1型)等。
1.2 方 法
1.2 .1 林分结构因子
分别于2017年6至9月和2018年6至9月进行林间实地调查,调查样地的林分郁闭度、林分密度、野苹果比例和树种丰富度等林分结构因子,坡度、坡位、坡向和海拔等立地条件因子。在每块样地内采用“Z”字型取样法随机选取10株野苹果树,分别从东、西、南、北四个方位,树冠2.5~3 m各随机截取一段枝条,用卷尺量取枝条长度,用解剖刀解剖枝条韧皮部和木质部记录并计算苹果小吉丁平均虫口数量。
平均虫口数量=总虫口数量(头)/总枝条长度[15]。
林分郁闭度采用对角线法进行测量,每1 m设1个样点,统计被树冠投影的样点数量占总样点数量的比值。对样地内所有胸径≥5 cm的树木种类和数量进行调查,使用手持测高仪(CGQ-1型)测定树高,同时计算野苹果比例和树种丰富度。树种丰富度(MA)公式为:
式中,S为样地中树种总数,N为各树种的数量之和[7]。
林分密度采用优势高-营养面积比(Z)作为指标进行调查。林分密度公式为:
式中,Z为优势高-营养面积比;H0为优势木平均高;N为株数密度(即每公顷株数)[16]。
1.2.2 立地条件因子调查
样地位置通过手持GPS(EXPLORIST500型)进行定位,同时对样地海拔高度进行测量。通过使用SYNTEK电子数显坡度仪(400 MM标准型)和罗盘仪(DQY-1型)测量样地的坡度和坡向。坡位按林分在山坡上的分布位置分为上、中、下3个坡位级[17]。
1.2.3 数据量化
设因变量Y为苹果小吉丁的虫口数量,自变量X项中设坡度为X1、海拔为X4、林分郁闭度为X5、林分密度为X6、树种丰富度为X8。其他调查因子先通过数据量化转换再在统计软件上进行输入。转换方式如下:
坡位X2:下坡、中坡、上坡分别用1、2、3表示;
坡向X3:阴坡、半阴半阳坡、阳坡分别用1、2、3表示;
野苹果比例X7:采用野苹果所占林分树种比例数表示,野苹果纯林用1表示。
1.3 数据处理
通过对苹果小吉丁虫口数量和各调查因子的关系进行偏相关分析,根据各调查因子的偏相关系数大小确定其对苹果小吉丁虫口数量的影响程度。
通过运用SPSS17.0统计软件将8个调查因子与苹果小吉丁的虫口数量进行偏相关分析,计算各调查因子的显著性及影响效应,筛选出关键影响因子。建立关键影响因子与苹果小吉丁的平均虫口数量之间的线性回归模型,利用模型对样地平均虫口数量进行预测,计算预测值与实测值的差异度,同时对两者进行独立样本检验,对模型的预测可靠性进行检验。
2 结果与分析
2.1 立地条件因子对苹果小吉丁种群数量影响
研究表明,立地条件因子中,缓坡(6°≤坡度≤15°)林分平均虫口数量显著高于陡坡(26°≤坡度≤35°),为1.24头/1 m样枝,但与斜坡林分平均虫口数量差异不显著,且斜坡与陡坡之间差异也不显著(P>0.05)。随着海拔的增加,苹果小吉丁平均虫口数量逐渐降低,1 200~1 249 m、1 250~1 299 m、1 300~1 349 m和1 350~1 399 m海拔范围之间平均虫口数量差异不显著(P>0.05),但与>1 400 m林分之间差异显著(P<0.05);海拔>1 400 m的林分内平均虫口数量仅为0.49头/m样枝。不同坡位间,上坡平均虫口数量最少,中坡次之,下坡最高,平均虫口数量依次为0.60头/m样枝、0.81头/m样枝和1.08头/m样枝,但3个坡位之间差异性不显著(P>0.05)。从不同坡向来看,阳坡林分平均虫口数量最高,达到1.5头/1 m样枝,显著高于阴坡和半阴坡,而后两者之间无显著差异(P>0.05)。图1

