
鞭段长度和径级对菲黄竹母鞭和新鞭主要养分特征的影响
2020-09-07 08:24陈双林郭子武
江西农业大学学报 2020年3期
谷 瑞,徐 森,陈双林*,郭子武,章 超
(1.中国林业科学研究院 亚热带林业研究所,浙江 杭州 311400;2.南京林业大学,江苏 南京 210037)
【研究意义】C、N、P 作为植物的三大主要元素,C 是植物各种生理生化过程的底物和能量来源,是植物体的结构性物质[1],N和P是蛋白质和遗传物质的重要组成元素[2],是植物生长的限制性养分,植物体内C 的储备一定程度上是由N、P 可获得的量来进行调控的[3]。C、N、P 的有效性反映了植物可利用的营养水平,对植物的生长发育有显著的影响[4]。碳水化合物是植物光合作用的主要产物,其组分包括结构性碳水化合物(structural carbohydrate,SC)和非结构性碳水化合物(non-structure carbohydrate,NSC),SC 主要用于植物的形态建成,包括木质素、纤维素等;NSC 是参与植物生命代谢的重要物质,包括可溶性糖(果聚糖、蔗糖、葡萄糖、果糖等)和淀粉等[5]。NSC既是光合作用的产物,也是呼吸作用的底物,是衡量植物碳吸收(光合作用)与碳消耗(生长和呼吸)之间关系的指标[6]。竹子是典型的根状茎型木质克隆植物[7-8],通过地下根状茎(竹鞭)在水平方向的快速生长,可以形成由大量分株(竹株)通过竹鞭彼此相连而成的克隆网络(clonal network),从而可以实现相连分株之间物质和能量的传递和共享[7,9]及分株的选择性放置(觅食行为foraging behavior)等[7]。而克隆器官(根状茎)是重要的营养资源短期贮藏库,其存储的物质和不定芽可以有效地控制新生分株(竹笋)的产生、存活和生长[10],分株的生长有赖于良好的根状茎结构,因此其形态及生理特征决定了克隆植物潜在的克隆性,在竹子的营养繁殖中起着重要的作用。【前人研究进展】已有对草本克隆植物的研究表明,匍匐茎的节间长度与养分储量呈正相关,增加节间长度可能有助于克隆片段的生存和生长[11];增加匍匐茎的粗度和节间长度,可以促进克隆片段的生长,提高其存活率[12],因此匍匐茎的形态及生理特征与克隆植物的生长状况密切相关。【本研究切入点】笔者前期研究表明,鞭长和鞭径对地被竹容器苗生长有明显影响,但对地被竹地下茎养分变化的影响尚不清楚。【拟解决的关键问题】为此,本研究以菲黄竹(Sasa auricomaE.G.Camus)一年生鞭段为试材,开展了不同鞭长和鞭径组合处理下的母鞭和新鞭C、N、P 含量与化学计量比和碳水化合物及其组分含量的差异比较,分析菲黄竹鞭段容器苗地下鞭养分对鞭长和鞭径的响应规律,筛选适于菲黄竹鞭段容器育苗的鞭长和鞭径组合,为地被竹高质量鞭段容器育苗提供参考。
