
紫金山针阔混交林主要植物种群生态位特征
2020-08-31 04:14袁在翔金雪梅关庆伟戴康龙
中南林业科技大学学报 2020年8期
陈 霞 ,袁在翔 ,金雪梅 ,关庆伟 ,朱 军 ,戴康龙 ,赵 宸
(1. 中山陵园管理局,江苏 南京 210014;2. 南京林业大学 a. 生物与环境学院;b. 南方现代林业协同创新中心,江苏 南京 210037)
生态位是指物种在生物群落或生态系统中的功能关系以及所占据的时、空上的特殊位置[1],能够反映物种之间、物种与环境之间的相互关系[2],是解释自然群落中种间共存与竞争机制的基本理论[3]。生态位宽度和生态位重叠度是生态位理论中常用的主要量化指标[4-5]。研究森林主要种群的生态位特征,有助于了解森林主要植物的资源利用能力以及它们所占据的生态空间的范围和相对地位[3]。近年来,国内学者对中亚热带和南亚热带的森林群落中种群生态位进行了一些研究[3,6-8],但对北亚热带次生老龄林植物群落种群生态位的研究较为匮乏。
紫金山针阔混交林是在人为干扰相对较弱的情况下,经过65 a 左右的演替,由栓皮栎Quercus variadilis、朴树Celtis sinensis等阔叶树种入侵马尾松Pinus massoniana纯林而形成的,在我国北亚热带地区具有一定的代表性[9-10]。当前已有对紫金山针阔混交林的研究多集中在演替动态[10]、空间结构[11-12]、植物多样性[12-13]、土壤性质[14-15]等方面,对群落主要物种的生态位研究较少。因此,本研究以紫金山65 a 林龄的针阔混交林为研究对象,分析了10 种乔木、10 种幼树、7 种灌木、6 种藤本、8 种草本的生态位特征,旨在明晰群落各层次主要物种对环境资源的生态适应性以及种间竞争性,探讨物种共存与多样性维持的潜在机制,为紫金山森林资源监测与管理以及次生林的恢复与重建提供一定的参考价值。
1 研究区概况
紫金山(118°48′24″~118°53′04″E,32°01′57″~32°06′15″N)位于南京市玄武区,主峰海拔高度448.9 m,相对高度420 m,总面积2 970 hm2,属于北亚热带季风气候,四季分明,年降水量1 000 ~1 050 mm,年均气温15.4 ℃,年均日照2 213 h,全年无霜期322 d。土壤类型以黄棕壤和灰棕壤为主,酸性至微酸性。林分类型有落叶阔叶林、马尾松纯林和针阔混交林。其中,针阔混交林主要乔木为马尾松、栓皮栎、朴树、紫薇Lagerstroemia indica,主要灌木为山莓Rubus corchorifolius、菝稧Smilax china、六月雪Serissa japonica、雀梅藤Sageretia thea,主要草本植物为苔草Carex chinensis、 麦 冬Liriope graminifolia、 狭 叶 鳞毛 蕨Dryopteris angustifrons、 野 青 茅Deyeuxia arundinacea。
