
金寨天马自然保护区山核桃群落的系统发育结构1)
2020-08-24 14:32:08王雷宏王灿辉张华莲杨俊仙刘俊龙曹翠萍
东北林业大学学报 2020年7期
王雷宏 王灿辉 张华莲 杨俊仙 刘俊龙 曹翠萍
(安徽农业大学,合肥,230036) (安徽省林业科学研究院) (安徽农业大学)
群落系统发育结构是把物种的系统进化和群落生态过程整合在一起,探索群落物种多样性维持、物种共存的机制[1-4]。建立代表区域物种库的系统进化树,计算群落内物种间的系统发育距离,与从系统进化树上随机抽取物种的零群落的系统发育距离比较,获得系统发育结构。一般而言,近缘物种具有相似的生态习性,若环境容量有限,相互间强烈竞争、排斥,最终会形成由亲缘关系较远的物种组成群落,即系统发育结构发散的群落[5-6];若环境选择了生态习性相似的近缘物种组成了群落,即形成系统发育结构聚集的群落[7];如果物种从区域种库随机进入局域群落,群落系统发育的结构与随机期望的情况无显著差别,属于中性群落[4,8]。虽然群落系统发育结构随空间尺度、分类群尺度等不同尺度而变化,但在同一尺度下,群落间进行比较时,本质上反映群落的物种组成、系统进化的信息,而且系统发育结构指数反映群落与环境变化的对应关系,分析这种变化关系,可以揭示特定时空上群落构建的主导驱动力量[9-10]。Whitfeld et al.[11]研究发现新几内亚的热带森林演替的主要驱动力是环境筛选作用;Yan et al.[12]证实在亚高山森林群落构建中,竞争排斥是群落中物种共存的主要机制;在大尺度上,随着海拔或者纬度的增加,群落的系统发育结构聚集,环境筛选作用成为主要力量[13]。群落演替中期阶段环境过滤导致形态特征、习性相似的近缘物种大量聚集,系统发育结构聚集,达到环境的容纳量后,竞争排斥可能又成为主要的力量,导致群落系统发育结构发散,而群落演替的早期阶段,若环境选择压力很小,物种从区域种库进入局域群落为随机过程[14]。
分布于安徽大别山的山核桃(CaryacathayensisSarg.)[15-16]在不同地段有不同的群落组成,大部分是零散混生于其它物种建群的群落中,也有的群落建群种不明显,而在个别地段却以山核桃为建群种,地域内表现出演替系列,从群落系统发育的角度来看,群落具有不同的系统发育结构。因此,本研究的目的是探索安徽大别山天马自然保护区内分布有山核桃的群落系统发育结构以及变化规律,推测山核桃群落构建和维持的主导力量。
1 研究区概况
以安徽天马国家级自然保护区为调查范围,该区位于皖西大别山腹地,鄂、豫、皖三省交界处,地处东经115°20′~115°50′,北纬31°10′~31°20′。气候属于湿润季风气候,年平均气温13.8 ℃,7月份极端最高温度38.1 ℃,1月份极端最低温度-23 ℃。年平均降水量1 489 mm,主要集中在5—9月份。海拔800 m以下土壤为山地黄棕壤,800 m以上土壤为山地棕壤,山顶偶见草甸土。植被有落叶常绿阔叶林和落叶阔叶林,阔叶树主要以栓皮栎(QuercusvariabilisBl.)、麻栎(QuercusacutissimaCarruth.)、枫香(LiquidambarformosanaHance)、青冈(Cyclobalanopsisglauca(Thunb.) Oerst.)为主;针叶林以马尾松(PinusmassonianaLamb.)、黄山松(PinustaiwanensisHayata)为主;山地矮林以黄山栎(QuercusstewardiiRehd.)、华东椴(TiliajaponicaSimonk.)、四照花(Dendrobenthamiajaponicavar.chinensis(Osborn.) Fang)为主;山地灌丛以三桠乌药(LinderaobtusilobaBl. Mus. Bot.)、川榛(Corylusheterophyllavar.sutchuenensisFranch.)为主[17]。
