
基质金属蛋白酶-2、9基因多态性与子宫内膜息肉发病风险的研究
2020-09-01 08:31唐世倩李晟辉褚春芳刘菊红李珊珊
中日友好医院学报 2020年3期
唐世倩,李晟辉,褚春芳,刘菊红,李珊珊,赵 虹
(首都医科大学附属北京妇产医院 妇科,北京 100026)
子宫内膜息肉(endometrial polyps,EP)是子宫内膜的良性病变,也是妇科常见疾病,可发生于青春期后的任何年龄,随着年龄增加发病率呈现上升趋势[1~3]。EP 可以引起异常子宫出血、不孕等多种问题,因此越来越受到人们的关注。
研究发现,参与机体炎症表达的基质金属蛋白酶-2 (MMP-2)、MMP-9 在EP 组织中表达升高,推测其可能参与了EP 的发生发展,但其作用机理并不清楚。众多研究表明多种疾病中MMP-2及MMP-9 的改变均与其基因启动子区域位点的多态性相关,而在EP 组织中是否也存在MMP-2及MMP-9 基因多态性的改变,尚未见报道。因此,本研究将探讨MMP-2 基因启动子区-735C/T位点及MMP-9 基因启动子区-1562C/T 位点多态性与EP 的相关性,以期深入了解EP 发生机制。
1 材料与方法
1.1 研究对象
收集2017年4月~2019年8月在首都医科大学附属北京妇产医院就诊,行手术治疗,经病理证实为子宫内膜息肉组织标本82 例,作为实验组。以年龄为匹配条件的同期于首都医科大学附属北京妇产医院行子宫内膜活检及正常绝经后妇女因子宫脱垂行子宫切除术的标本51 例,经病理证实为正常子宫内膜组织,作为对照组。所有研究对象既往均未使用激素药物治疗,且无甲状腺、子宫肌瘤、恶性肿瘤、糖尿病、多囊卵巢综合征等病史。本研究经伦理委员会批准,所有研究对象在纳入试验前均被详细告知本研究的目的和研究方法,并签署了知情同意书。
1.2 标本采集及DNA 提取
组织标本均采用传统的酚-氯仿抽提法提取基因组DNA,经紫外分光光度计测DNA 浓度及纯度。
1.3 MMP-2 基因分型
采用聚合酶链反应限制性片段长度多态性技术(PCR-RFLP)法对MMP-2 启动子-735C/T 位点单核苷酸多态性进行基因分型。扩增MMP-2启动子-735C/T 位点的上游引物为 5' -ATAGGGTAAACCTCCCCACATT- 3';下游引物为5'-GGTAAAATGAGGCTGAGACCTG-3'。PCR 反应条件为:94℃预变性5min,94℃变性45s,65.5℃退火45s,72℃延伸45s,经35 个循环,72℃总延伸7min。PCR 产物经限制性内切酶Hinf I(TakaRa生物工程(大连)有限公司合成),于37℃16h 酶切后,于3.0%的琼脂糖凝胶中电泳,电压85V,30min。电泳后,紫外灯下观察结果,记录并绘制电泳图谱。PCR 产物 为300bp,T/T 基因型因 存在Hinf I 的识别位点,产生254bp 和46bp 2 条DNA片段,C/C 基因型因缺乏Hinf I 的识别位点保持原有PCR 产物300bp 片段,C/T 基因型则显示为300bp、254bp 和46bp 三条片段(图1)。