1 材料与方法
1.1 试验地概况
试验地位于浙江省杭州市临安区(29°56′~30°23′N,118°51′~119°72′E)太湖源观赏竹种园,属亚热带季风气候,温暖湿润,四季分明,年降水量1 250~1 600 mm,年平均气温15.4 ℃,1月平均气温3.2 ℃,7月平均气温29.9 ℃,极端最低温度-13.3 ℃,极端最高温度40.2 ℃,全年大于10 ℃的平均活动积温5 100 ℃,年平均无霜期235 d,年日照时数1 850~1 950 h,土壤为红壤,土层厚度大于60 cm,土质肥沃,结构疏松,十分适宜竹类植物生长。
1.2 试验材料与设计
菲黄竹外形秀丽、枝叶密集、耐修剪,具有观秆、观叶、观丛等多种观赏价值,且生长繁殖速度快、抗逆性强,种植后易于管理、连年不衰,地下竹鞭纵横交错,有很强的护坡和水土保持等功能,是园林绿化中广泛应用的优质地被类观赏竹种[13-15]。近年来,随着地被竹在园林绿化和护岸固土等方面的需求不断扩大,而鞭段容器育苗因育苗繁殖成本低、育苗技术简单易操作、生长繁殖速度快需时短、苗木规格和质量易于控制而越来越受到关注,成为竹类扩繁的重要方式[16-18]。
2018年3月上旬,在健康生长的菲黄竹林中,挖取生长健壮、鞭芽饱满、无病虫害、长度20 cm以上的一年生菲黄竹竹鞭,选取细鞭(D1,鞭径为2.67±0.32 mm)、粗鞭(D2,鞭径为5.20±0.46 mm)2种径级的竹鞭开展3 种鞭段长度(L1,3 cm;L2,6 cm;L3,9 cm)6 种处理的埋鞭育苗试验,分别记作D1L1(细鞭,3 cm)、D1L2(细鞭,6 cm)、D1L3(细鞭,9 cm)、D2L1(粗鞭,3 cm)、D2L2(粗鞭,6 cm)、D2L3(粗鞭,9 cm),每个处理各50 盆,共计300 盆。埋鞭育苗时每个容器内总鞭长一致,均为18 cm,即2 种鞭径3、6、9 cm 3 种处理长度的每盆容器埋鞭的鞭段数分别为6、3、2 段。育苗容器为下口径10 cm、上口径13 cm、高度13 cm 的塑料营养钵,育苗时先在营养钵中填充基质至离容器口沿5 cm 高度左右,再将鞭段平放于容器的基质上,后将基质填充至离容器口沿1 cm左右,压实,浇水后置于遮荫棚内进行养护,注意及时灌溉和人工清除杂草,保护竹笋-幼竹生长。栽培基质为泥炭、废砻糠、黄心土体积比5∶3∶2 均匀混合而成,同时基质中均匀拌入体积比为1/30缓释肥(N∶P2O5∶K2O=15∶15∶15)。
