
外源激素对水稻籽粒碳氮代谢相关酶基因表达影响
2020-08-04 09:12金正勋王思宇张忠臣李钢夑朴钟泽
东北农业大学学报 2020年7期
金正勋,王思宇,王 珊,王 剑,张忠臣,李钢夑,朴钟泽
(1.东北农业大学农学院,哈尔滨 150030;2.韩国农村振兴厅农业科学院,全罗北道 全州 54874,3.上海市农业科学院作物育种栽培研究所,上海 201403)
淀粉和蛋白质是水稻胚乳主要储藏物质,也是碳氮代谢最终产物,其含量和分子结构等对稻米蒸煮食味品质影响较大[1]。籽粒淀粉合成积累代谢过程复杂,经一系列酶促反应,参与酶种类较多,其中ADPG焦磷酸化酶(AGPase)、颗粒结合型淀粉合成酶(GBSS)、可溶性淀粉合成酶(SSs)、淀粉分支酶(BEs)、异淀粉酶(ISA)是参与该酶促反应关键酶,酶活性强弱直接影响淀粉组分含量和精细结构等,各关键酶均有很多同工酶和相应同工型基因,OsGBSS1、OsAGPL2、OsSSSⅠ、OsSBEⅡb和OsISA1是在水稻胚乳中特异性表达且表达量较大的同工型基因之一,各种同工型基因表达均与淀粉组分含量和结构关系密切[2-3]。OsRSR1(Rice starch regulator 1)基因编码的蛋白是水稻众多转录因子中的一个,属于AP2/EREBP转录因子家族成员,参与淀粉合成调控,OsRSR1基因过表达使种子中淀粉合成基因下调表达,OsRSR1缺乏导致种子中淀粉合成基因表达量增加[4]。谷氨酰胺合成酶是调控蛋白质合成代谢关键酶之一,OsGS1;3基因是GS同工型基因之一,在籽粒中表达量较高[5]。
DNA转录是基因表达重要调控节点,其转录速度和量影响酶活性,调控性状表现。围绕基因转录调控国内外已有较多报道[6-7],但外源激素对水稻籽粒碳氮代谢相关酶基因转录表达的影响报道较少。鉴于外源激素和氮素营养对水稻生理特性、叶片衰老及活性氧代谢[8-9]、产量和品质等产生较大影响,是水稻生产上广泛使用的调控措施,且上述碳氮代谢相关酶基因转录表达量对水稻产量和品质性状影响较大,本试验选用不同类型粳稻品种比较分析灌浆成熟期籽粒碳氮代谢相关酶基因转录表达量对外源激素和氮素营养调控的响应变化,旨在阐明外源激素与基因表达关系和淀粉品质形成分子调控机理,为建立优质高产水稻栽培技术提供理论依据。
1 材料与方法
1.1 供试材料
本试验选用寒地稻作区具有代表性穗数型国标一级优质米中晚熟品种龙稻18号和穗重型晚熟超级稻品种松粳9号,供试的两个不同类型品种在哈尔滨地区均安全成熟。
1.2 试验方法及处理
试验于2018年和2019年在黑龙江省哈尔滨市香坊区东北农业大学农学院内开展盆栽试验,盆规格为长60 cm、宽40 cm、高40 cm,盆栽用土过筛混拌均匀后等量装盆。4月5日播种,大钵体盘育苗,每孔单粒点播催芽籽,大棚旱育秧管理。5月15日选取长势一致秧苗插秧,行距30 cm,穴距10 cm,每穴插3棵苗,每盆插2行8穴,每个处理插3盆,每个品种各处理3次重复,正常水管理。总施氮量为纯氮90 kg·hm-2,以盆表面积折算成每盆施氮量和磷钾肥施用量,N∶P2O5∶K2O比例为1∶0.5∶0.8。氮肥为尿素,磷肥为磷酸二铵,钾肥为硫酸钾。施肥方法为磷肥全作基肥施,钾肥50%作基肥,50%作穗肥施,氮肥50%作基肥,30%作分蘖肥,20%作穗肥施。插秧前3 d施入基肥与上层20 cm土壤混拌均匀,然后灌水泡土,插秧后15 d施分蘖肥,幼穗分化始期施穗肥。