2 研究方法
2.1 样地设计
于2018年7月进行了野外调查,在天马自然保护区共找到了6个典型的地段分布有山核桃,均在山体北坡至西坡向,溪谷岸边至中坡位地段,海拔高度在471~968 m。每个样地取20 m×20 m样方,记录了所有木本植物的名称、数量、平均胸径、平均高度(见表1)。

表1 样地生境概况及群系建群种
2.2 分析方法
采用V. PhyloMaker软件,生成了研究区域物种系统树,估算了各进化枝的分化年龄[18-21],分化年龄的单位为百万年。生成的系统树直接应用于Phylocom软件,结合样地数据计算系统发育多样性指数、系统发育结构指数、样地间平均系统发育距离[6,22]。传统的群落多样性指数以物种多度为属性,分别计算辛普森指数(D)、香农威纳指数(H′)、均匀度指数(J)[23]。分析群落间的系统发育亲缘关系,并进行非度量多维度分析(NMDS)排序;比较各群落的系统发育多样性指数与传统多样性指数的变化趋势;观测各群落的系统发育结构,并揭示其与群落演替阶段的对应关系。
系统发育多样性(PD)是每个群落的物种系统树上所有进化枝的分化年龄和,单位是百万年[22]。
净亲缘指数(I),它测度系统发育树上分类单元深层和浅层水平的系统发育亲缘关系,但测重于分类单元在目、科、属的系统发育亲缘关系。
式中:Is为某群落(样地)净亲缘指数,Ms为实际观测某群落(样地)中分类单元间平均系统发育距离,Mr为随机产生的零群落中分类单元间的平均系统发育距离,V为零群落中分类单元间系统发育距离的标准差。如果Is>0,表明群落系统发育结构聚集;如果Is<0,表明群落系统发育结构发散;如果Is≈0表明群落系统发育结构是随机的[5]。
3 结果与分析
3.1 群落间系统发育距离排序分类
现存的山核桃主要存在于其它植被群系,群系的分类主要依据群落的建群种,并不能反映群落间系统亲缘关系,计算群落间的系统发育距离,直接用于非度量多维尺度分析对样地排序(见图1)。由图1可知,样地1、样地3为一类,样地2为一类,样地4、样地5、样地6归为一类;从群落演替的角度来看,它们应处于不同的演替阶段,2号样地是山核桃顶级群落。
3.2 群落系统发育树
由图 2可知,6个样地中,共有133种(含有变种),物种的学名及系统学位置主要以植物名录的接受为准,保证同属的物种、同科的属、同目的科都相应地聚在了一起[24],枝梢代表种、变种,枝梢以上节点代表了种以上的各分类阶元,对各节点标注了分化年龄。一般枝梢到最近节点的长度代表了种或者变种的分化时间,继续往上的共同节点分别代表属、科、目,并标有分化年龄。有些属下物种的分化时间是不准确的,简单归并到属,如葛罗槭(Acerdavidiisubsp.grosseri)、五角枫(Acerpictumsubsp.mono)、三角枫(Acerbuergerianum)、毛果槭(Acernikoense)、阔叶槭(Aceramplum)、天童锐角槭(Aceracutumvar.tientungense)和青榨槭(Acerdavidii)等物种具有共同的节点,代表其属的分化年龄为-14.10×106a。检测群落的系统发育结构时,phylocom软件自动处理指定的零模型产生零群落,每次零群落的建立都是随机从这个系统树上抽取物种,并在这个树的基础上构建每个零群落的系统树,计算零群落平均系统发育距离。
3.3 群落系统发育多样性与传统多样性指数变化趋势的一致性
群落系统发育多样性指数不仅包含简单的物种丰富度的信息,而且反映了群落中物种的进化历史信息。为方便在同一数量级上观测5种指数随样地的变化趋势,对群落的物种丰富度取自然对数(lnS),对群落系统发育多样性指数取以10为底的对数(log10PD)。由表2可知,log10PD与香农威纳指数(H′)、lnS变化趋势基本一致;辛普森指数(D)和均匀度指数(J)变化趋势一致。