1.3 取样及养分含量测定
于2018年7月上旬,每个处理随机取竹苗15盆,5盆为1个重复,共3个重复。将不同处理的每盆容器苗去除基质洗净,用吸水纸擦干后,分离母鞭(埋鞭)与新鞭,置于烘箱内先经105 ℃杀青30 min,然后80 ℃烘至恒质量,粉碎,过40目筛,装袋储于真空干燥器中以备化学分析。竹鞭全碳含量采用重铬酸钾容量法测定,全氮含量采用凯氏定氮法测定,全磷含量采用钼锑抗比色法测定[19],可溶性糖、淀粉含量采用蒽酮比色法测定[20],纤维素、木质素含量采用酸性洗涤纤维法测定[21]。
1.4 数据分析
试验数据在Excel 2007 统计软件进行整理和图表制作,在SPSS 24.0 统计软件中采用双因素(Twoway ANOVA)方差分析检验鞭长、鞭径及其交互效应对菲黄竹地下鞭养分含量的影响;采用单因素(Oneway ANOVA)方差分析比较不同鞭长间地下鞭养分含量的差异,采用独立样本t检验(Student’s t Test)分析不同鞭径间地下鞭养分含量的差异(α=0.05)。试验数据均表示平均值±标准差。
2 结果与分析
2.1 鞭长和鞭径对菲黄竹鞭段繁育容器苗地下鞭C、N、P含量的影响
鞭长对菲黄竹母鞭的C、N、P 含量均有显著影响,但鞭径仅显著影响P 含量,对C、N 含量无显著影响,且鞭长、鞭径对母鞭C、N、P 含量的交互作用显著(表1)。随鞭长的增大,D1 鞭径母鞭C 含量显著升高,D1L1 处理显著高于D1L2、D1L3 处理,D1L3 处理显著高于D1L2 处理,N、P 含量呈降低趋势,D1L1 处理显著高于D1L2、D1L3 处理,后两者间无显著差异;D2 鞭径母鞭C 含量总体呈降低趋势,N、P 含量呈升高趋势,其中,D2L1 处理显著低于D2L2、D2L3 处理,后两者间无显著差异。随鞭径的增大,L1 鞭长母鞭C 含量显著升高,而N、P 含量显著降低,L2、L3 鞭长则相反(表2)。但鞭长、鞭径及其交互作用对菲黄竹新鞭C、N、P 含量均无显著影响(表2)。可见,菲黄竹新鞭在不同鞭长和鞭径处理下均能正常生长,鞭长和鞭径对菲黄竹母鞭C、N、P含量有较明显的影响,但对菲黄竹新鞭的影响不显著。
