
红椿与毛红椿半同胞家系苗期生长变异与家系选择
2020-08-12 00:42肖兴翠张春花杨勇智郭洪英
湖南林业科技 2020年3期
肖兴翠,龚 详,张春花,杨勇智,刘 军,郭洪英
(1.四川省林业科学研究院,四川 成都 610081;2. 茂县综合林场,四川 阿坝 623200;3.攀枝花市农林科学研究院,四川 攀枝花 617061;4.中国林业科学研究院亚热带林业研究所,浙江 富阳 311400)
红椿Toonaciliatavar. ciliata和毛红椿Toonaciliatavar. pubescens都是楝科香椿属大乔木,毛红椿是红椿的变种,两个树种均生长迅速、树干通直、材质曙红,木纹美丽,都有“中国桃花心木”之称,是我国亚热带及热带地区速生珍贵用材树种,具有很高的经济价值和开发前景[1-2]。在国内,红椿主要分布在福建、湖南、广东、广西、四川和云南等省区,毛红椿主要分布在江西、湖北、湖南、广东、四川、贵州和云南等省区[3]。但由于红椿和毛红椿种子在天然环境中难于自然发芽成活,加上人为破坏,各地资源逐渐变少,被各地列为濒危保护树种[4-5],并先后被列为国家Ⅱ级保护濒危种[6]。随着国民经济和人民生活水平的提高,人们对高档木制家具的需求日益增大。国内学者从2000年左右开始进行遗传改良方面的研究,刘军等[7-8]对浙江、江西、安徽、贵州和云南等5省的毛红椿天然群体的遗传多样性及其取样方法[9]进行了研究,得出影响毛红椿种实大小的主要因素是无霜期和海拔[10]。红椿[11]和毛红椿[12]半同胞家系间存在较为丰富的变异,具选育潜力。陶丹等[13]对江西种源的15个毛红椿家系、李庆洪等[14]对全国9省14个红椿家系进行了苗期优良家系选择。红椿不同种源与家系的树高、地径生长均有极显著差异,可开展早期选择[15],刘军等从安徽南陵[16]及浙江开化[17]、黄红兰等[18]从江西、吴际友等[19-20]从湖南汨罗的毛红椿家系幼林中分别选择出了适宜造林点的优良家系,为红椿和毛红椿的良种选育打下了较好的基础。
“十二五”以来,红椿作为速生珍贵用材树种得到各方面的重视,四川省把红椿作为全省优先发展的6个珍贵用材树种之一,在川南地区、攀西地区优先发展。四川对于红椿育种方面的研究起步较晚,从2013年开始,四川省红椿育种团队先后在四川各地天然林中进行优树选择,并相继开展了红椿及其近缘种种质资源收集、评价、苗期选择及子代测定等方面的研究。2016年,本课题组对收集的红椿及毛红椿优树种子,以及亚林所和湖南省林业科学院提供的毛红椿种子,在四川省林业科学研究院成都市郫都区唐昌镇试验基地进行了育苗。通过对1年生红椿半同胞家系苗苗期生长性状进行测定和比较,旨在初步筛选出红椿苗期优良的半同胞家系,为红椿家系选育提供参考。
1 材料与方法
1.1 试验地概况
试验地位于成都市郫都区唐昌镇四川省林业科学研究院的试验基地,地理位置103°46′49″E,30°55′52″N。属亚热带季风性湿润气候,具有春早、夏长、秋雨、冬暖、无霜期长、雨量充沛、冬季多雾、日照偏少和四季分明的特点。年平均气温16 ℃,1月平均气温5 ℃,8月平均气温26 ℃左右。降水量979.4 mm,日照1 014.0 h。土壤为岷江新冲积灰色水稻土细沙粒泥层。
1.2 试验材料及来源
参试的40个红椿及毛红椿优树半同胞家系来源于7个省13个地区,其中8个四川种源的半同胞家系种子由本院红椿课题组采集,其余32个半同胞家系由亚林所和湖南省林业科学院提供,具体见表1。

表1 不同家系树种类别及来源Tab.1 Species categories and sources of different families编号家系树种类别家系来源编号家系树种类别家系来源1SC-119红椿四川德昌21YL-28毛红椿湖南张家界2SC-129红椿四川德昌22YL-31毛红椿湖南张家界3SC-205红椿四川米易23YL-33毛红椿江西九连山4SC-209红椿四川米易24YL-34毛红椿湖南张家界5SC-308红椿四川盐边25YL-35毛红椿浙江富阳6SC-361毛红椿四川盐边26YL-36毛红椿湖南张家界7SC-404毛红椿四川仁和27YL-37毛红椿江西九连山8SC-407红椿四川仁和28YL-38毛红椿浙江富阳9YL-1毛红椿江西官山29YL-40毛红椿江西九连山10YL-2毛红椿安徽泾县30YL-41毛红椿江西九连山11YL-3毛红椿江西官山31YL-42毛红椿江西九连山12YL-9毛红椿江西官山32YL-43毛红椿浙江富阳13YL-11毛红椿安徽泾县33YL-45毛红椿云南腾冲14YL-12毛红椿江西官山34YL-46毛红椿福建来舟15YL-13毛红椿江西官山35YL-47毛红椿福建来舟16YL-15毛红椿安徽泾县36YL-50毛红椿福建来舟17YL-16毛红椿江西官山37YL-51毛红椿福建来舟18YL-17毛红椿江西官山38YL-54毛红椿福建来舟19YL-18毛红椿浙江富阳39湖南2红椿湖南江华20YL-26毛红椿江西官山40湖南8红椿湖南石门
