
Cloning and Expression Analysis of Heat Shock Protein Gene DpHsp70 in Dahlia
2020-08-15 13:35LanLUOHuaGUANSiyuZHOUSiyuZHUChuanleZHAOJianzhongTAN
农业生物技术(英文版) 2020年4期
Lan LUO Hua GUAN Siyu ZHOU Siyu ZHU Chuanle ZHAO Jianzhong TAN
Abstract [Objectives] This study was conducted to investigate the relationship between HSPs and the response of high temperature stress.
[Methods] Molecular biological techniques were applied to clone and analyze the gene sequence of DpHsp70 gene. The changes in the expression of DpHsp70 gene under high temperature stress and exogenous salicylic acid (SA) were observed and further analyzed by qRT PCR.
[Results] The coding region of the Dahlia DpHsp70 gene was 705 bp, encoding 234 amino acid residues (GenBank accession number: MH102288). Aligned with Compositae plants, the Dahlia DpHsp70 gene shared more than 83% homology in gene sequence while 99%-100% homology in amino acid sequence. Under the 35 ℃ high temperature stress, the expression of DpHsp70 gene in Dahlia petals significantly increased. Meanwhile, the expression of DpHsp70 gene further increased under SA at 35 ℃ temperature, which was significantly higher than those of the control group and the 35 ℃ high temperature stress treatment group. It was demonstrated that the Hsp70 gene in Compositae is with highly conservative property and its expression could be up-regulated in response to high temperature stress. It can also be concluded that applying exogenous SA can improve the high temperature resistance of Dahlia.
[Conclusions] This study provides a new experimental basis for elucidating the physiological function and mechanism of Dahlia in response to high temperature stress.
Key words Dahlia; High temperature stress; Salicylic acid (SA); Heat shock protein; Gene expression
Heat shock proteins are a class of proteins that are rapidly expressed by organisms stimulated by high temperature. According to the different molecular weights, heat shock proteins can be divided into five categories: HSP100, HSP90, HSP70, HSP60 and sHSP. At present, the research on HSP70 is the most extensive and in-depth. The HSP70 family has been found in the cytoplasm, chloroplast, endoplasmic reticulum and mitochondria of plant cells[1]. Under high temperature stress, HSP70 can bind with other heat shock proteins (HSP90, HSP60, HSP40) as molecular chaperone complexes, which participate in the processes of cell signal transduction[2] and protein folding[3], and can prevent the aggregation of denatured proteins, maintain the balance of cellular proteins, assist in the repair and depolymerization of denatured proteins[4], promote the degradation of oxidatively damaged proteins[5], improve the tolerance of cells to adversity stress, and thus maintain the normal physiological functions of cells and plants.
Dahlia (Dahlia pinnata Cav.) is a perennial bulb flower of the genus Dahlia. It has many varieties, various flower types and rich colors, and has high ornamental value. It is widely used in garden cultivation in China. Dahlia likes cool and half-shady environmental conditions, and when the growth temperature is higher than 30 ℃, it shows phenomena of growth stagnation, flower deformity or even death[6], which directly affects the application value of Dahlia in garden ornamentation. In this study, the RNA of petals of Dahlia at the full-bloom stage were used as material, the cDNA sequence of the coding region of Dahlia heat shock protein gene HSP70 was cloned and analyzed by the molecular biology experimental technology, and the expression changes of the Dahlia HSP70 gene under high temperature stress and exogenous salicylic acid (SA) treatment conditions were analyzed by the real-time fluorescence quantitative PCR technology, aiming to provide a new experimental basis for elucidating its physical function and mechanism in response to high temperature stress.
Materials and Methods
Plant materials
The single-petal type, yellow line Dahlia variety ‘Danbanhuang was selected as a test material. When the potted seedlings developed to the squaring stage, the plants with similar growth were placed into an artificial climate box for room temperature pre-cultivation. After the flower buds reached the translucent period, a partition test was carried out: spraying water on the leaf surface at 25 ℃ (marked as 25H), spraying water on the leaf surface at 35 ℃ (marked as 35H), and spraying 0.05 mol/L SA on the leaf surface at 35 ℃ (marked as 35S). After 1 week of treatment, the petals at the full-bloom stage were rinsed with double distilled water, frozen quickly with liquid nitrogen after air-drying, and stored in an ultra-low temperature refrigerator.