图1 苹果小吉丁发生与立地因素的关系Fig. 1 The relationship between the occurrence of Agrilus mali Matsumura and site factors
2.2 林分结构因子对苹果小吉丁种群数量影响
研究表明,各林分结构因子中,随着林分郁闭度的增加,平均虫口数量逐渐降低,林分郁闭度为0~0.29的林分平均虫口数量最高,达到1.44头/m样枝,显著高于郁闭度为0.30~0.49和0.50~0.79的林分(P<0.05),而后两者之间差异性不显著(P>0.05)。树种丰富度>0.60的林分平均虫口数量仅为0.49头/m样枝,显著低于树种丰富度为<0.30的1.5头/m样枝和0.31~0.59的1.13头/m样枝。野苹果纯林(即野苹果比例为1)平均虫口数量最高,达到1.5头/m样枝,显著高于野苹果比例≤0.50的林分,但与0.51~0.79和0.80~0.99三者之间无显著差异(P>0.05)。林分密度<0.11的稀疏林分平均虫口数量最高,为1.5头/m样枝,随着林分密度的增加,平均虫口数量呈下降趋势。图2

图2 苹果小吉丁虫口数量与林分因素的关系Fig. 2 The relationship between the population of Agrilus mali Matsumura and forest factors
2.3 偏相关系数
研究表明,海拔X1、坡度X4、和坡位X3的偏相关系数绝对值较大,分别为0.598、0.542和0.423。T检验结果表明,海拔X1和坡度X4与平均虫口数量的关系达到显著水平(PX1=0.031;PX4=0.047;<0.05);坡位X3与平均虫口数量关系未达到显著水平(PX3=0.150;>0.05)。表2

表2 各自变量偏相关系数Table 2 Partial correlation coefficients of respective variables
2.4 苹果小吉丁种群数量与关键因子
2.4.1 苹果小吉丁平均虫口数量与海拔的关系
通过将苹果小吉丁平均虫口数量(Y)与海拔(X)的回归关系进行计算,得到方程为:Y= -0.007 5x+ 10.849,经F检验表明,相关性达到显著水平(F=7.503,df=1,16,P=0.015)。海拔位于1 370 m以下的11块样地中,有7块样地的平均虫口数量在1头/m样枝以上,其中1 238 m林分平均虫口数量达到1.94头/m样枝,为所有林分内最高。海拔为1 427 m的样地平均虫口数量最低,仅为0.19头/m样枝。图3
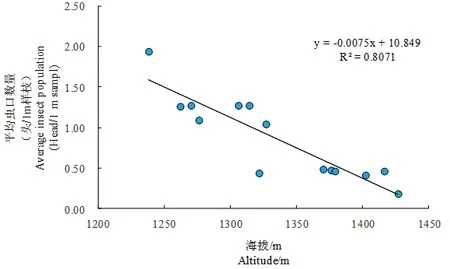
图3 苹果小吉丁虫口数量与海拔的关系Fig. 3 The relationship between the population of Agrilus mali Matsumura with altitude
2.4.2 苹果小吉丁平均虫口数量与坡度的关系
通过将苹果小吉丁平均虫口数量(Y)与坡度(X)的回归关系进行拟合,得到方程为Y= -0.049 8X+ 1.906 2,对该方程经过F检验表明相关性达到显著水平(F=12.025,df=1,16,P=0.003)。林间调查发现,随着林地坡度的增加,林分平均虫口数量逐渐降低。平缓立地(0°<坡度<15°)样地内平均虫口数量最高,达到1.24头/m样枝,显著高于陡峭立地(25°<坡度<35°)的0.39头/m样枝。图4