1.3 试验布置
2016年5月,对40个红椿及毛红椿优树半同胞家系种子进行分家系播种育苗。幼苗长到4~5片真叶时开始移栽到直径4.5 cm、高8 cm的无纺布轻基质段中,集中培养一个月后移到控根穴盘中,在大棚中进行培养,采用相同的方法进行苗期管理。
1.4 数据测量
2016年11月对不同家系1年生轻基质容器苗的苗高和地径进行测量,苗高用直尺测量,地径用游标卡尺测量,10株小区,3次重复。
1.5 相关计算公式
遗传变异系数:C=S/X×100%
(1)
式中:S为标准差;X为某一性状的群体平均值;
家系遗传力:h2=(MSv-MSt)/MSv×100%
(2)
式中:MSv为处理间均方;MSt为处理内均方[22];
遗传增益:△G=h2Si/X
(3)
式中:h2为性状的遗传力,Si为家系选择差,X为个体平均数[21-22];
现实增益:G=(Xi-X)/X×100%
(4)
式中:Xi表示个体某性状的大小,X为某一性状的群体平均值 。
1.6 数据统计分析
采用Excel 2003对数据进行整理,并采用SPSS 19.0软件对数据进行方差分析及多重比较。
2 结果与分析
由表2方差分析结果可知,40个红椿及毛红椿半同胞家系间的苗高和地径均有极显著差异(P<0.01),表明家系变异较大,可进一步开展优

表2 不同家系苗期生长方差分析Tab. 2 Analysis of variance of growth at seedling stage of different families项目变异来源平方和df均方F显著性遗传力家系间3 977.39639101.98542.1120.000**0.976苗高家系内193.740802.422总数4 171.135119家系间13.127390.3374.8090.000**0.792地径家系内5.600800.070总数18.727119 注:**表示差异极显著(P<0.01),*表示差异显著(P<0.05)。下同。
良家系选择[23]。苗高和地径的遗传力分别为0.976和0.792,遗传力均较大,苗高的遗传力大于地径,表明苗高受遗传的控制比地径更大。
2.1 不同家系苗高生长与变异
40个红椿及毛红椿半同胞家系的平均苗高为18.66 cm,苗高大小在10.20~35.50 cm之间,不同家系间差异显著(P<0.05)。苗高超过平均值的有17个家系,占家系总数的42.5%。苗高以33号家系最大,苗高为33.50 cm,显著高于其余39个家系,苗高变幅在30.00~35.50 cm之间,遗传增益和现实增益分别达77.69%和79.58%。其次是1号家系,苗高为28.72 cm,与33、7、6、3号4个家系间没有显著差异,显著高于其余35个家系,苗高变异区间在26.80~31.57 cm之间,遗传增益和现实增益分别达52.69%和53.97%。排在第3的家系是7号、6号和3号3个家系,苗高分别为28.31 cm、28.17 cm和27.50 cm,与33、7、6、3、2、12、8号7个家系间没有显著差异,显著高于其余32个家系,3个家系的遗传增益分别为50.54%、49.78%和46.29%,现实增益分别为51.77%、50.99%和47.41%。40个家系苗高的平均变异系数为6.70%,除了20号家系外,其余39个家系的变异系数均小于15%,表明苗木的苗高生长均匀一致,而20号家系的变异系数为18.07%,表明该家系的苗高存在丰富的遗传变异。详见表3。
2.2 不同家系地径生长与变异
由表4可见,40个红椿及毛红椿家系的平均地径为4.05 mm,地径大小在3.08~4.94 mm之间,超过平均地径的家系有18个,占家系总数的45%;不同家系间差异显著(P<0.05)。其中以33号家系的地径最大,为4.85 mm,与19、17、25、1、37、10号6个家系间没有显著差异,显著高于其余33个家系,地径变异区间在4.77~4.94mm,遗传增益和现实增益分别达15.66%和19.77%;其次是19号家系,地径为4.79 mm,与33、17、25、1、37、10、12号7个家系间没有显著差异,但显著高于其余32个家系,遗传增益和现实增益分别达14.55%和18.37%;排在第3的是17号家系,地径为4.68 mm,显著高于14个家系。40个家系的平均变异系数为5.84%,各家系的变异系数均小于15%,表明40个家系的地径生长比较均匀一致。

