
花生四烯酸代谢基因对小鼠肝细胞癌中环氧化酶P450途径的影响
2020-08-10 13:41:00倪安妮梁清洋姚鸿飞李根亮唐玉莲
基础医学与临床 2020年8期
倪安妮,梁清洋,姚鸿飞,李根亮*,唐玉莲
(右江民族医学院 1.基础医学院 生化教研室; 2.临床医学院 骨科, 广西 百色 533000)
肝细胞癌(hepatocellular carcinoma,HCC)是世界上第6大最常见的癌,也是第3大癌死亡原因[1]。花生四烯酸(arachidonic acid,AA)代谢过程在肿瘤发生中起着关键作用[2]。环氧化酶P450途径(epoxygenase P450 pathway,EPP)是AA的一种代谢途径,环氧化酶P450蛋白及mRNA表达水平与癌细胞表达水平正相关[3]。环氧化酶P450主要在内质网膜或线粒体膜与特定的磷脂结合,这些膜由磷脂双分子层组成,可通过产生脂质微区影响环氧化酶P450的功能[4]。环氧化酶P450的活性中心是含有铁卟啉结构的血红素,活性中心的铁离子传递电子在环氧化酶P450参与生命活动的氧化还原反应中发挥重要作用[5]。因此内质网膜上氧化酶P450的铁离子结合(iron ion bonding,IIB)对EPP的作用、对AA代谢,及对癌的发生产生影响。然而,在HCC微环境中花生四烯酸(AA)代谢中内质网膜相关基因的功能还未见报道。本研究应用小鼠HCC模型,采用RNA-seq和生物信息学分析花生四烯酸(AA)代谢相关内质网膜基因在HCC微环境中的表达情况及参与AA代谢的差异表达内质网膜基因(AA metabolism differentially expressed endoplasmic reticulum genes,ADEGs)的功能,并用RT-qPCR验证内质ADEGs的表达情况,并通过HCC微环境中ADEGs位点多态性分析其与基因表达的可能相关性。研究结果对理解HCC的发生机制具有重要的理论意义。
1 材料与方法
1.1 材料
1.1.1 实验动物:洁净级6周雌雄各半昆明小鼠100只,体质量(30±5)g[长沙市天勤生物技术有限公司,许可证号:SCXK(湘)2019-0014]。
1.1.2 实验试剂和耗材:H22细胞系(小鼠肝癌细胞系)(南京凯基生物科技有限公司);RNA提取试剂盒(Invitrogen 公司);Cyp2b9、Cyp2c39、Gpx8、Gapdh引物(上海生工生物工程股份有限公司合成)。
1.2 方法
1.2.1 动物的分组及处理:细胞系H22在1640培养基常规培养48 h后通过细胞计数仪检测活细胞密度达到105~106个/10 μL,按细胞原液:0.9%氯化钠溶液=1∶1至1∶10稀释,放置室温无菌条件下待用。随机选取100只昆明小鼠,每只用配好的细胞悬液1×106~3×106个/0.1 mL在层流柜中腋下注射成瘤,饲养4周后,取肿瘤组织和癌旁组织。
1.2.2 RNA提取和测序:两组(n=40)样本组织各取50 μg,RNA质量检测、反转录、文库构建及测序数据的处理按常规程序进行。
1.2.3 花生四烯酸(AA)相关差异基因(DEGs)的筛选:通过DAVID 6.8在线软件搜索AA代谢信号通路中所有基因,与测序数据中的DEGs比较,筛选出两样本中内质网膜相关的AA代谢相关基因。