2.2 鞭长和鞭径对菲黄竹鞭段繁育容器苗地下鞭C∶N∶P的影响
鞭长对菲黄竹母鞭C∶N、C∶P影响显著,鞭径对菲黄竹母鞭N∶P影响显著,且鞭长、鞭径对母鞭C∶N∶P交互作用显著(表1)。随鞭长的增大,D1鞭径母鞭N∶P无显著差异,C∶N、C∶P显著升高,D1L3处理均显著高于D1L1、D1L2处理,D1L2处理均显著高于D1L1处理;D2鞭径母鞭N∶P亦无显著差异,而C∶N、C∶P均呈降低趋势,D2L1 处理均显著高于D2L2、D2L3 处理,后两者间无显著差异。随鞭径的增大,L1 鞭长母鞭C∶N、C∶P 均显著增大,而L2、L3 鞭长母鞭C∶N、C∶P 则均显著降低,不同鞭长母鞭N∶P 并无显著差异(表2)。但鞭长、鞭径及其交互作用对菲黄竹新鞭C∶N∶P 均无显著影响(表2)。表明,鞭长和鞭径对菲黄竹母鞭C∶N∶P有较明显的影响,但对菲黄竹新鞭的影响不显著,体现了菲黄竹新鞭具有较高的养分内稳性。
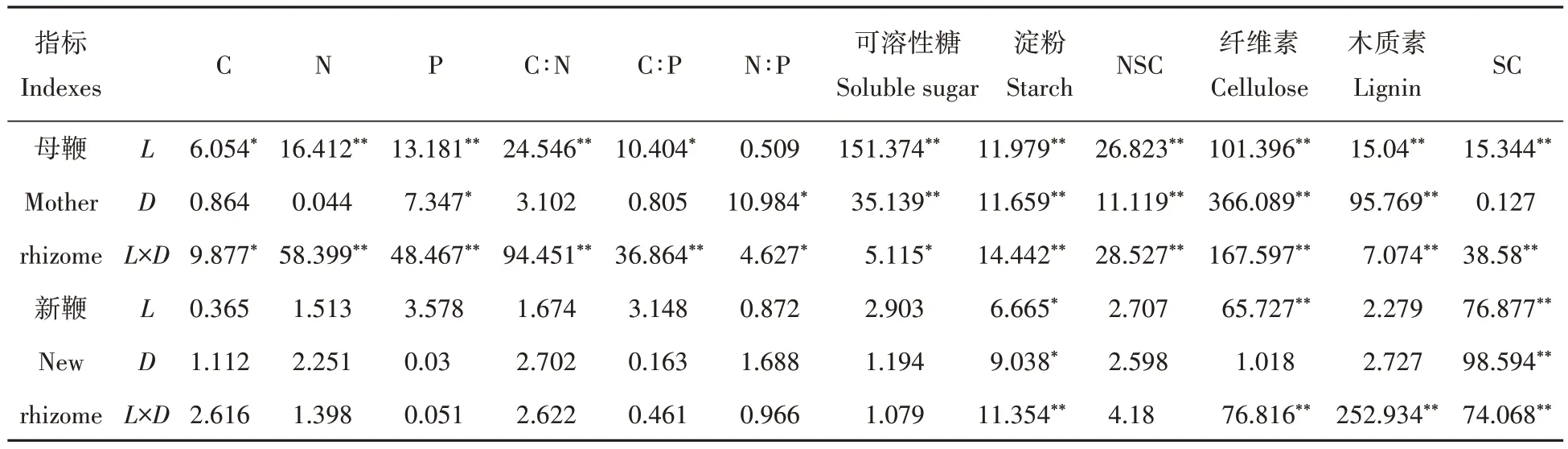
表1 鞭长和鞭径对菲黄竹鞭段繁育容器苗地下鞭养分影响的双因素方差分析Tab.1 Two-factor variance analysis of rhizome length and rhizome diameter on underground rhizome nutrients of container seedlings bred by Sasa auricoma rhizome segment
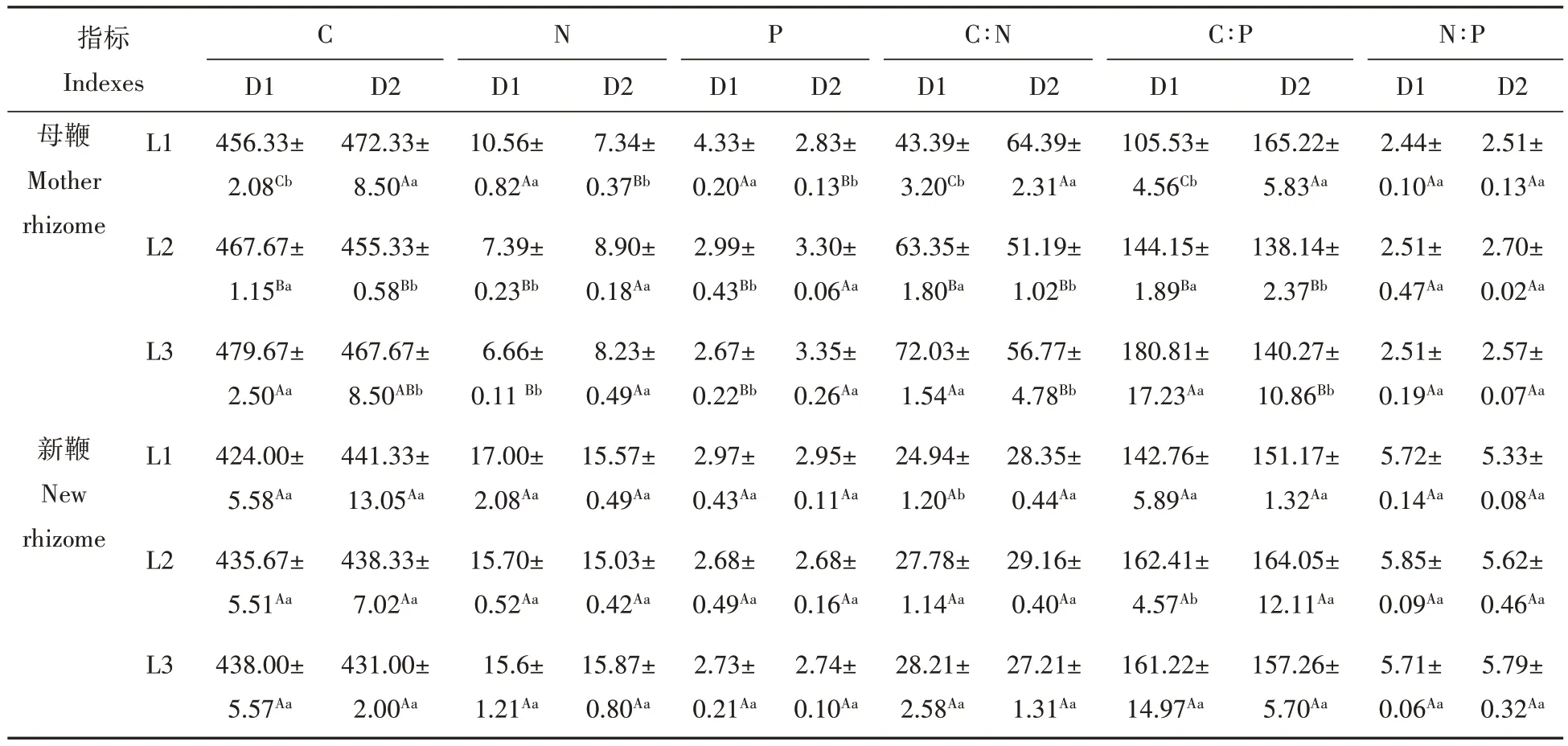
表2 鞭长和鞭径处理下菲黄竹鞭段繁育容器苗地下鞭C、N、P含量及化学计量比Tab.2 C、N、P content and stoichiometric ratio of underground rhizome of rhizome segment breeding container seedlings of Sasa auricoma under the rhizome length and rhizome diameter treatment mg/g
2.3 鞭长和鞭径对菲黄竹鞭段繁育容器苗地下鞭非结构性碳水化合物含量的影响