2 材料与方法
2.1 样地设置
于2014 年10 月,在紫金山南麓的针阔混交林中用全站仪和罗盘仪划定典型样地1 hm2,样 地 内 GPS 测 定 点 的 位 置 为 118°50′23.7″E,32°04′03.9″N,海拔 230 m,上坡位。运用相邻格子法把典型样地划分为25 个20 m×20 m 的调查样方。在每个调查样方内,进行每木调查,对样地内胸径≥5 cm 的树木分别进行定位,记录所研究林分的每株个体的位置,以坐标值表示,分别记录每株林木的树种、胸径(≥5 cm)、树高、冠幅、坐标定位,坐标值用距离(m)直接表示,并在每个调查样方内4 个角及中心设置2 m×2 m小样方,调查小样方内的更新幼树、灌木、草本和藤本植物,记录植物的种名、基径、平均高度、盖度等指标。
2.2 试验指标的测定
2.2.1 植物重要值的测定
通过重要值(I.V.)大小来描述乔木层、乔木更新层、灌木层、藤本层、草本层的植物种群优势度,进而确定各层片中主要植物种群。重要值具体计算公式为:1)胸径≥5 cm 乔木树种的重要值=相对密度+相对显著度+相对频度,相对密度=(样方内某一树种的个体数/样方内全部树种的个体数)×100%,相对显著度=(样方内某一树种的所有个体胸高断面积总和/样方内全部树种的所有个体胸高断面积总和)×100%,相对频度=(某一树种的频度/所有树种的频度总和)×100%[16];2)更新幼树、灌木、草本和藤本植物的重要值=相对盖度+相对频度,其中相对盖度=(某层次内某种植物的盖度/该层次内全部植物种类盖度之和)×100%,相对频度=(某层次内某种植物的频度/该层次内全部植物种类频度之和)×100%[1]。
2.2.2 资源轴的确定
资源轴的定义主要有以下3 种方式[17]:1)食物资源,不同种类的食物和不同食物的数量等。2)生境资源,生物学上或物理化学(温度,湿度,pH 值,……)上定义的一系列状态。3)抽样单位,一组自然或人为的抽样单位。本研究用人为的抽样单位来定义资源轴,同时为了体现所有层次植物种群的资源利用状态,故选择以20 m×20 m 的乔木调查样方为资源轴。
2.2.3 生态位宽度的计算
由于无法确定样地内资源状态的丰富度和可获得性,故而选用对丰富的资源有更大权重的Levins[18]生态位宽度(BL)和对稀缺资源有更大权重的Shannon-Wiener[19]生态位宽度(BSW)来描述各层次主要植物种群的生态位宽度,BL 和BSW 的具体计算公式为:
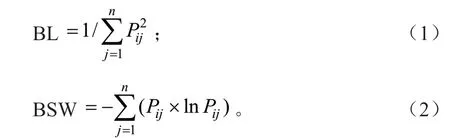
式(1)~(2)中:n为资源轴个数即乔木调查样方数;Pij为物种i在第j个资源状态下的重要值占该种所有资源状态下的重要值总和的比例。
2.2.4 生态位重叠度的计算方法