Extraction of total RNA and synthesis of the first strand of cDNA
The stored Dahlia petals of each test group were used to extract the total RNA of the petals according to the instructions of the RNA extraction kit (TIANGEN). The first strand of cDNA was synthesized using FastQuant RT Kit (with gDNase) reverse transcription kit.
Cloning of HSP70 gene
Primer premier 5.0 software was used to design primers (F: CAGCCTACTTCATGGACTC; R: CTAACCACTTCAA GATAGCG), and the specificity of primers was initially detected by NCBI Blast. The reverse-transcribed cDNA stock solution was diluted by 10 times and used as a template for PCR amplification. The reaction system (25 μl) included PCR mix 12.5 μl, ddH2O 10 μl, forward primer 1 μl, reverse primer 1 μl and template cDNA 0.5 μl, and the reaction parameters were set according to the primers. After the amplification, the product was recovered and entrusted to Shanghai Biotech Engineering Co., Ltd. for sequencing.
Standard preparation and qRT PCR
The mRNA of petals of Dahlia at full-bloom stage was obtained. The reverse transcribed cDNA obtained was used as a template and adjusted to 5 gradients as standards, to make standard curves of the internal reference gene and target gene. Based on the cloned DpHsp70 gene sequence, fluorescent quantitative PCR primers (F: TTCTCGACCGGCTCCATACA; R: TAGGCGGTTGAGGACATCGT) were designed; and β-actin gene was selected as the internal reference gene, and its primer sequences were F: ACGAAACACCTACAA and R: TTCTCGACCGGCTCCA TACA. Using Dahlia petal cDNA standard as a template, fluorescent quantitative PCR was performed on a StepOne Real-TimeTM PCR System using SYBR Green I reagent. The reaction system was 20 μl in volume. The PCR program was started with pre-denaturation at 95 ℃ for 30 s, followed by 40 cycles of 95 ℃ for 5 s and 60 ℃ for 30 s. Dissociation stage was added for subsequent analysis. All samples were repeatedly measured three times, and the data reading was completed by the fluorescence quantitative PCR instrument.
Results and Analysis
Cloning of cDNA sequence of Dahlia HSP70 gene
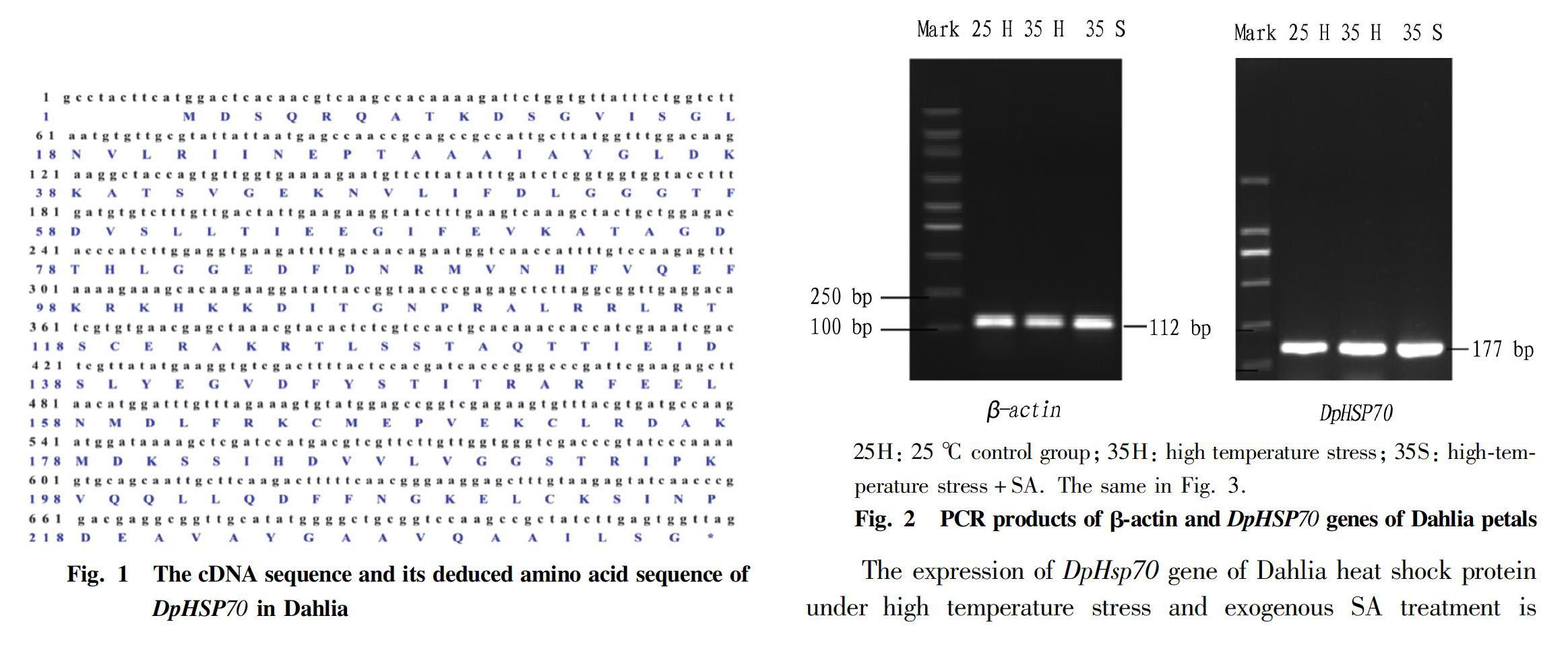
Using the cDNA after reverse transcription of the total RNA of Dahlia petals as a template, a band of about 700 bp was obtained after PCR amplification, which was similar to the expected fragment size. The amplified fragment was sequenced at 714 bp (Fig. 1), in which the Dahlia HSP70 gene coding region sequence was 705 bp, encoding 234 amino acid residues and a stop codon, designated DpHSP70 gene, and the gene accession number was MH102288.