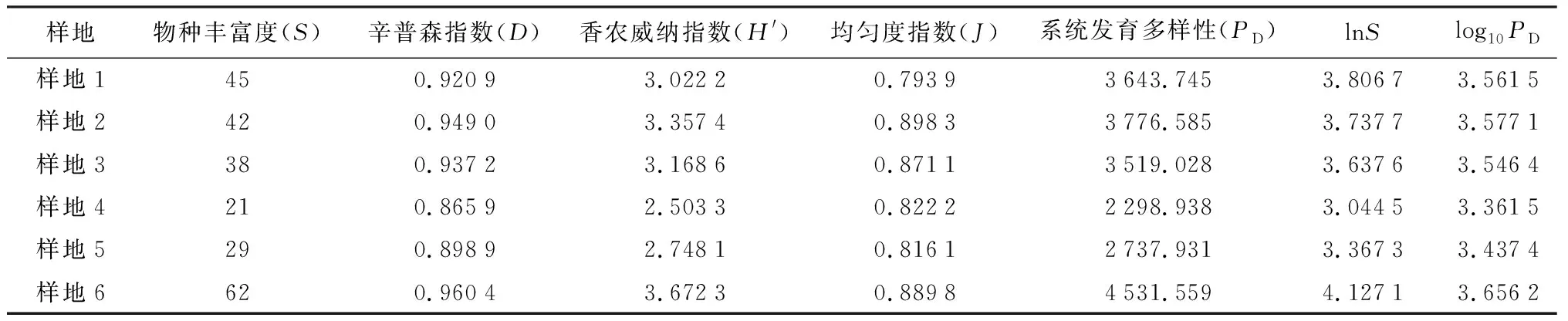
表2 各个样地的多样性指数
3.4 群落系统发育结构及与群落排序的关系
由表 3可知,样地4、样地5和样地6的净亲缘系数大于零,系统发育结构聚集;样地2的净亲缘系数小于零,系统发育结构均匀或者发散;样地1、样地3的净亲缘系数接近于零,两个样地系统发育结构随机。
发现净亲缘指数大于零的样地4、样地5和样地6在排序图上是一类,净亲缘系数小于零的2号样地在排序图是第二类,净亲缘系数接近于零的样地1、样地3在排序图上为第三类。

表3 用净亲缘系数检测群落系统发育结构
4 讨论与结论
群落系统发育多样性与群落香农威纳指数、物种丰富度指数的变化趋势基本是一致的。从系统发育多样性的原理来看,系统发育多样性是一个样地内所有物种构建的系统发育树上所有物种的枝长和,节点越多,物种的枝长越长,多样性就越高。所以,当样地的物种组成不一样时,系统树上节点不一样,各物种的枝长不一样,科、属起源古老,进化枝内物种分化复杂,其对应的各种的进化枝长和就会越大。例如样地1物种数虽然稍多于样地2,但是样地2系统发育多样性要高于样地1。
样地1、样地3群落系统发育结构随机,样地2系统发育结构发散,样地4、样地5、样地6系统发育结构聚集,与群落的排序是一致的。说明了群落的物种组成有一定的变化规律,对应一定的综合生态条件,而不是由某一个生态因子来主导。从山核桃的分布、样地的设置来看,并不是在连续的生态空间上抽取样地,样地是不连续的、比较零散或者孤岛式分布,体现不出明显的主导因子生态梯度。从时空转化的角度来看,反映的是山核桃所在群落的不同演替阶段,其系统发育结构不一样。样地2形成了以山核桃为主的建群种,生境是在较开阔的溪河谷地两岸,系统发育结构发散,说明了群落发展到顶级群落,山核桃及其相对亲缘物种达到了环境的极限,种间竞争、排斥作用维持着此类群落;样地1、样地3是群落的初级阶段,各物种随机进入样地;样地4、样地5、样地6的建群种分别是化香、枫香、榉树,山核桃在这些群落中虽然数量较少,但环境选择了习性相似的物种聚集,山核桃与这些相对亲缘物种共存于特定的生境中。
山核桃群落的系统发育多样性与传统的香农威纳指数在序列样地上的变化趋势是一致的,系统发育多样性不仅反映了群落的物种组成,还反映出了物种的进化复杂程度。山核桃群落系统发育结构与群落所处的演替阶段(以山核桃林顶级群落为参照系)有关联。群落演替的初期阶段,系统发育是随机的,物种随机进入群落,群落构建和维持的机制是中性随机过程;在群落演替的中期阶段,群落系统发育结构是聚集的,群落构建和维持的主要机制是生境的过滤,山核桃及其相对近缘类群具有相似的习性或者功能,共存于一定的生境内;群落演替的顶级阶段(相对于山核桃林)群落系统发育结构是发散的,山核桃及其相对近缘类群形态状性、功能性状、生态习性相似,种间竞争或排斥成了群落构建和维持的主要机制。因此,揭示了山核桃群落的构建和维持是物种进化、生态过程的共同作用,在群落的不同演替阶段有不同的主导作用。
猜你喜欢
作物学报(2022年2期)2022-11-06 12:11:02
中国现代中药(2021年9期)2021-11-16 07:43:32
数学大王·中高年级(2021年3期)2021-03-10 11:19:30
晚晴(2020年1期)2020-06-19 07:57:10
老年教育(老年大学)(2020年6期)2020-01-02 07:45:54
大学教育(2019年9期)2019-09-20 05:24:08
天然产物研究与开发(2018年10期)2018-11-06 07:43:46
广东农业科学(2017年5期)2017-08-29 10:37:26
作文与考试·小学高年级版(2015年5期)2015-05-30 10:48:04
食品科学(2013年14期)2013-03-11 18:25:06