生态位重叠度能反映出物种对资源利用性质的相似性,重叠度越大,两物种对资源利用性质越相似,反之亦然[20]。运用Pianka[21]生态位重叠度公式来计算各层次主要植物种群之间的生态位重叠度,具体计算方法如下:

式(3)中:Oik为物种i和物种k之间的生态位重叠度;n为资源轴个数即乔木调查样方数;Pij、Pkj分别为物种i和物种k在第j个资源状态下的重要值占该种所有资源状态下的重要值总和的比例。此外,各层次主要物种总体的生态位重叠度以每层次内所有主要物种对的生态位重叠度算术平均值来表示。
2.3 数据分析
用Excel 2016 软件整理样方每木调查的胸径、树高、冠幅、盖度等原始数据,计算每种植物的重要值;通过 R 3.4.1 计算样方主要物种的生态位宽度、生态位重叠度以及各层次生态位宽度、生态位重叠度的均值和标准差。
3 结果与分析
3.1 主要物种生态位宽度
根据典型样地植物的重要值大小顺序,选择10 种乔木、10 种幼树、7 种灌木、6 种藤本、8 种草本作为各层次的主要物种,并分别计算它们的生态位宽度,其结果见表1。由表1 可知,各层次主要物种Levins 和Shannon-Wiener 生态位宽度的计算值存在一定差异,但物种之间的大小排列顺序基本一致。各层次最大生态位宽度的物种分别是乔木层的马尾松(19.58、3.05)、乔木更新层的白檀(17.34、2.92)与朴树(15.85、2.94)、灌木层的山莓(18.78、3.02)、藤本层的络石Trachelospermum jasminoides(22.43、3.14)、 草本层的苔草(18.25、3.02),表明这些物种在该生境中资源利用能力强,分布幅度较广。最小生态位宽度的物种分别是乔木层的短柄枹Quercus glandulifera(11.71、2.62)、乔木更新层的油茶Camellia oleifera(4.40、1.64)、灌木层的狭叶山胡椒Lindera angustifolia(2.54、1.00)、藤本层的海金沙Lygodium japonicum(4.37、1.76)、草本层的南山堇菜Viola chaerophylloides(4.88、1.83),表明这些物种在该生境中资源利用能力弱,生态适应性较差。
标准差(SD)能反映组内个体的离散程度。各层次主要物种生态位宽度的SD 大小顺序为:藤本层(6.74、0.51)>灌木层(5.91、0.70)>草本层(4.99、0.44)>乔木更新层(4.31、0.41)>乔木层(2.62、0.15),表明乔木层生态位宽度离散度较小,主要物种对生境资源的利用能力相差不大,而藤本层、灌木层、草本层的情况则相反。此外,仅藤本层主要物种生态位宽度的大小顺序与相对应的重要值大小顺序相同,其他层次均表现出不同程度上的不一致。例如,乔木层中栓皮栎、短柄枹等栎类植物的重要值排序靠前,而它们的生态位宽度值排序靠后;乔木更新层中白檀、青桐Firmiana platanifolia等植物的重要值排序较后,但它们的生态位宽度值排序较前。