1.4 MMP-9 基因分型
采用PCR-RFLP 法对MMP-9 启动子-1562C/T 位点进行单核苷酸多态性基因分型。扩增MMP-9 启动子-1562C/T 位点上游引物为5'-GCCTGG CAC ATA GTA GGC CC-3',下游引物为5'-CTT CCT AGC CAG CCG GCA TC-3'。PCR反 应条 件为:94℃预变 性5min,94℃变 性35s、63.5℃退火40s,70℃延伸50s,经40 个循环,72℃总延伸5min。PCR 产物经限制性内切酶SphI(TakaRa 生物工程大连有限公司合成) 于37℃恒温水浴过夜约16h 酶切后,进行3.0%琼脂糖凝胶电泳,电压85V,30min。电泳后,紫外灯下观察结果,记录并绘制电泳图谱。PCR 产物为435bp,T/T基因型产生188bp 和247bp 两条DNA 片段,C/C基因型因缺乏识别位点保持原有PCR 产物435bp 片段,C/T 基因型有435、247、188bp 3 个片段(图1)。
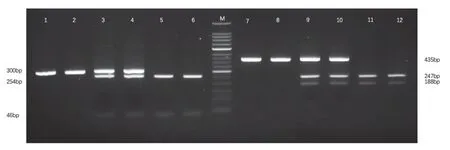
图1 MMP-2、MMP-9 酶切后PCR 扩增产物
1.5 统计学方法
应用SPSS17.0 统计软件,所测基因型和等位基因频率均采用基因直接计数法计算,Hardy-Weinberg平衡检验检测对照组和病例组中各基因型频率,确定选择对象的随机性。检验比较正常对照组与病例组间基因型和等位基因分布频率的差异。经年龄矫正,以非条件Logistic 回归法计算优势比(OR)及其95%可信区间(CI)表示相对风险度。
2 结果
2.1 研究对象的一般特征
病例组平均年龄为49.37±1.32 岁,对照组平均年龄44.17±1.69 岁,2 组相比较差异无统计学意义 (P>0.05),具有可比性。病例组及对照组MMP-2 启动子-735C/T 位点基因型分布符合Hardy-Weinberg 遗传平衡定律(均P>0.05),病例组及对照组MMP-9 启动子-1562C/T 位点基因型分布也符合Hardy-Weinberg 遗传平衡定律 (均P>0.05)。
2.2 MMP-2 启动子-735C/T 位点基因型及等位基因频率分布比较
在子宫内膜息肉组织中,MMP-2 启动子-735C/T 位 点T/T、C/T、C/C基因型频率分别为42.68%、51.22%、6.10%,与对照组比较差异有显著性(P<0.05)。与C/T、C/C 基因型相比,T/T 基因型使子宫内膜息肉的发病风险增加约2.7 倍(OR=2.708;95%CI 1.219~6.014)。并且T、C 等位基因频率分布差异有显著性 (P<0.05)(表1)。-735C/T 位点T 等位基因使子宫内膜息肉的发病风险增加约2.8 倍 (OR=2.839;95%CI 1.377 ~5.853)。