鞭长和鞭径对菲黄竹母鞭NSC及其组分含量均有显著影响,且存在显著的交互作用(表1)。随鞭长的增大,D1 鞭径母鞭可溶性糖含量呈“∧”变化规律,D1L2 处理显著高于D1L1、D1L3 处理,D1L3 处理显著高于D1L1 处理,而淀粉含量则呈降低趋势,D1L1 处理显著低于D1L2、D1L3 处理,且后两者间无显著差异,但NSC 含量变化不明显;D2 鞭径母鞭可溶性糖含量也呈“∧”变化规律,D1L2 处理显著高于D1L1、D1L3 处理,D1L3 处理显著高于D1L1 处理,而淀粉和NSC 含量则呈升高趋势,D2L3 处理均显著高于D2L1、D2L2 处理,后两者间无显著差异。随鞭径的增大,L1、L2 鞭长母鞭可溶性糖含量变化不明显,淀粉和NSC含量则显著降低,L3鞭长母鞭NSC及其组分含量则均随鞭长增大而显著增加(表3)。
鞭长、鞭径及其交互作用仅对菲黄竹新鞭淀粉含量有显著影响,而对可溶性糖、NSC 含量的影响均未达显著水平(表1)。随鞭长的增大,D1 鞭径新鞭可溶性糖含量变化不明显,而淀粉、NSC 含量均呈降低趋势,D1L1 处理均显著大于D1L2、D1L3 处理,后两者间无显著差异,D2 鞭径新鞭NSC 及其组分含量均无明显变化。随鞭径的增大,L1 鞭长新鞭可溶性糖含量变化不明显,而淀粉和NSC 含量则显著降低,L2、L3鞭长新鞭NSC 及其组分含量均无明显变化(表3)。可见,鞭长和鞭径对菲黄竹鞭段容器苗地下鞭NSC组分含量的影响主要体现在母鞭上,对新鞭的影响较小。
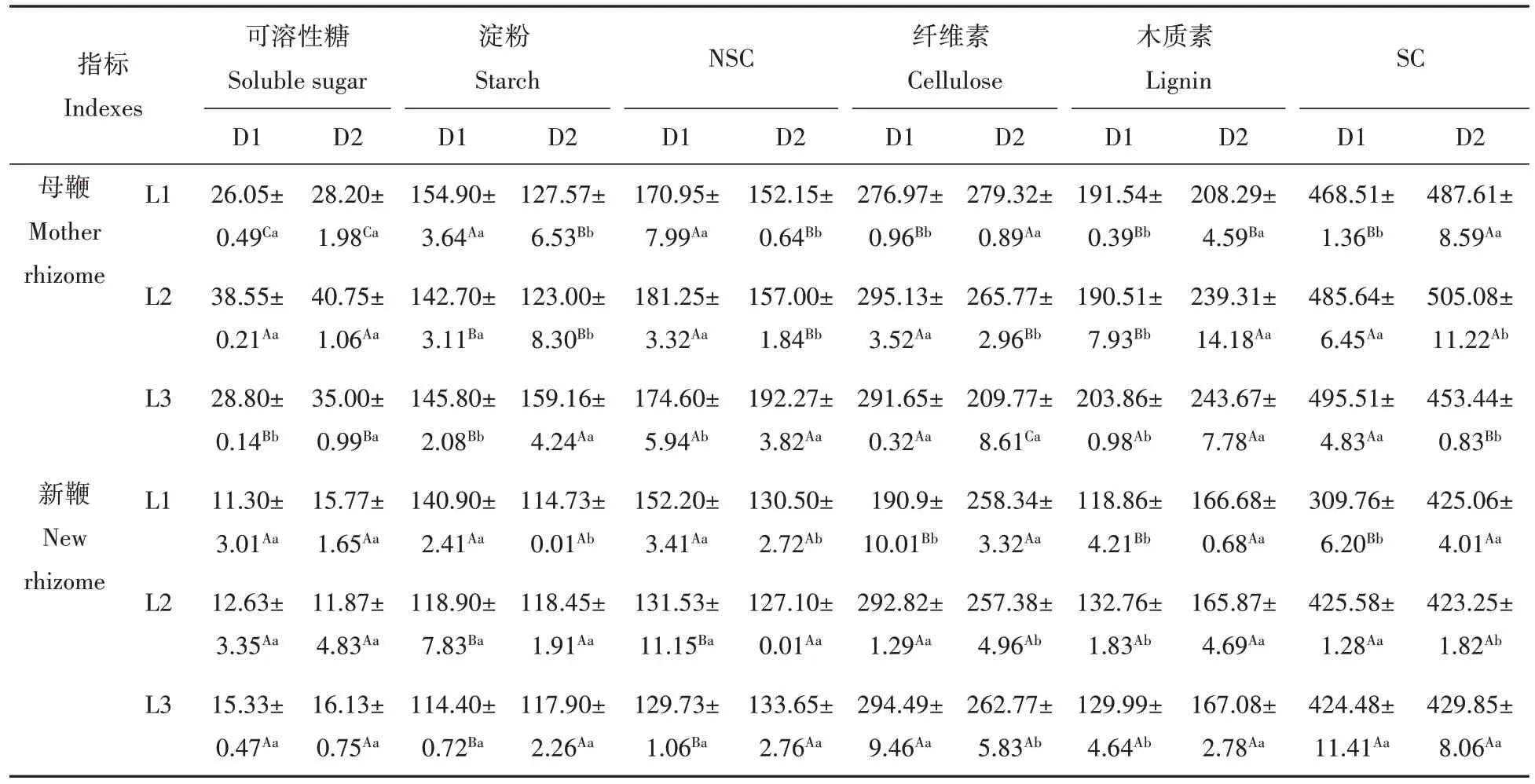
表3 鞭长和鞭径处理下菲黄竹鞭段繁育容器苗地下鞭碳水化合物含量Tab.3 The content of carbohydrates in the underground rhizome of rhizome segment breeding container seedlings of Sasa auricoma under the rhizome length and rhizome diameter treatment mg/g
2.4 鞭长和鞭径对菲黄竹鞭段繁育容器苗地下鞭结构性碳水化合物含量的影响
鞭长对菲黄竹母鞭SC 及其组分含量均有显著影响,鞭径对菲黄竹母鞭纤维素、木质素含量有显著影响,而对SC 含量影响并不显著,但鞭长和鞭径的交互作用对SC 及其组分含量均有显著影响(表1)。随鞭长的增大,D1鞭径母鞭SC及其组分含量均总体呈升高趋势,D2鞭径母鞭木质素含量总体呈升高趋势,而纤维素、SC含量总体呈降低趋势。随鞭径的增大,L1鞭长母鞭SC及其组分含量均显著升高,L2鞭长母鞭纤维素含量显著降低,而SC 和木质素含量则显著升高,L3 鞭长母鞭纤维素和SC 含量显著降低,而木质素含量则显著升高(表3)。