3.2 主要物种之间的生态位重叠度
从表2 ~6 可知,各层次主要物种的生态位重叠度分别在0.26 ~0.85(乔木层)、0.01 ~0.70(乔木更新层)、0.00 ~0.74(灌木层)、0.13 ~0.77(藤本层)、0.15 ~0.67(草本层)之间,最大生态位重叠度的物种对分别为乔木层的朴树与山胡椒、乔木更新层的山胡椒和白檀、灌木层的六月雪与黑果菝稧Smilax glaucochina、藤本层的络石与地锦Euphorbia humifusa、草本层的苔草与野青茅Deyeuxia arundinacea,最小生态位重叠度的物种对分别为乔木层的短柄枹与石楠Photinia serrulata、乔木更新层的油茶与紫薇、灌木层的狭叶山胡椒与黑果菝稧、藤本层的木防己Cocculusorbiculatus与海金沙、草本层的麦冬与南山堇菜、荩草Arthraxon hispidus。各层次主要物种生态位重叠度的均值大小顺序以及各自的标准差分别为:乔木层(0.621、0.14)>藤本层(0.430、0.20)>乔木更新层(0.408、0.19)>灌木层(0.406、0.22)>草本层(0.364、0.16),表明乔木层主要树种对生境资源利用的性质更为相似,种间竞争的潜力较大。各层次生态位重叠度大于0.5 的物种对个数为:乔木层36 个、更新层16 个、灌木层7 个、藤本层5 个、草本层7 个;占比分别为:80%、35.56%、33.33%、33.33%、25%。此外,生态位重叠度高于0.8 的物种对均在乔木层,具体为:朴树与山胡椒(0.85)、朴树与紫薇(0.83)、紫薇与黄连木Pistacia chinensis(0.83)、紫薇与椤木石楠Photinia davidsoniae(0.81)、椤木石楠与冬青Ilex chinensis(0.80),并且乔木层的马尾松、朴树与其他主要树种之间的生态位重叠度均在0.5以上。
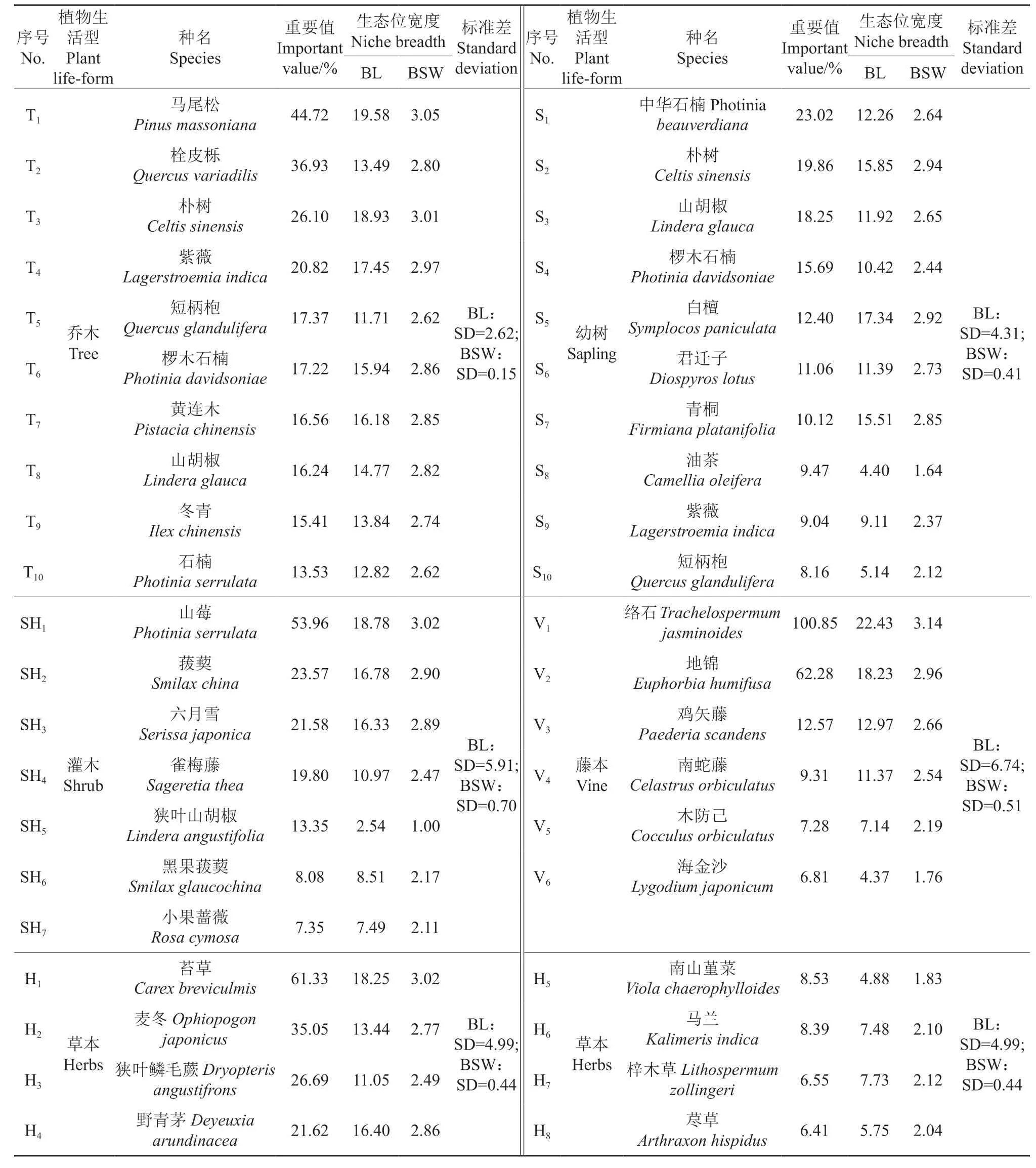
表1 针阔混交林各层次主要植物种群的生态位宽度†Table 1 Niche breadth of main plant populations of all layers in a coniferous and broad-leaved mixed forest

表2 针阔混交林乔木层主要种群的生态位重叠†Table 2 Niche overlap of main populations of tree-layer in a coniferous and broad-leaved mixed forest
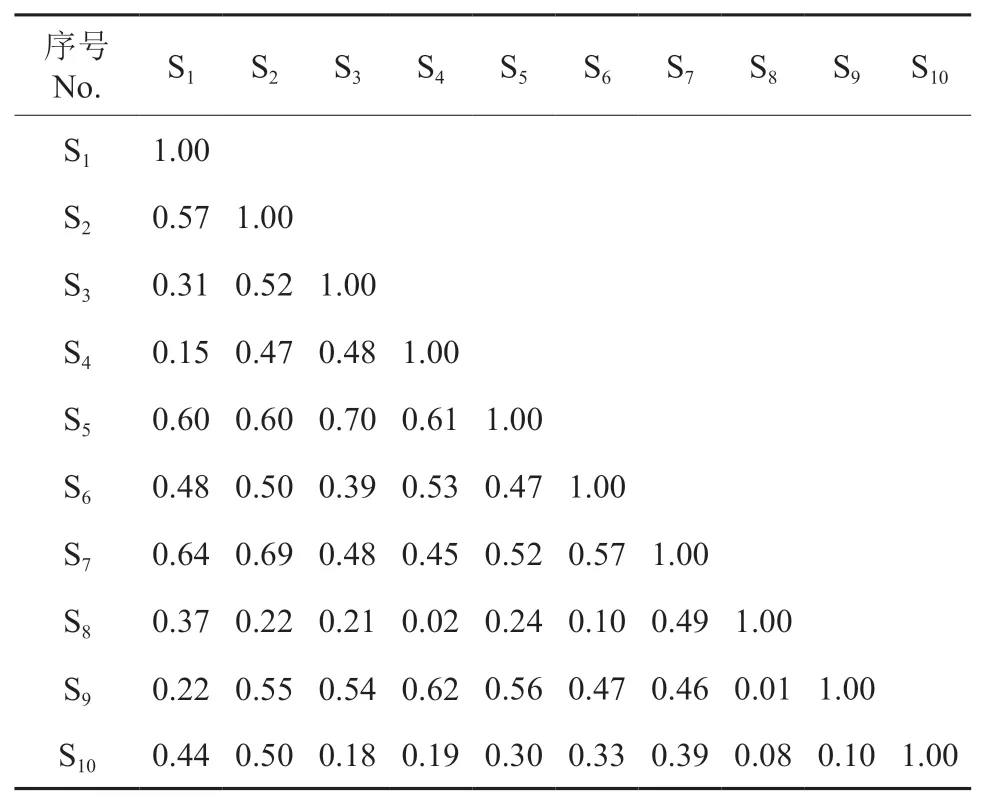