表3 不同家系苗高生长变异Tab.3 Growth variation of seedling height in different families编号均值±标准差/cm变幅变异系数/%遗传增益/%现实增益/%3333.50±3.04 a30.00~35.509.0877.6979.58128.72±2.52 b26.80~31.578.7652.6953.97728.31±0.43 bc28.00~28.801.5150.5451.77628.17±0.58 bc27.50~28.502.0549.7850.99327.50±0.70 bc26.80~28.202.5546.2947.41225.72±0.52 cd25.17~26.202.0236.9937.891225.57±1.70 cd23.60~26.606.6636.1737.05825.47±1.20 cd24.21~26.604.7135.6636.532824.60±2.65 d21.60~26.6010.7631.1131.87423.28±1.79 de21.65~25.207.7024.2224.812023.08±4.17 de18.57~26.8018.0723.1423.70520.80±1.06 ef20.00~22.005.0911.2211.502320.29±3.12 fg17.40~23.6015.398.548.751819.89±1.08 fgh18.71~20.835.446.486.641719.02±1.49 fghi17.40~20.337.831.911.963418.85±2.08 fghi17.17~21.1711.021.001.033218.73±1.21 fghi17.60~20.006.440.410.42918.45±1.97 fghi16.50~20.4310.65-1.06-1.083118.13±2.39 fghij15.40~19.8013.16-2.73-2.801418.03±0.90 fghij17.00~18.604.97-3.25-3.331517.70±0.98 ghijk16.60~18.505.56-5.00-5.123916.97±1.50 hijkl15.25~18.008.85-8.80-9.012916.91±2.07 ijkl14.86~19.0012.25-9.15-9.371016.76±0.81 ijklm15.86~17.434.84-9.90-10.142515.47±0.64 jklmn15.00~16.204.16-16.69-17.093015.26±0.31 jklmno15.00~15.602.03-17.78-18.223814.84±0.36 klmnop14.57~15.252.42-19.95-20.431914.33±1.89 lmnopq13.00~16.5013.21-22.62-23.17

续表3 不同家系苗高生长变异Continued Tab.3 Growth variation of seedling height in different families编号均值±标准差/cm变幅变异系数/%遗传增益/%现实增益/%4014.33±0.70 lmnopq13.60~15.004.90-22.62-23.172214.33±0.67 lmnopq13.67~15.004.64-22.62-23.172713.89±0.71 mnopq13.29~14.675.09-24.94-25.543613.40±0.53 nopqr12.80~13.803.95-27.50-28.171112.92±0.50 nopqr12.60~13.503.87-30.00-30.721612.42±0.23 opqr12.17~12.601.81-32.61-33.411312.19±0.69 pqr11.40~12.675.66-33.83-34.662112.15±0.59 pqr11.80~12.834.82-34.02-34.853512.06±0.59 pqr11.50~12.674.87-34.53-35.372411.85±0.56 pqr11.20~12.174.73-35.63-36.503711.61±1.14 qr10.33~12.509.79-36.87-37.762610.68±0.72 r10.20~11.506.71-41.75-42.77均值18.66±5.9210.20~35.506.70 注:表中小写字母为0.05水平上多重比较。下同。