图4 苹果小吉丁虫口数量与坡度关系Fig.4 The relationship between the population of Agrilus mali Matsumura with grade
2.5 多元线性回归模型的建立及检验
将海拔(X1)和坡度(X4)2个关键因子与林分苹果小吉丁平均虫口数量(Y)建立回归模型为:Y=5.541-0.003X1-0.033X4。经F检验表明,该线性回归预测模型达到显著水平(F=12.021,df=2,16,P=0.001)。通过使用该模型对苹果小吉丁的虫口数量进行预测评估较为可靠。通过使用15块样地平均虫口数量的调查数据对该回归模型进行检验,将计算得出的平均虫口数量预测值与实测值进行对比,换算出差异度[18,19]。
研究表明,有53%的样地差异度小于0.2,而所有样地平均差异度为0.193,多元线性回归模型的预测值较为可靠。对苹果小吉丁平均虫口数量实测值与预测值进行独立样本T检验,两者之间无显著差异。表3
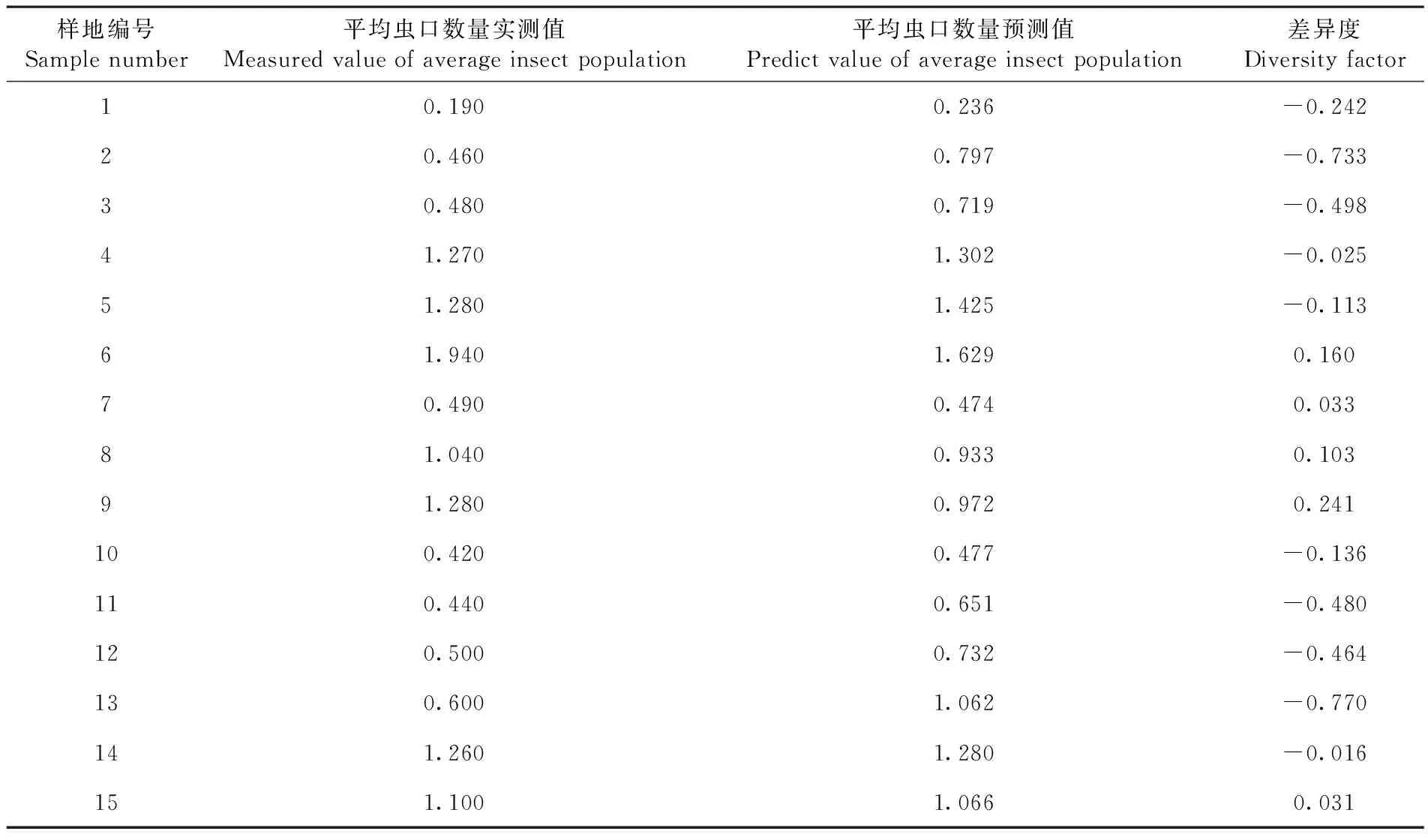
表3 多元线性回归模型检验Table 3 Multiple linear regression model test
3 讨 论
森林生态系统中,植食性昆虫与寄主林木之间的关系是主要的互作关系和核心问题[20]。就野苹果树木个体而言,在外部形态特征上,野苹果单株胸径、树高、树皮厚度和树势均对苹果小吉丁的危害程度产生显著的影响作用[8];在内部营养物质方面,野苹果植株单宁与总酚的含量越高,其抵御苹果小吉丁入侵的能力越强[21]。而植物与植食性昆虫的互作关系并非都是独立存在的,除了植食性昆虫的内禀增长能力外,往往受到生存环境的影响,使寄主-昆虫-环境各因素协同构成植物的抗虫性[5, 22]。
海拔、坡度、坡向等立地因子,可以综合反映出地表形态结构的变化,各立地因子之间相互作用、相互联系,使温度、湿度和光照等气候因子形成了梯度效应[23, 24]。海拔、坡位和坡向均主导着林分温度和光照等条件,海拔越高、林分越接近上坡,林间的温度相对越低。1 250~1 299 m海拔范围林分平均虫口数量达到1.26头/m样枝;当海拔>1 400 m时,平均虫口数量降低为0.49头/m样枝。随着坡位的上升,平均虫口数量逐渐降低;与阴坡相比,阳坡接受光照的条件明显更好,所以阳坡林分更利于苹果小吉丁种群的稳定和发展。由于立地平缓的林分接受光照的时间和林分内环境温度要大于立地陡峭的林分,所以坡度较为平缓的林分苹果小吉丁发生程度更为严重。当林分坡度达到26°(陡坡)以上时,林间苹果小吉丁虫口数量仅为0.39头/m样枝。由坡度导致的光照和温度条件依然是影响苹果小吉丁种群数量的关键因素,该研究结论与红脂大小蠹(DendroctonusvalensLeConte)与林分生境的关系结论基本一致[25]。
植食性昆虫选择寄主植物一般依靠触角来识别寄主植物释放的挥发性物质[26],国外学者通过试验证明了寄主树种栎树(QurecusL.)树叶和树皮中的挥发性气味能够引起栎双点吉丁(AgrilusbiguttatusFabricus)的强烈反应[27]。在寄主定位过程中许多昆虫的视觉也发挥着重要作用[28],所以树种丰富度越大、野苹果比例越低的林分其林木的种类和数量越多,林间气味成分更为复杂,且由于寄主树种和非寄主树种的交错分布可能会对苹果小吉丁通过嗅觉和视觉进行寄主植物定位造成一定的阻碍,影响了苹果小吉丁种群数量的发展。