表3 针阔混交林乔木更新层主要种群的生态位重叠†Table 3 Niche overlap of main populations of sapling-layer in a coniferous and broad-leaved mixed forest
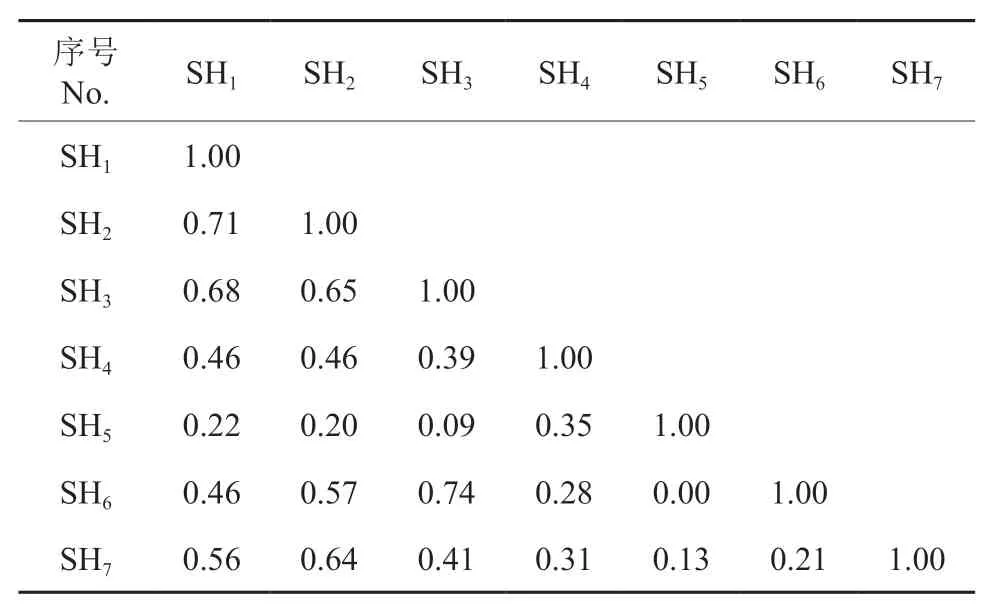
表4 针阔混交林灌木层主要种群的生态位重叠†Table 4 Niche overlap of main populations of shrub-layer in a coniferous and broad-leaved mixed forest

表5 针阔混交林藤本层主要种群的生态位重叠†Table 5 Niche overlap of main populations of vine-layer in a coniferous and broad-leaved mixed forest

表6 针阔混交林草本层主要种群的生态位重叠†Table 6 Niche overlap of main populations of herb-layer in a coniferous and broad-leaved mixed forest
4 讨 论
1)生态位宽度可以反映物种对资源的利用能力及其生态适应性和分布幅度[22]。在乔木层中马尾松生态位宽度最大,这是因为其喜光、耐贫瘠、喜微酸性土壤等生长特性有利于马尾松在紫金山微酸性土壤中广泛分布,并且作为早期植被恢复的先锋树种,经过长时间演替,大径阶个体是种群的主体部分,种群在时空上占据着优势地位,但是在更新层中因林冠郁闭导致马尾松幼苗更新困难,因而其更新层的生态位为零。白檀、朴树、山莓、络石、苔草均为稍耐阴、耐贫瘠、适应力较强的植物,其种群在群落演替过程中个体分布广泛而均匀,故而这些物种的生态位宽度也较大。乔木层中栓皮栎、短柄枹等栎类植物是北亚热带地带性优势种,能在演替过程中形成广泛的分布,但其种子较大,散播能力有限,并且是森林中啮齿类动物的取食对象,故而种群个体分布的均匀度较差,所以其生态位宽度较小。油茶、狭叶山胡椒、海金沙、南山堇菜等种群的个体分布范围小,仅在少数调查样方出现过,且在各分布样方之间的均匀度较差,所以种群的生态位趋于特化。此外,生态位宽度和重要值均可反映物种在群落中的地位和作用[3,23],但二者在物种之间的表现方式略有不同。研究中乔木层、乔木更新层、灌木层、草本层均有一些物种的生态位宽度大小排序与其重要值大小排序不同,如栓皮栎、短柄枹、白檀、青桐等,这说明物种的生态位宽度不仅受种群的重要值影响,同时也与种群个体分布的广度和均匀度密切相关。