表4 不同家系地径生长变异Tab.4 Growth variation of ground diameter in different families编号均值±标准差/mm变幅变异系数/%遗传增益/%现实增益/%334.85±0.09 a4.77~4.941.7615.6619.77194.79±0.09 ab4.72~4.891.8214.5518.37174.68±0.05 abc4.63~4.731.0812.2715.49254.55±0.27 abcd4.37~4.865.939.7912.3614.44±0.24 abcde4.18~4.665.467.649.65374.41±0.24 abcdef4.25~4.685.396.998.82104.40±0.02 abcdef4.38~4.420.456.868.66124.34±0.11 bcdefg4.24~4.452.445.627.10184.28±0.33 cdefgh3.90~4.497.654.455.6134.27±0.19 cdefgh4.08~4.464.454.325.45364.26±0.21 cdefghi4.02~4.384.824.065.12294.21±0.07 cdefghij4.16~4.291.623.214.05314.19±0.26 cdefghij4.01~4.496.242.753.47234.17±0.37 cdefghij3.85~4.578.832.302.90264.17±0.25 cdefghij3.89~4.386.032.302.90284.13±0.19 defghijk3.98~4.344.501.642.07154.11±0.10 defghijk4.00~4.192.441.251.58324.08±0.50 defghijk3.63~4.6112.160.530.68113.99±0.24 efghijkl3.77~4.256.10-1.23-1.55163.97±0.15 efghijklm3.85~4.143.86-1.62-2.04143.96±0.49 efghijklm3.45~4.4312.41-1.68-2.12383.95±0.37 efghijklm3.60~4.349.43-2.01-2.5443.94±0.27 efghijklm3.72~4.256.96-2.07-2.62203.93±0.08 efghijklm3.85~4.001.94-2.27-2.86403.92±0.37 efghijklm3.51~4.229.35-2.59-3.2893.90±0.10 fghijklm3.81~4.002.43-2.86-3.61

续表4 不同家系地径生长变异Continued Tab.4 Growth variation of ground diameter in different families编号均值±标准差/mm变幅变异系数/%遗传增益/%现实增益/%343.89±0.21 fghijklm3.74~4.135.40-3.12-3.93393.86±0.10 ghijklm3.79~3.972.50-3.70-4.6853.84±0.30 ghijklm3.50~4.047.76-4.03-5.0963.82±0.45 ghijklm3.35~4.2511.81-4.55-5.7573.81±0.29 ghijklm3.52~4.097.48-4.68-5.91243.79±0.16 hijklm3.64~3.954.11-5.14-6.4983.75±0.54 hijklm3.34~4.3614.43-5.92-7.47303.73±0.22 ijklm3.57~3.985.82-6.18-7.80213.71±0.21 jklm3.47~3.855.69-6.57-8.30223.69±0.21 jklm3.49~3.905.55-6.96-8.79273.62±0.13 klm3.54~3.773.52-8.33-10.5223.61±0.17 klm3.46~3.804.77-8.53-10.77353.51±0.33 lm3.18~3.849.39-10.48-13.24133.45±0.34 m3.08~3.759.89-11.66-14.72均值4.05±0.403.08~4.945.84
2.3 不同家系苗期选择