这与松墨天牛(MonochamusalternatusHope)发生与马尾松(PinusmassonianaLamb.)混交模式的关系基本一致[29]。郁闭程度高的林分,透光条件明显比低郁闭林分差,降低了苹果小吉丁成虫的活跃程度和产卵选择,所以林分郁闭度达到0.50以上时苹果小吉丁种群数量显著小于低郁闭林分。由于条件有限,试验仅在伊犁天山野果林中开展了相关研究,而苹果小吉丁在栽培果园中也已造成一定的危害。可以进一步研究平原栽培果园的种植密度、冠幅大小、套种植物等因素对果园苹果小吉丁数量的影响作用。
4 结 论
4.1 林分的海拔和坡位主要对林间的温度起着调控作用,海拔越高、林分越接近上坡,林间的温度相对越低。坡向和坡度主要对林间的受光条件起着主导作用,坡向越接近阳坡、坡度越平缓的林分,受光条件越好。苹果小吉丁虫在低海拔、下坡位的林分中活跃程度更强、种群数量更多。那些处于阳坡、立地较为平缓的林分,由于受光条件更好,更有利于苹果小吉丁进行产卵选择。
各立地条件因子中,海拔和坡度是苹果小吉丁种群数量的关键影响因子,偏相关系数依次达到-0.598和-0.542,均为负影响作用,且相关性均达到显著水平(P<0.05)。坡位和坡向的影响作用次之,且相关性均未达到显著水平(P>0.05)。根据各因子对苹果小吉丁种群数量的影响作用大小可依次排序为海拔>坡度>坡位>坡向。
4.2 树种丰富度和野苹果比例反映的是林间的树种组成情况,树种丰富度越大、野苹果比例越低的林分,林间非寄主植物占比越高、树木组成结构越为复杂,越容易形成稳定的群落系统从而对苹果小吉丁形成更高的抵御能力。林分郁闭度和林分密度反映着林木的郁闭程度和拥挤程度,随着林分郁闭程度和拥挤程度的增加,林间温度和透光条件随之降低,对苹果小吉丁的活跃能力起到一定的限制作用。
各林分结构因子中,林分郁闭度、林分密度、野苹果比例和树种丰富度与苹果小吉丁种群数量的偏相关系数较小、影响作用相对稍弱,且相关性均未达到显著水平(P>0.05),均为非关键因子。但根据各因子对苹果小吉丁种群数量的影响作用大小可依次排序为:林分密度>林分郁闭度>野苹果比例>树种丰富度。
4.3 对野苹果林进行人工抚育时,应该重点对海拔低于1 400 m和分布于平缓立地(坡度<15°)的林分进行苹果小吉丁虫口监测及防治。同时,通过人工补植逐渐提高林分密度和郁闭度,也可一定程度提高对苹果小吉丁的抗性。
4.4 由海拔、坡度与苹果小吉丁虫口数量建立的预测模型Y=5.541-0.003X1-0.033X4,可以较为可靠的对林间苹果小吉丁虫口数量进行初步预估,减少工作量,提高林间实地调查工作效率。
猜你喜欢
今日农业(2021年3期)2021-12-05 01:46:23
现代园艺(2021年23期)2021-12-01 07:47:44
今日农业(2021年9期)2021-07-28 07:08:22
今日农业(2021年4期)2021-06-09 06:59:44
天津农林科技(2021年2期)2021-04-16 02:54:24
林业勘查设计(2020年1期)2021-01-18 02:40:48
新农业(2020年18期)2021-01-07 02:17:08
防护林科技(2020年6期)2020-08-12 13:34:40
绿色科技(2019年6期)2019-04-12 05:38:42
绿色科技(2019年6期)2019-04-12 05:38:42