Homologous analysis of DpHSP70 gene in Dahlia
In order to explore the relationship between the DpHSP70 gene of Dahlia and the HSP70 gene of other plants, Blast analysis was performed on the DpHSP70 gene sequence in NCBI, and 100 sequences showing homology in the range of 73%-85% were found (figure omitted). Specifically, the homology with Chrysanthemum×morifolium (AB503697), C. indicum (KJ561911) and Helianthus annuus (EF469896) in Compositae ranged from 83% to 85%, and the homology with Spinacia oleracea (AF034618) and Arabis alpine (LT669790) which do not belong to Compositae was more than 82%, indicating that the homology of HSP70 gene sequence in plants is high.
The deduced amino acid sequence of Dahlia DpHSP70 gene was subjected to pBlast analysis in NCBI. The results showed that the homology with the 100 sequences was in the range of 95%-100% (figure omitted). Specifically, the homology with H. annuus (XP022034520) in Compositae was highest, reaching 100%, and the homology with other plants in Compositae and non-Compositae plants was also higher than 95%, indicating that the amino acid sequence of HSP70 gene in higher plants has a high degree of conservation.
Analysis of DpHSP70 gene expression of dahlia under exogenous SA and high temperature stress
Using the cDNAs of the petals of Dahlia at the full-blood stage from three different treatments as templates (Fig. 2), amplified fragments of β-actin and DpHSP70 genes were obtained by PCR amplification. The fragment sizes were as expected, and only one specific band was detected in each amplified product. There was no primer dimer. It indicated that the designed primers were suitable for real-time quantitative PCR of SYBR Green. Secondly, according to the amplification curve obtained by fluorescence quantitative PCR (figure omitted), it was found that as the concentration of the template increased, the Ct value became smaller and smaller, showing a better linear relationship. The amplification efficiency of the β-actin and DpHSP70 genes were 107.34% and 114.24%, respectively. The relative expression level of DpHsp70 gene was calculated according to the formula RQ=2-△△Ct.