1.2.4 AA相关DEGs的功能富集分析:采用DAVID 6.8在线软件进行功能富集,分析其功能。
1.2.5 AA相关DEGs编码蛋白的互作网络分析:采用STRING 11.0在线软件对内ADEGs编码的蛋白进行蛋白共表达网络互作分析,分析其分布和功能相关性。
1.2.6 RT-qPCR验证AA相关DEGs的转录情况:随机选择了3个ADEGs(Cyp2b9、Cyp2c39、Gpx8),内参基因为GAPDH,进行RT-qPCR。两样本分别用RNA提取试剂盒提取总RNA,反转录试剂盒构建cDNA。qPCR检测用荧光定量PCR试剂盒,设置程序:三步法PCR程序,预变性95 ℃ 30 s;变性95 ℃ 10 s;退火60 ℃ 20 s;延伸72 ℃ 15 s;循环40次。溶解曲线反应程序,72 ℃~95 ℃,升温2.05 ℃/s,恒定温度95 ℃持续15 s;95 ℃~60 ℃,升温-1.71 ℃/s,恒定温度60 ℃持续60 s;60 ℃~95 ℃,升温0.05 ℃,恒定95 ℃持续15 s。用2-ΔΔCt计算结果,验证其表达量以及RNA-seq测序结果。
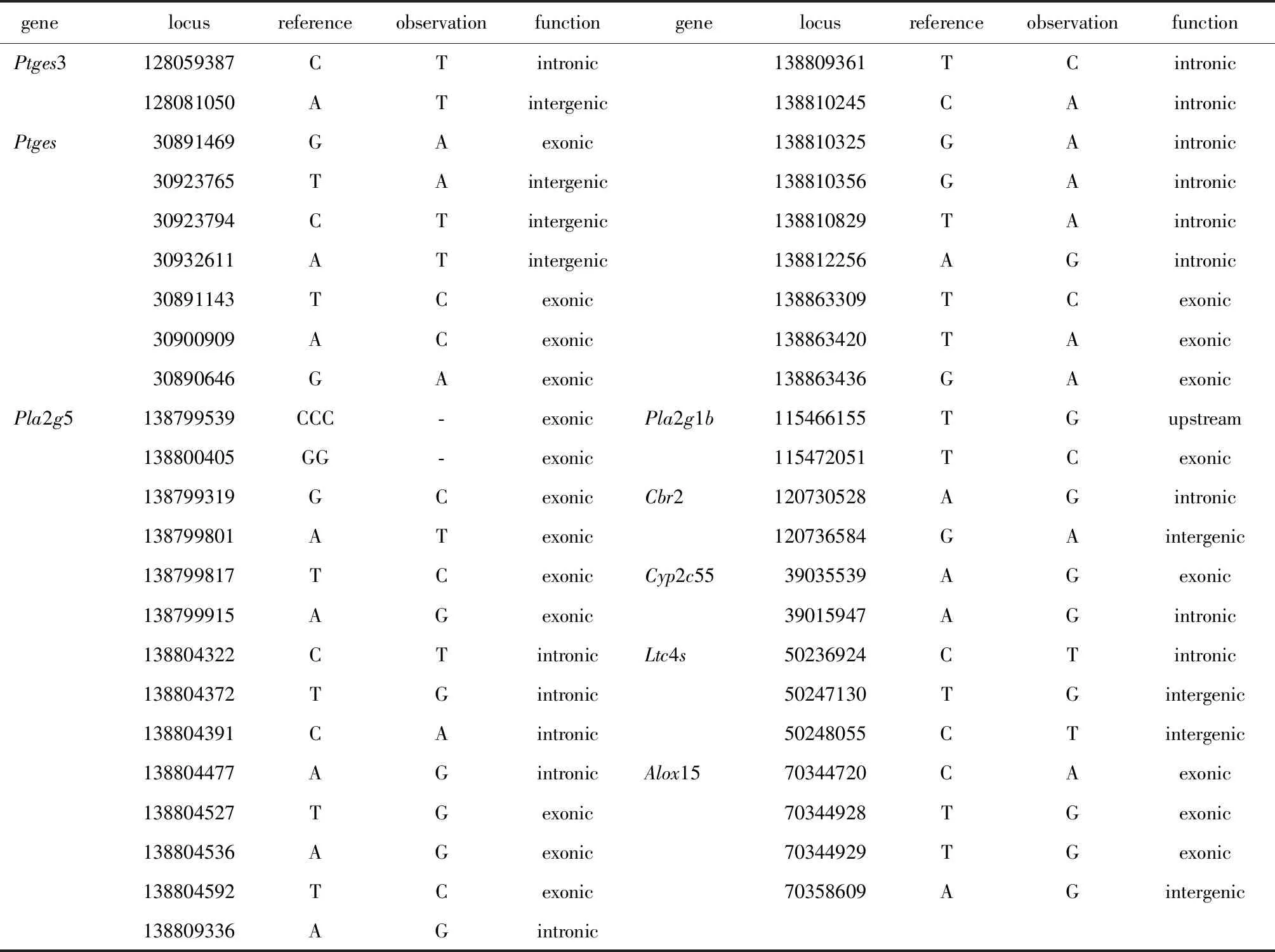
1.2.7 AA相关DEGs的AS(选择性剪切)和SNV(单核苷酸位点变异)/INDEL(插入和缺失)与基因表达的相关性分析:利用Excel软件对两样本中ADEGs中出现的频率倍数不小于2的AS和SNV/INDEL进行统计学相关分析,如果两样本间发生AS和SNV/INDEL频率倍数不小于2且0 基因表达以每百万测序碱基中每千个转录子测序碱基中所包含的测序片断数(expected number of fragments per kilobase of transcript sequence per millions base pairs sequenced,FPKM)比值≥2或≤0.5,且错误发现率(false discovery rate,FDR)≤0.05的基因为DEGs。使用Excel软件对ADEGs在两样本间发生频率数不小于2的AS和SNV/INDEL与DEGs进行统计学相关分析。 两组样本总RNA送公司测序,分别获得了5.01G base(35.28M clean reads)和5.02G base(35.32M clean reads)的数据。两样本所得clean reads数据与基因组的匹配率分别为93.65%和89.20%,且两样本测序数据的Q20皆大于 99%,测序数据质量较高。 DAVID 6.8搜索AA代谢信号通路相关基因89个,与测序数据比对,共获得了35个AA代谢相关DEGs,14个上调,21个下调(FPKM≥2或≤0.5, FDR≤0.05)。在肿瘤微环境到肝癌发生上调表达14个基因分别为Lta4h、Cyp4f18、Ltc4s、Gpx5、Gpx8、Cyp4f18、pla2g5、Cyp2j9、Cyp2c55、Alox15、Ptgs2、Ptg2s3、ptges、Cbr2;下调表达基因分别为Cpy2c29、Cpy4a10、Cpy4a12a、Cpy2c38、Cpy2c44、Cpy2c37、Cpy2c70、Cpy2c68、Cpy4f14、Cpy2b9、Cpy2j5、Cpy4a31、Cpy2e1、Cpy4a14、Cpy2c39、Cpy2c54、Cpy2c50、Cpy2j6、Ephx2、Pla2g12b、Cbr1经功能富集分析,共得到2个BPs(生物学程序),5个CCs(细胞组分),9个MFs(分子功能),1个US(蛋白位点序列特征)和6个KPs(信号通路)(图1)。 