鞭长显著影响菲黄竹新鞭纤维素、SC 含量,鞭径显著影响新鞭木质素含量,鞭长和鞭径的交互作用对SC 及其组分含量均有显著影响(表1)。随鞭长的增大,D1 鞭径新鞭SC 及其组分含量总体呈升高趋势,D2鞭径新鞭SC及其组分含量均无明显变化。随鞭径的增大,L1鞭长新鞭SC组分含量显著升高,L2、L3 鞭长新鞭纤维素含量显著降低,而木质素含量则显著升高,且SC 含量无显著变化(表3)。表明,鞭长和鞭径对菲黄竹母鞭、新鞭SC 及其组分含量均有明显影响,且对母鞭的影响更明显,反映出新鞭碳水化合物优先供应的生长机制。
3 讨论与结论
3.1 鞭长和鞭径对菲黄竹鞭段繁育容器苗地下鞭C、N、P养分化学计量特征的影响
C、N、P 均是植物体的主要组成元素,植物C、N、P 化学计量特征在一定程度上反应了植物养分的限制状况及其适应策略。同时,N 和P 也是陆地生态系统生产力最重要的两个限制元素,对植物生长和各种生理机能的调节起着非常重要的作用[22]。本研究发现,不同鞭径,菲黄竹母鞭C、N、P含量随鞭长增大的变化规律不同。随鞭长的增大,D1 鞭径菲黄竹母鞭C 含量显著升高,这说明鞭段长度增大,母鞭内C积累量较高,维持新竹—竹鞭生长的碳的消耗比例相对较小;同时,鞭长增大也导致立竹地上部分生长相对旺盛,碳同化能力增强,C 同化物向库器官的运输增多。而母鞭N、P 含量明显降低,这一方面可能是由于鞭段长度增大,立竹的快速生长导致土壤N、P 的不平衡吸收和利用效率提高,使传输至竹鞭N、P 含量降低[23];另一方面可能是因为母鞭存储和鞭根吸收的N、P 更多地用于菲黄竹地上部分生长、母鞭自身消耗以及竹鞭生长的稀释作用引起[24](表4)。可见,细鞭条件下,如果母鞭鞭长过短,C 供应不足、竹苗地径、苗高、分株数量较低、地上部分生长较差、长势不佳(表4)。但如果母鞭鞭长过长,竹鞭基础代谢高,N、P 消耗大,并不一定会对竹鞭生长有正向作用,因此可以认为D1L2 处理相较于D2L1、D2L3 处理更利于鞭段容器苗生长。随鞭长的增大,D2 鞭径菲黄竹母鞭C 含量呈降低趋势,这可能与容器苗生物量明显增大的C 大量消耗等密切相关(另文待发表),由此可以说明,D2L1 处理鞭段因生物量较低,只有积累足够的C 才能来支持新鞭的良好生长。