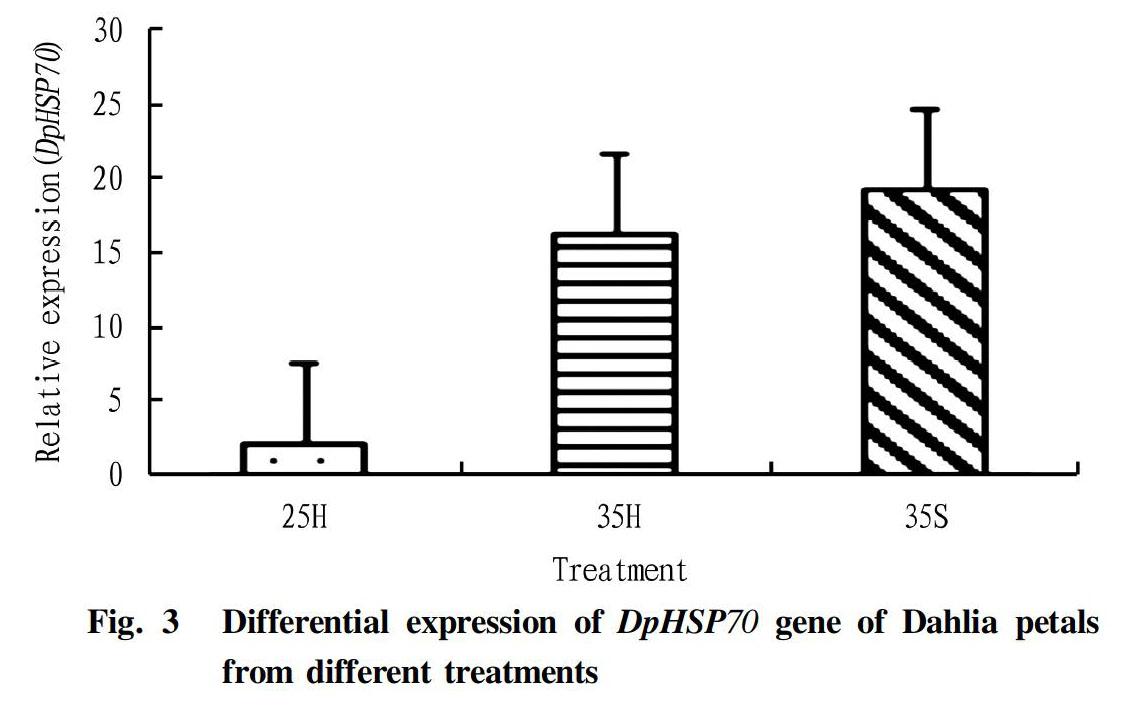
The expression of DpHsp70 gene of Dahlia heat shock protein under high temperature stress and exogenous SA treatment is shown in Fig. 3. Under the condition of high temperature stress (Fig. 3, 35H), the expression of DpHSP70 gene in Dahlia petals changed significantly, and the transcription level showed an up-regulation trend, which was 7.78 times that of the control (Fig. 3, 25H). It could be considered that the expression of DpHSP70 gene in Dahlia is related to the response to high temperature stress. Under the condition of external SA pretreatment plus high temperature stress (Fig. 3, 35S), the up-regulated expression of DpHSP70 gene in Dahlia petals was more significant, and its expression was also greater than that under high temperature stress (35H), proving that Dahlia DpHSP70 gene expression was induced by exogenous SA. It is speculated that this up-regulated expression could alleviate the damage caused by high temperature stress to Dahlia to a certain extent, and could improve its heat resistance.
Lan LUO et al. Cloning and Expression Analysis of Heat Shock Protein Gene DpHsp70 in Dahlia
Discussion and Conclusions
Plants are affected by various environmental factors during their growth and development, and accordingly protect themselves through a series of defense mechanisms. Under high temperature stress and other adverse conditions, the induced expression of heat shock proteins (HSPs) in plants is an effective defensive measure. At present, the genes of HSPs and transcription factor genes of various plants have been cloned and their role in resisting stress has been studied in various plants. In this study, on the basis of cloning Dahlia DpHSP70 gene, through homology comparison analysis, it was found that Dahlia DpHSP70 gene sequence shared the highest homology with chrysanthemum, and its deduced DpHSP70 gene amino acid sequence has the highest homology with sunflower, indicating that HSP70 gene is highly conservative in plants.
HSPs, as a class of stress response related proteins, are induced and expressed quickly and efficiently under stress, which is helpful for plants to alleviate stress damage. A large number of studies have shown that HSPs are positively correlated with plant heat resistance, and the expression level under high temperature stress is significantly higher than normal. For example, in Ageratina adenophora, the four genes HSP90, HSP70, HSP60 and HSP17 are induced by high temperature and are abundantly expressed in leaves and stems[7]. In this study, the expression of DpHsp70 gene in Dahlia petals under high temperature stress (35 ℃) significantly increased, indicating that Dahlia itself could induce the synthesis of heat shock proteins under high temperature stress to improve its tolerance to high temperatures.