进一步对ADEGs富集结果分析发现,这35个DEGs中Cyp2c68、Cyp2c29、Cyp4f14、Cyp2b9、Cyp2j5、Cyp4a10、Cyp2c55、Cyp4a14等参与内质网膜中的IIB,Cyp2c29、Cyp2c68、Cyp2e1、Cyp2c44、Cyp2c39、Cyp2c55、Cyp4f18等参与EPP,其中Cyp2c29、Cyp2c38、Cyp2c55、Cyp2c39、Cyp2c54、Cyp2c37、Cyp2c70、Cyp2c50等一同参与内质网膜中IIB影响EPP。 ADEGs编码共表达蛋白的互作网络关系分析,发现11个DEGs编码蛋白相关性较强(图2)。 图2 ADEGs编码蛋白的互作网络关系Fig 2 Interaction network relationship of ADEGscoding proteins 随机选取3个ADEGs进行RT-qPCR,内参基因为Gapdh。数据结果表明,其结果与RNA-seq结果一致(图3)。引物序列见表1。 表1 RT-qPCR 引物序列Table 1 Primer sequence of RT-qPCR 对ADEGs的可变剪切事件(AS)、单核苷酸变异(SNV)/插入缺失标记(INDEL)分析,结果显示:均未发现AS。SNV和INDEL分析则显示,HCC肿瘤组织中共有21个DEGs发生了86个SNV和5个INDEL位点的改变。这些基因位点的改变主要发生在基因的4个部位,即外显子区(exonic)、内含子区(intronic)、基因间区(intergenic)和基因上游区(upstream),其中共有13个基因的SNV改变和1个基因的INDEL改变具有统计学意义(P<0.05)且13个基因的SNV改变和其中1个基因的INDEL改变与其基因的表达呈正相关(表2)。基因位点多态性分析结果还显示,有8个ADEGs在HCC肿瘤组织中出现43个SNV和2个INDEL,而癌旁组织中则没有出现这些SNV/INDEL(表3)。 BP.biological procedure; CC.cell component; MF.molecular function; KP.signaling pathways; US.protein site sequence characteristics 表2 SNV/INDEL与基因表达的相关性Table 2 Correlation between SNV/INDEL and gene expression 表3 HCC肿瘤组织中ADEGs的SNV/INDEL的发生情况Table 3 Occurrence of SNV/INDEL of ADEGs in HCC tumor tissues AA及其代谢产物参与各种组织的炎性反应和肿瘤的发生发展[6]。对HCC肿瘤及HCC微环境中AA代谢相关基因RNA-seq分析显示,其中有35个DEGs,富集分析DEGs显示基因功能集中在内质网膜中IIB对EPP的影响。