但母鞭N、P 含量明显升高,可能与母鞭自身生长、新鞭萌发以及地上部分迅速生长的大量养分需要,从而增加了对土壤N、P 元素的吸收等有关(表4)。可见,D2 母鞭鞭长过短,鞭根吸收N、P 含量较低,母鞭鞭长较长C 含量不充足,可以认为D2L2 处理相较于D2L1、D2L3 处理更有利于竹鞭生长。随鞭径的增大,L2 鞭长母鞭虽C 含量显著降低,但N、P 含量显著升高,而且新鞭生物量也显著升高,地上部分生长旺盛,综合分析认为D2L2 较D1L2 竹苗生长更好。不同鞭长、鞭径处理菲黄竹新鞭C、N、P 含量均无明显差异,表明不同鞭长和鞭径处理均能支持新鞭的正常生长。

表4 鞭长和鞭径处理下菲黄竹鞭段繁育容器苗生长状况Tab.4 Growth condition of breeding container seedlings of Sasa auricoma rhizome segments under rhizome length and rhizome diameter treatment
植物叶片的C∶N、C∶P 意味着植物吸收营养所能同化C 的能力,在一定程度上反映了植物的养分利用效率,因而具有重要的生态学意义[25]。本研究发现,随鞭长的增大,D1 鞭径母鞭C∶N、C∶P 均显著增大,说明立竹的生长活力提高,对N、P 利用效率提高,这在一定程度上缓解了立竹生长消耗所造成的土壤养分有效性降低对菲黄竹个体生长的限制[26];D2 鞭径母鞭C∶N、C∶P 均呈降低趋势,说明N、P 的利用效率降低,体现了菲黄竹适应养分的一种生存策略。不同鞭长、鞭径N∶P 均无明显变化,较N、P 含量更加稳定,反映了菲黄竹鞭系N、P含量的相对一致性,保持较高的养分内稳性以保障菲黄竹正常的生长发育和生理代谢[27]。
3.2 鞭长和鞭径对菲黄竹鞭段繁育容器苗地下鞭碳水化合物的影响
碳水化合物是植物光合作用产物和呼吸作用的底物,它为植物生长发育提供能量和碳构架。植物体内碳水化合物组分水平通常可反映植物整体的碳供应水平,表征植物生长状况及其对外界干扰、胁迫的缓冲能力与适应策略[28]。本研究发现,随鞭长的增大,D1 鞭径母鞭的可溶性糖含量呈“∧”变化规律,而淀粉含量呈降低趋势,SC 及其组分含量均呈升高趋势,表明适当增加鞭段长度,使得立竹活性增加,动员大量淀粉的活化与分解[29],一部分转化为可溶性糖用于呼吸、生命活动,另外合成纤维素等结构物质用于增加竹鞭机械能力以利于其在土壤中穿行和新鞭的扩展[30]。