试验设4个处理,分别为齐穗期至灌浆初期叶面喷外源激素6-苄氨基嘌呤(6-BA)和脱落酸(ABA)及齐穗期增施总施氮量20%氮肥(以下称为增施氮肥处理N),以喷施等量清水作空白对照。其中地上部叶片表面喷施50 mg·L-16-BA、10 mg·L-1ABA,每盆每次喷液量为150 mL,自齐穗期开始每隔5 d喷施1次,共喷3次。为使激素附着于植株上,喷施前加入1%吐温20。每次喷施6-BA和ABA溶液时,施氮处理和对照表面喷清水,以免水分干扰。抽穗前盆栽场搭建遮雨棚,以防降雨干扰,保证正常叶面喷施处理。
1.3 取样方法
抽穗时每个品种各处理选取长势一致且在同一天抽出叶鞘3 cm稻穗挂牌标记,自标记后第10、20、30天取标记穗,每个品种各处理每次取5个穗,迅速液氮处理后选取灌浆程度接近的穗中部籽粒于冰浴中去颖壳,放入冻存管密封,置于-80℃冰柜保存备用。
1.4 基因转录表达量测定
根据基因登陆号从NCBI上搜索并获取淀粉合成相关酶同工型基因OsGBSS1、OsSSSI、OsISA1、OsAGPL2、OsSBEⅡb、OsRSR1和谷氨酰胺合成酶同工型基因OsGS1;3以及Ubi5内参基因序列,然后通过BLAST确定各序列CDS区准确性。分别将各基因CDS区序列输入Primer Premier 5.0软件中,限定所需引物和产物长度等获得一系列引物,从中选取长度合适且无错配、发卡结构等引物,委托上海生工生物有限公司合成引物(见表1)。选用表1列出引物对灌浆不同时期各处理籽粒样品cDNA检测荧光定量PCR。本试验药品选用愚公生命科技有限公司提供的NOVA®TaqSYB®Green qPCR Premix,仪器使用Roche LightCyclerTM。参照Delte-DelteCt法分析基因相对转录表达量[10],①ΔCt=Ct(目的基因)-Ct(内参基因);②ΔΔCt=ΔCt(试验组)-ΔCt(对照组);③基因表达量=2-ΔΔCt。
基因上调率(%)=(各处理基因表达量-对照基因表达量)/对照基因表达量×100%。
基因衰减率(%)=(20 d基因表达量-30 d基因表达量)/20 d基因表达量×100%。
1.5 数据分析
2018年和2019年试验结果接近,本文主要根据2019年试验结果处理并分析数据,使用Excel 2007和SPSS 19.0处理数据并生成图表。

表1 RT-qPCR反应引物设计Table 1 Design of RT-qPCR reaction primer
2 结果与分析
2.1 外源激素对灌浆不同时期籽粒OsGBSS1基因转录表达量的影响
不同类型品种灌浆不同时期籽粒OsGBSS1基因转录表达量处理间多重比较结果列于表2。
供试的两个不同类型品种在灌浆不同时期,叶面喷施6-BA和ABA及增施氮肥处理的籽粒OsGBSS1基因转录表达量显著高于同时期喷水对照,其中抽穗后10和30 d叶面喷施6-BA和ABA处理的基因转录表达量又显著高于增施氮肥处理,而抽穗后20 d显著低于增施氮肥处理。从抽穗后20 d的基因转录表达量上调率比较可知,供试的穗重型超级稻品种松粳9号和穗数型优质品种龙稻18号均为增施氮肥处理的上调率最大,分别为133.22%和242.65%,其次6-BA分别为50.52%和154.36%,ABA上调率最小,分别为46.52%和123.85%,平均上调率优质品种龙稻18号为201.94%显著大于超级稻品种松粳9号的112.54%(见表2)。说明叶面处理外源激素6-BA和ABA及增施氮肥处理均显著上调灌浆过程中籽粒OsGBSS1基因转录表达量,基因转录表达量受外源因素调控作用较大,且基因转录表达量上调程度因品种类型及灌浆时期不同而有显著差异,优质品种基因转录表达量受外源调控因素影响大于超级稻品种。