在IIB的21个DEGs中有18个基因在HCC中下调表达,3个上调表达。EPP的14个DEGs中有12个基因在HCC中下调表达,2个上调表达。这些基因可与Cbr1、Alox15、Gpx8、Cbr2、Ptgs2一起参与氧化还原过程。这些基因中Cyp2c29、Cyp2b9、Cyp2c70、Cyp2j6等15个也属于内质网膜相关差异基因。以上基因中有22种即Cyp2b9、Cyp2c38、Cyp2c44、Cyp2c70、Cyp2c50、Cyp2j6等,还参与血红素结合,这些基因大部分都下调表达,表明从HCC的癌旁组织到肿瘤发生过程中,参与血红素结合的蛋白质组分可能表达量减少,又由于血红素是环氧化酶P450的活性中心,从而使其与正常的环氧化酶P450功能有差异。 GO分析结果还表明,这些基因中的17种:Cyp2c29、Cyp2b9、Cyp2c38、Cyp2c44、Cyp2c70、Cyp2c50、Cyp2j6等还参与花生四烯酸环氧酶活性。KP分析表明,AA代谢相关差异表达基因也参与以上 GO功能相关的6个信号通路,还参与GO相关的1个蛋白位点序列特征。这些结果表明,在 HCC中,大量AA相关的DEGs异常表达主要集中在EPP。而且,IIB的异常在改变EPP中发挥着极其重要的作用。蛋白互作分析也说明了这些基因功能的相关性。 SNV/INDEL结果表明,SNV/INDEL引起的基因多态性在 HCC中扮演着调控基因表达的重要功能。另外,研究结果还显示,HCC 中下调及上调表达在ADEGs的IIB相关DEGs中多具有SNV现象,这些基因突变后则会影响EPP功能。 综上所述,在HCC中共获得了35个ADEGs。功能主要集中在内质网膜中IIB对EPP的影响。在ADEGs中IIB相关的DEGs涉及的许多相关基因都有广泛的 SNV 现象,因此提示,IIB在 HCC 微环境的内质网膜中可通过自身的基因多态性突变引起EPP异常对AA代谢产生影响实现肿瘤的发生,在癌细胞的增殖过程中发挥重要作用,SNV 可能是一些基因上调及下调表达的重要调控因素。1.3 统计学分析
2 结果
2.1 RNA-seq测序
2.2 AA代谢相关DEGs筛选和功能富集分析
2.3 AA相关DEGs编码蛋白的互作网络分析

2.4 AA相关DEGs的RT-qPCR验证

2.5 AA相关DEGs的AS和SNV/INDEL与基因表达的相关性


3 讨论
猜你喜欢
解放军医学杂志(2021年12期)2022-01-18 03:53:24
现代临床医学(2021年1期)2021-01-26 00:55:52
昆明医科大学学报(2020年12期)2021-01-26 00:44:12
安徽医科大学学报(2016年12期)2017-01-15 14:21:55
中国粮油学报(2016年5期)2016-01-23 02:45:04
中国粮油学报(2016年5期)2016-01-23 02:44:50
中国粮油学报(2016年5期)2016-01-23 02:44:40
实用肝脏病杂志(2015年5期)2015-12-03 06:28:07
中国当代医药(2015年33期)2015-03-01 02:09:08
茶叶通讯(2014年2期)2014-02-27 07:55:39