随着鞭长的增大,D2 鞭径母鞭的可溶性糖含量呈“∧”变化规律,淀粉和NSC 含量均总体呈显著升高趋势,SC 含量呈降低趋势,一方面表明适当增加鞭段长度,地上部分生长旺盛,光合碳同化能力增强,光合同化物向库器官的运输增加,另一方面表明C 从结构性成分更多地向活性代谢物质糖类分配,为维持地下鞭生物量增加和器官的加快生长提供足够的底物[31]。可见,适当增加鞭段长度,立竹生长代谢活性增加,立竹生长状况较好。而随着鞭径的增大,L2 鞭长母鞭可溶性糖含量无明显变化,而淀粉、NSC 含量显著下降,SC 含量均显著增大,这可能与鞭的持续生长,母鞭内的淀粉转移至新鞭,供新鞭生长有关[32],或者是NSC 更多地参与到形态建成。
菲黄竹新鞭的碳水化合物含量随鞭长的变化存在一定差异。随着鞭长的增大,D1鞭径新鞭NSC 含量呈下降趋势,SC 及其组分含量总体呈升高趋势,表明NSC 更多地参与到形态建成,碳消耗增加,体现新鞭生长过程中碳平衡的自我调节;D2 鞭径新鞭碳水化合物组分含量无明显变化。而且不同鞭段长度,菲黄竹新鞭碳水化合物含量随鞭径的增大总体上无显著变化,表明,鞭长和鞭径对菲黄竹鞭段容器苗地下鞭碳水化合物的影响主要体现在母鞭上,也反映出碳同化物优先供应新鞭的生长机制。
综上,鞭长和鞭径对菲黄竹鞭段容器育苗的母鞭主要养分含量及其化学计量比有明显的影响,并存在显著的交互作用,鞭径效应具有明显的鞭长水平依赖性,受养分稀释作用的调节,但对新鞭的影响较小,存在新鞭的养分优先供应的生长机制。本试验中,6 种不同鞭长鞭径处理均能支持菲黄竹新鞭的正常生长,其中,母鞭鞭径5 mm、鞭长6 cm 的鞭段处理总体上容器苗地上部分生长情况较好,地下鞭养分含量和生物量均较高,考虑到容器苗生长质量和育苗鞭段的经济节约,建议菲黄竹容器育苗时选择鞭径5 mm、鞭长6 cm左右的鞭段。
猜你喜欢
中学生数理化·八年级物理人教版(2022年4期)2022-04-26
煤气与热力(2021年12期)2022-01-19
湖北林业科技(2021年4期)2021-09-07
山东林业科技(2020年6期)2020-01-01
读者·校园版(2019年24期)2019-12-10
中成药(2018年8期)2018-08-29
中成药(2018年2期)2018-05-09
自然保护地(2017年2期)2017-06-26
小朋友·聪明学堂(2015年8期)2015-11-30
云南林业(2015年1期)2015-02-10