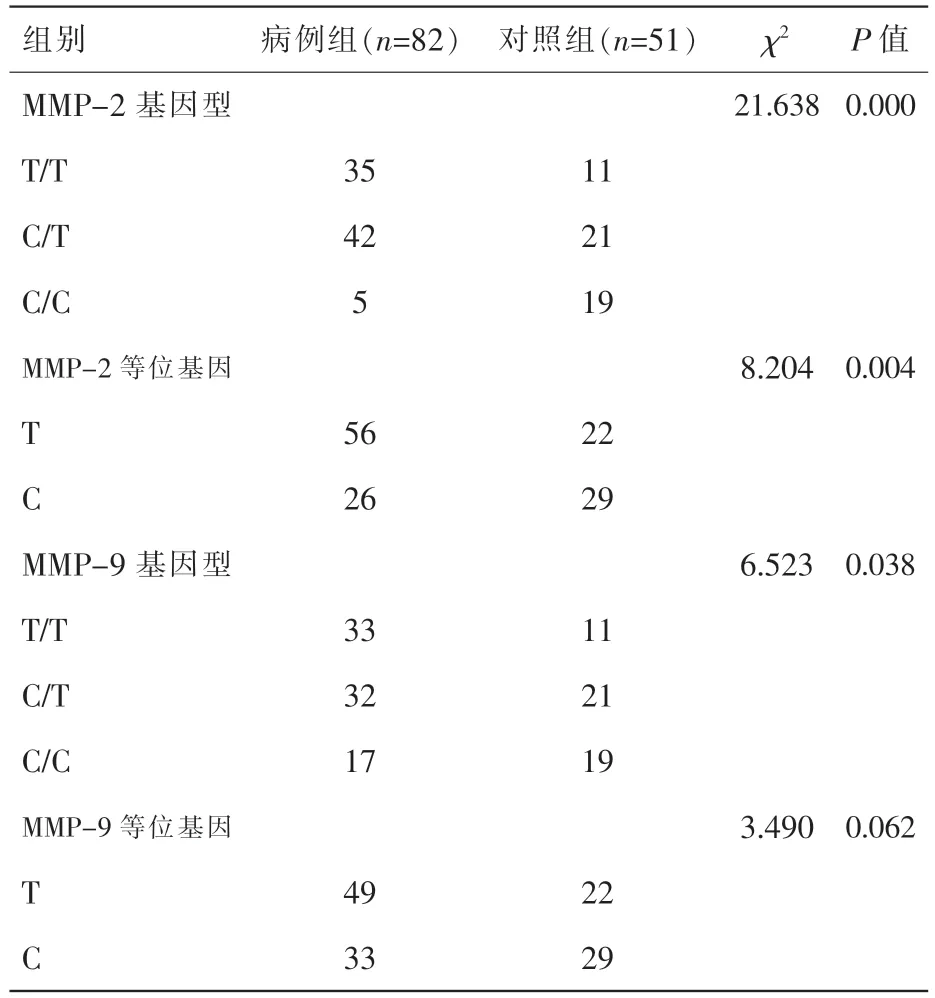
表1 2 组研究对象MMP-2、MMP-9基因型及等位基因的分布
2.3 病例组及对照组MMP-9 启动子-1562C/T位点基因型及等位基因频率分布比较
在子宫内膜息肉组织中,MMP-9 启动子-1562C/T 位 点T/T、C/T、C/C 基因型频率分别为40.24%、39.02%、20.73%,与对照组比较差异有显著性(P<0.05)。与C/C 基因型相比,C/T、T/T 基因型使子宫内膜息肉的发病风险增加约2.3 倍(OR=2.270;95%CI 1.041~4.949)。但T、C 等位基因频率分布差异无统计学意义(P<0.05)(表1)。
3 讨论
子宫内膜息肉由子宫内膜腺体,间质和血管组成,是子宫局部内膜过度增殖引起的内膜病变,发病率约1%~8%[4],较小的息肉可无临床症状,仅在行超声检查时发现,但体积较大的息肉可引起不规则阴道出血、不孕等多种临床症状,部分患者子宫内膜息肉反复发生,甚至会发生恶变[5,6],严重影响人们健康。
迄今为止,子宫内膜息肉的发生机制并不完全明确,目前普遍认为其发生原因与子宫内膜的炎症、体内激素环境紊乱、细胞因子及受体失调,细胞增殖凋亡失衡相关,其中炎症因素受到人们的广泛关注,当子宫内膜出现炎症时,内膜腺上皮和间质细胞致炎因子、血管形成因子的表达上调,从而引起内膜增生,息肉形成。
基质金属蛋白酶(MMPs)是目前研究较多的与机体炎症改变相关的酶类,它是一类依赖于锌、钙的内源性蛋白水解酶,主要由内皮细胞、血管平滑肌、巨噬细胞等产生[7],目前已经发现的MMPs有26 种,其主要生理功能是降解细胞外基质成分,维持细胞外基质的动态平衡,参与人体的生理及病理过程[8,9]。正常生理情况下,MMPs 在机体内表达量极低,而在胚胎形成、炎症、新生血管生成、肿瘤细胞的生长和转移等条件下表达水平显著升高[10]。
MMP-2、MMP-9 均为基质金属蛋白酶家族的重要成员,他们由肝细胞分泌,属于明胶酶,主要作用为降解基膜的IV 型胶原,参与炎症的发生及扩散,血管的形成等病理变化[11,12]。为了明确MMP-2、MMP-9 是否与子宫内膜息肉这一疾病发病相关,2014年张希奇等人研究发现与正常子宫内膜组织相比,在子宫内膜息肉组织中,MMP-2、MMP-9 均呈现出高表达[13],2016年,张勤等人再次验证了这一结果[14],这些研究表明MMP-2、MMP-9 在子宫内膜息肉组织中表达升高,但引起其升高的原因尚不清楚。