2)生态位重叠反映了物种对同一类生境资源的联合利用以及它们之间的空间配置关系[3]。两物种生态位重叠有以下5 种情况[1]:①内包生态位,其竞争结果为一种物种迫使另一种物种减少对公共资源的利用,或者将它排除在外;②等宽生态位重叠;③不等宽生态位重叠,即一种物种的生态位有较大比例被另一物种所共用,两物种的竞争关系不对等;④邻接生态位,两物种回避竞争;⑤分离生态位,一般不竞争。本研究中各层次主要物种的生态位重叠度均在0 ~1 之间,生态位宽度均大于1,且任意两物种的生态位宽度均不相同,故主要物种之间的生态位重叠多为不等宽生态位重叠,也有少量邻接生态位如狭叶山胡椒和黑果菝稧。此外,物种之间的生态位重叠度与其生态位宽度密切相关[5,21]。一般来说,生态位宽度较大的物种之间生态位重叠度也较大[5],反之亦然,但生态位宽度较大的物种与生态位宽度较小的物种之间生态位重叠度不一定较小[23]。本研究中也证实了这一点,例如,藤本层、草本层的最大生态位重叠度出现在生态位宽度前两位物种之间,乔木层、藤本层的最小生态位重叠度出现在生态位宽度后两位物种之间,但大多数生态位重叠度的极值出现在生态位宽度相差较大的两物种之间,如乔木层的朴树与山胡椒,乔木更新层的山胡椒和白檀、油茶与紫薇,灌木层的六月雪、狭叶山胡椒与黑果菝稧,草本层的麦冬与南山堇菜、荩草等,这可能与物种自身的生物生态学习性有关[5]。
5 结 论
紫金山针阔混交林中马尾松、白檀、朴树、山莓、络石、苔草等物种在样地生境中资源利用能力较强,分布幅度较广;乔木层生态位宽度离散度较小,主要物种对生境资源的利用能力相差不大,而藤本层、灌木层、草本层的情况则相反;乔木层主要树种对生境资源利用性质的更为相似,种间竞争的潜力较大。
马尾松作为先锋树种,在演替过程中有显著的时空优势,占据着大量的环境资源,虽然在更新层中马尾松的生态位宽度为零,但当前演替阶段其种群多为大径阶个体,需要较长的演替时间被乡土落叶阔叶树种所取代。因此,可通过适当的森林经营措施来加速这一进程,缩短达到森林顶级群落的时间。
综上所述,本研究分析了紫金山针阔混交林当前演替阶段的主要植物种群的生态位特征,探讨了群落各层次优势种群对环境资源的生态适应性以及种间竞争关系,其结果对紫金山森林监测与管理具有一定的指导意义,也为南京丘陵地区以马尾松为先锋树种进行植被的恢复与重建提供一定的参考价值。此次研究中将抽样单位来定义资源轴,虽然体现了样方所有环境资源,但没有明确何种环境资源(如光照、土壤、水分、地形)是植物生态位分化的主要因素,今后可通过相关试验的主成分分析来进一步明确。
猜你喜欢
林业勘查设计(2022年1期)2022-02-15
乡村科技(2021年17期)2021-10-20
安顺学院学报(2021年4期)2021-09-16
青年文学家(2021年16期)2021-08-05
安徽农业科学(2021年7期)2021-04-29
少儿美术(2020年8期)2020-11-06
老年教育(2020年5期)2020-05-02
少儿科学周刊·儿童版(2019年5期)2019-08-01
现代农业研究(2016年8期)2016-12-02
绿色科技(2014年4期)2015-01-06