As a plant defense hormone, SA can not only effectively control plant metabolic processes such as seed germination and stomatal closure, but also act as a system signaling molecule in the plant stress defense response network, regulating the production of various proteins, thereby enhancing resistance in plants against stress[10]. For example, in ornamental plants Dendrobium officinale[11], Phalaenopsis aphrodite[12], Lupinus micranthus[13] and Rosachinensis minima[14], external application of SA could increase the contents of soluble protein, soluble sugar and proline in plants under high temperature stress, and enhance heat tolerance in plants. It has also been found in Dahlia that the application of SA can improve the physiological and biochemical indicators of the response to high temperature stress, reduce the adverse effects on the growth of branches and leaves and photosynthesis, and help to reduce the damage of high temperature stress[15]. In this study, after spraying exogenous SA on the leaves of Dahlia, the expression level DpHsp70 gene in petal increased significantly under 35 ℃ high temperature stress. It could be considered that the DpHsp70 gene plays an important role in regulating the process of high temperature stress response, which is beneficial to improve the heat resistance of Dahlia. Moreover, the amino acid sequence of the heat shock protein Hsp70 is highly conserved in Compositae plants, which also shows that exogenous SA has certain value in open field cultivation and landscaping application of Compositae plants.
References
[1] RENNER T, WATERS ER. Comparative genomic analysis of the Hsp70s from five diverse photosynthetic eukaryotes[J]. Cell Stress & Chaperones, 2007, 12(2): 172-185.
[2] SUNG DY, VIERLING E, GUY CL. Comprehensive expression profile analysis of the Arabidopsis Hsp70 gene family[J]. Plant Physiology, 2001, 126(2): 789-800.
[3] TANIA M, MATTHIAS PM, STEFAN GDR. The Hsp70-Hsp90 chaperone cascade in protein folding[J]. Trends in Cell Biology, 2019, 29(2): 164-177.
[4] NILLEGODA NB, BERND B. Metazoan Hsp70-based protein disaggregases: emergence and mechanisms[J]. Frontiers in Molecular Biosciences, 2015(2): 36-47.
[5] REEG S, JUNG T, CASTRO JP, et al. The molecular chaperone Hsp70 promotes the proteolytic removal of oxidatively damaged proteins by the proteasome[J]. Free Radical Biology and Medicine, 2016(99): 153-166.
[6] LIU AC, WANG Q, PANG CM. Advances in horticultural study of dahlia in China[J]. Northern Horticulture, 2010, 34(11): 225-228. (in Chinese)
[7] GONG WN. Effect of heat shock protein gene of invasive plant Ageratina adenophora during low temperature stress[D]. Beijing: Chinese Academy of Agricultural Sciences, 2009. (in Chinese)
[8] PENG H, SONG WL, WANG XQ, et al. Effects of salicylic acid on plant stress resistance[J]. Horticulture & Seed, 2016(2): 70-74, 78. (in Chinese)
[9] YANG L, SHI S, WANG HJ, et al. Effects of salicylic acid on heat-resistance of Dendrobium officinale seedling under high temperature stress[J]. Acta Botanica Boreali-Occidentalia Sinica, 2013, 33(3): 534-540. (in Chinese)
[10] YANG HG, YAN SL, CHEN HJ, et al. Effect of exogenous methyl jasmonate, calcium and salicylic acid on the heat tolerance in Phalaenopsis seedlings under high temperature stress[J]. Chinese Agricultural Science Bulletin, 2011, 27(28): 150-157. (in Chinese)
[11] WANG XL, GAO Z, YU FX, et al. Physiological effect of exogenous salicylic acid on ornamental lupin under high-temperature stress[J]. Chinese Agricultural Science Bulletin, 2011, 27(25): 89-93. (in Chinese)
[12] QIN YZ, ZHOU ZL, LIU HT, et al. Effects of exogenous salicylic acid on physiological indexes of Rosachinensis minima under high temperature stress[J]. Journal of Southern Agriculture, 2018, 49(10): 2028- 2033. (in Chinese)
[13] ZHANG PP. Basic study on mechanism of thermotolerance and chemical regulation in Dahlia[D]. Suzhou: Soochow University, 2016. (in Chinese)

- 农业生物技术(英文版)的其它文章
- Investigation on Agronomic Characters of Dwarf Mutant 778 in Broomcorn Millet (Panicum miliaceum L.) and Analysis of Its Sensitivity to GA
- Construction of Technology System on Development and Repropagation in Vitro of Several Cultivars in Pear
- Construction of Camellia oleifera Cultivation Standardization System
- Nutrients Determination in Nuts from Different Torreya grandis Cultivars
- Photosynthetic Physiological Response to Drought Stress of Populus euphratica at Different Ages in Minqin
- Effects of UV-B Radiation on the Activity of Antioxidants in Flue-cured Tobacco Leaves