在灌浆过程中供试的两个不同类型品种各处理籽粒OsGBSS1基因转录表达量均呈单峰曲线变化,随灌浆进程基因转录表达量逐渐上升,抽穗后20 d达峰值,此后开始下降。但下降速率和程度不同外源因素间差异较大。从抽穗后20~30 d基因转录表达量衰减率比较可知,松粳9号和龙稻18号两个不同类型品种均为增施氮肥处理的衰减率最大,分别为62.79%和71.76%,明显高于喷水对照的衰减率40.09%和30.51%,6-BA衰减率分别为13.46%和30.88%,ABA衰减率分别为22.43%和26.84%,均低于比喷水对照(见表2)。说明灌浆中后期籽粒OsGBSS1基因转录表达量下降速率和程度与外源调控因素间关系密切,不同外源调控因素对灌浆中后期籽粒OsGBSS1基因转录表达量调控作用机理差异明显。

表2 灌浆不同时期处理间OsGBSS1基因转录表达量比较Table 2 Comparition of mRNA transcription expression quantity of OsGBSS1 in rice sword between different fertility treatments during grain filling stage
2.2 外源激素对灌浆不同时期籽粒OsAGPL2和OsSSSⅠ基因转录表达量的影响
不同类型品种灌浆不同时期籽粒OsAGPL2和OsSSSⅠ基因转录表达量处理间多重比较结果列于表3。两个不同类型品种在灌浆不同时期,叶面喷施6-BA和ABA及增施氮肥处理的籽粒OsAGPL2和OsSSSⅠ基因转录表达量显著高于同时期喷水对照,不同处理两种基因转录表达量依次为6-BA>ABA>N>CK,各时期不同处理间差异达显著水平。从抽穗后20 d基因转录表达量上调率比较可知,松粳9号和龙稻18号均为6-BA处理上调率最大,其次是ABA处理,而增施氮肥处理上调率最小,籽粒OsAGPL2和OsSSSⅠ基因平均上调率超级稻品种松粳9号分别为87.36%和208.58%,显著大于优质品种龙稻18号的72.24%和64.87%(见表3)。说明叶面处理外源激素6-BA和ABA及增施氮肥处理均显著上调灌浆过程中籽粒OsAGPL2和OsSSSⅠ基因转录表达量,其中外源激素调控作用又显著优于增施氮肥处理,且基因转录表达量上调程度因品种类型及灌浆时期不同而差异较大,超级稻品种基因转录表达量受外源调控因素影响高于优质品种。
在灌浆过程中供试的两个不同类型品种各处理籽粒OsAGPL2和OsSSSⅠ基因转录表达量均呈单峰曲线变化,随灌浆进程基因转录表达量逐渐上升,抽穗后20 d达峰值,此后开始下降。但下降速率和程度不同外源因素间差异较大。从抽穗后20 d至30 d基因转录表达量衰减率比较可知,对于OsAGPL2基因转录表达量而言,松粳9号和龙稻18号均为6-BA衰减率最大,分别为61.06%和57.37%,其次是增施氮肥处理衰减率55.81%和ABA衰减率53.20%,再次是ABA衰减率53.19%和增施氮肥处理衰减率51.53%,喷水对照衰减率分别为41.48%和39.17%,各处理衰减率均高于喷水对照;对于OsSSSⅠ基因转录表达量而言,松粳9号为6-BA处理衰减率最大,为71.47%,而龙稻18号ABA处理衰减率最大,为56.42%,松粳9号ABA和增施氮肥处理间及龙稻18号6-BA和增施氮肥处理间衰减率差异均较小,但均高于喷水对照,其中松粳9号处理与对照间差异较大(见表3)。说明灌浆中后期籽粒OsAGPL2和OsSSSⅠ基因转录表达量下降速率和程度与外源调控因素间关系密切,不同外源调控因素对灌浆中后期籽粒OsAGPL2和OsSSSⅠ基因转录表达量调控作用机理差异明显。