作为酶类,MMPs 的活性可受多个位点调节,研究证实MMPs 启动子区多态性位点在人群中的分布可能会导致MMP 表达水平的差异,导致某些疾病在人群中的易感性不同[15]。MMP-2 基因启动子区域-735C/T 位点单核苷酸多态可导致内转录因子SP1 结合区的序列发生改变,阻碍了启动子SP1 结合区结合特异的核内蛋白,从而改变基因转录的活性及水平[16],研究发现MMP2 启动子区-735 C/T的C/C 基因型与食管癌风险和转移风险增加相关[17],C 等位基因与乳腺癌风险增加有关[18],当T/T 基因型与C/T 基因型存在时,膀胱癌风险增加[19]。此外,研究还发现MMP-2 启动子-735C/T 位点单核苷酸多态性与骨髓增生疾病[20]、卵巢癌[21]等疾病具有相关性。本实验中通过对实验组和对照组组织的MMP-2 启动子区-735C/T位点进行基因多态性检测,结果发现两组间等位基因频率具有显著差异性(P<0.01),其中T/T 基因型、T 等位基因均明显增加子宫内膜息肉的发病风险,OR 值分别为2.708、2.839,这表明MMP-2 启动子-735C/T 位点多态性的改变可以影响子宫内膜息肉的患病风险。
同样,1999年Zhang 等人[22]发现MMP-9 基因启动子区域存在-1562C/T 多态性,其位点位于MMP-9 基因启动子的转录起始位点,同时是转录抑制因子所识别的位点。C/T 的改变可破坏转录抑制因子的识别以及结合的序列位点,因此,具有显著增强启动子活性的作用[23]。但目前发现,MMP-9 基因启动子区域-1562C/T 位点上两个等位基因在不同疾病中的作用不同。2007年Sfar 等人发现C 等位基因可降低前列腺癌的发病风险,而T 等位基因则会将前列腺癌发病风险提升3倍[24],同年Rollin 等人发现,C 等位基因与肺鳞状细胞癌的风险增加有关[25]。在本研究中,子宫内膜息肉组织标本与正常子宫内膜组织标本相比较,MMP-9 启动子-1562C/T 位点等位基因的频率具有显著性(P<0.05),与C/C 基因型相比,C/T、T/T基因型可使子宫内膜息肉的发病风险增加2.270倍,由此推断C/T、T/T 基因型与子宫内膜息肉的易感性正相关。
本实验通过聚合酶链反应限制性片段长度多态性技术检测方法,首次探讨了MMP-2 基因启动子区域-735C/T 及MMP-9 启动子-1562C/T 基因多态性与子宫内膜息肉之间发病关系,结果发现不同的基因型及等位基因会影响子宫内膜息肉易感性,这一结果将为进一步探讨子宫内膜息肉的发病机制提供理论依据。
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22
健康博览(2022年7期)2022-07-08
川北医学院学报(2022年6期)2022-06-24
昆明医科大学学报(2022年2期)2022-03-29
保健与生活(2021年20期)2021-10-15
昆明医科大学学报(2021年6期)2021-07-31
智慧健康(2021年17期)2021-07-30
昆明医科大学学报(2021年3期)2021-07-22
保健与生活(2021年11期)2021-06-10
中国产前诊断杂志(电子版)(2020年1期)2020-05-21