40个红椿及毛红椿半同胞家系苗高的遗传力大于地径的遗传力,因此以苗高为主、地径为辅进行苗期家系选择,将40个家系分为Ⅰ、Ⅱ、Ⅲ、Ⅳ 4类,遗传增益Ⅰ>Ⅱ>Ⅲ>Ⅳ,分为优、良、中、差4等。
由表5可见,Ⅰ类家系共5个,包括33、1、12、3、28号,入选率12.5%,苗高和地径选择差分别为9.32 cm和0.36 mm,平均苗高和地径分别为27.98 cm和4.41 mm,遗传增益分别为48.79%和6.98%,苗高和地径增益均较大,为优等家系,分别来源于云南、四川、江西和浙江四省;Ⅱ类家系包括23、18、17、32号4个家系,地径的选择差均为正值,平均苗高和地径分别为19.48 cm和4.30 mm,遗传增益分别为4.33%和4.89%,苗高和地径均有一定的增益,为良等家系,主要来源于江西和浙江;Ⅲ等家系包括7、6、2、8、4、20、5、34号8个家系,苗高选择差和遗传增益均为正值,但地径的选择差和遗传增益均为负值,苗高生长较好,但地径生长较差,为中等家系;Ⅳ等家系为除Ⅰ、Ⅱ、Ⅲ类之外的23个家系,其苗高和地径的选择差均为负值,长势较差,平均苗高和地径均比40个家系的平均值低,为差等家系。
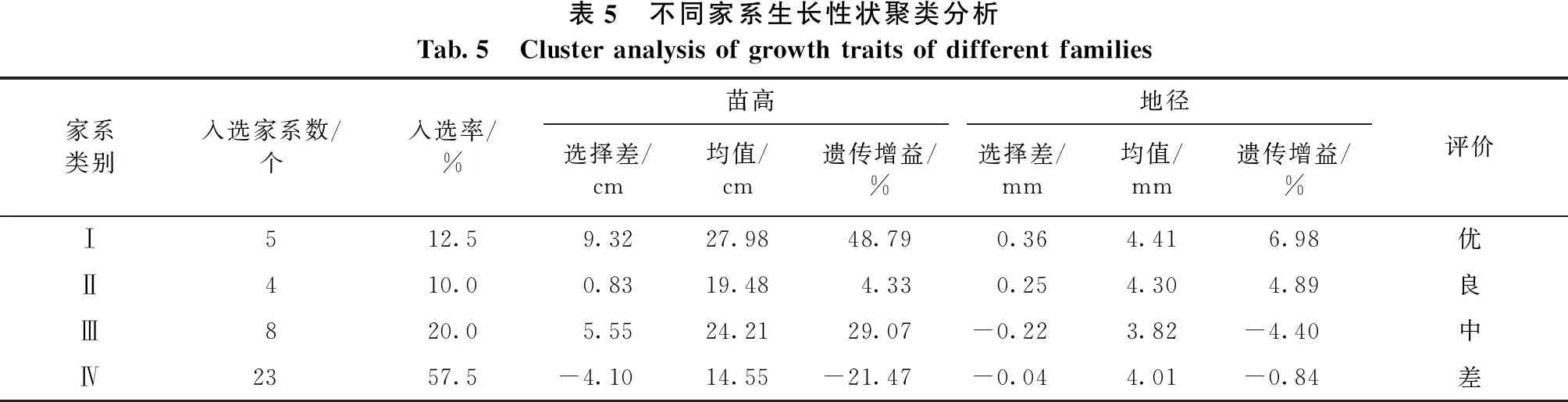
表5 不同家系生长性状聚类分析Tab.5 Cluster analysis of growth traits of different families 家系类别入选家系数/个入选率/%苗高地径选择差/cm均值/cm遗传增益/%选择差/mm均值/mm遗传增益/%评价Ⅰ512.59.3227.9848.790.364.416.98优Ⅱ410.00.8319.484.330.254.304.89良Ⅲ820.05.5524.2129.07-0.223.82-4.40中Ⅳ2357.5-4.10 14.55 -21.47-0.044.01-0.84差
3 结论与讨论
40个1年生红椿及毛红椿半同胞家系轻基质容器苗的平均苗高和地径分别为18.66 cm和4.05 mm,不同家系间均有极显著差异,超过苗高和地径平均值的家系个数均低于50%。
遗传力反映亲代将把性状遗传给子代的能力[23]。本文40个家系的苗高和地径的遗传力分别为0.976和0.792,表明苗高受到的遗传控制大于地径,苗高可作为苗期选择的主导因素。40个家系的遗传力低于李庆洪等[14]在汉源育苗的各省区红椿家系的遗传力(苗高0.993,地径0.992),但高于江西关山、马头山、九连山(苗高0.85,地径0.57)[13]和江西宜丰(苗高0.80,地径0.63)[12]毛红椿家系的苗期遗传力,表明这40个红椿及毛红椿家系间存在较为丰富的遗传变异,为以后红椿及毛红椿的良种选育和遗产改良提供了宝贵的基础资料。
40个半同胞家系的苗高大小在10.20~35.50cm之间,地径大小在3.08~4.94 mm之间,苗高和地径均以33号(YL-45)家系最大,分别为33.50 cm和4.85 mm,遗传增益分别为77.69%和15.66%,现实增益分别为79.58%和19.77%;其次是1号(SC-119)家系,苗高和地径分别为28.72 cm和4.44 mm,遗传增益分别为52.69%和7.64%。但40个家系苗高和地径的变异系数总体均小于15%,明显比李庆洪等[14]汉源育苗的14个家系低,家系内生长比较均匀一致,对今后选择优良家系造林有利。
为了提高选择效果,降低入选率、提高选择差是一种很好的方法[24]。本文以苗高为主,地径为辅,选择出33、1、12、3、28号5个优等家系,入选率12.5%,入选家系苗高和地径的选择差分别为9.32 cm和0.36 mm,遗传增益分别为48.79%和6.98%,选择差较大,则选择率较低,苗高和地径增益均较大,则选择效果较好。但苗期表现不能代表整个树体成长的过程,只能为红椿家系选育提供参考,最终还需要经过后期区域化造林试验,才能选育出适宜各区域推广的生长快、材性好、抗逆性强的优良家系。
猜你喜欢
中华实用诊断与治疗杂志(2022年1期)2022-08-31
中华实用诊断与治疗杂志(2022年1期)2022-08-31
养猪(2022年4期)2022-08-17
园艺与种苗(2021年8期)2021-09-23
农技服务(2020年1期)2020-12-17
山东农业科学(2019年11期)2019-12-24
现代农业科技(2019年1期)2019-07-11
森林工程(2018年1期)2018-05-14
现代农业科技(2017年24期)2018-01-22
湖北畜牧兽医(2015年11期)2016-01-11