表3 灌浆不同时期处理间OsAGPL2和OsSSSⅠ基因转录表达量比较Table 3 Comparition of mRNA transcription expression quantity of OsAGPL2 and OsSSSⅠin rice sword between different fertility treatments during grain filling stage
2.3 外源激素对灌浆不同时期籽粒OsSBEⅡb和OsISA1基因转录表达量的影响
不同类型品种灌浆不同时期籽粒OsSBEⅡb和OsISA1基因转录表达量处理间多重比较结果见表4。
两个不同类型品种在灌浆不同时期,叶面喷施6-BA和ABA及增施氮肥处理的籽粒OsSBEⅡb和OsISA1基因转录表达量均显著高于同时期喷水对照,且各时期不同处理间差异均达显著水平,灌浆不同时期OsSBEⅡb基因转录表达量依次是6-BA>ABA>N>CK,OsISA1基因抽穗后10 d顺序是ABA>6-BA>N>CK,抽穗后20 d顺序是N>6-BA>ABA>CK,抽穗后30 d顺序是6-BA>N>ABA>CK。从抽穗后20 d基因转录表达量上调率比较可知,OsSBEⅡb基因为松粳9号和龙稻18号均为6-BA处理的上调率最大,分别为498.69%和782.96%,其次是ABA处理,分别为266.48%和525.97%,而增施氮肥处理的上调率最小,分别为220.75%和364.20%;OsISA1基因为增施氮肥处理的上调率最大,分别为466.22%和284.79%,其次是6-BA处理,分别为194.43%和189.22%,ABA处理的上调率最小,分别为147.25%和148.03%。籽粒OsSBEⅡb和OsISA1基因平均上调率超级稻品种松粳9号分别为328.64%和269.30%,优质品种龙稻18号分别为557.71%和207.34%,不同类型品种和基因间上调率差异较大(见表4)。说明叶面处理外源激素6-BA和ABA及增施氮肥处理均显著上调灌浆过程中籽粒OsSBEⅡb和OsISA1基因转录表达量,其中外源激素对籽粒OsSBEⅡb基因上调表达作用显著大于增施氮肥处理,但OsISA1基因为增施氮肥处理的上调表达作用显著大于外源激素,基因转录表达量上调程度因品种类型和基因及灌浆时期不同而差异较大。
在灌浆过程中供试的两个不同类型品种各处理籽粒OsSBEⅡb和OsISA1基因转录表达量均呈单峰曲线变化,随灌浆进程基因转录表达量逐渐上升,抽穗后20 d达到峰值,之后开始下降。但下降速率和程度不同外源因素间差异较大。从抽穗后20 d至30 d基因转录表达量衰减率比较可知,对于OsSBEⅡb基因转录表达量而言,松粳9号和龙稻18号均为6-BA衰减率最大,分别为73.29%和80.56%,其次是松粳9号增施氮肥处理和龙稻18号ABA处理的衰减率,分别为67.10%和75.40%,而松粳9号ABA处理和龙稻18号增施氮肥处理的衰减率最小,分别为57.16%和74.92%,喷水对照衰减率分别为5.45%和1.26%,各处理衰减率均高于喷水对照;对于OsISA1基因而言,松粳9号和龙稻18号均为增施氮肥处理的衰减率最大,分别为62.46%和59.38%,其次是ABA处理,分别为50.00%和49.13%,而6-BA处理衰减率最小,分别为21.90%和37.34%,喷水对照衰减率分别为7.74%和3.08%,各处理衰减率均高于喷水对照(见表4)。说明灌浆中后期籽粒OsSBEⅡb和OsISA1基因转录表达量下降速率和程度与外源调控因素间关系密切,不同外源调控因素对灌浆中后期籽粒OsSBEⅡb和OsISA1基因转录表达量调控作用机理差异明显。
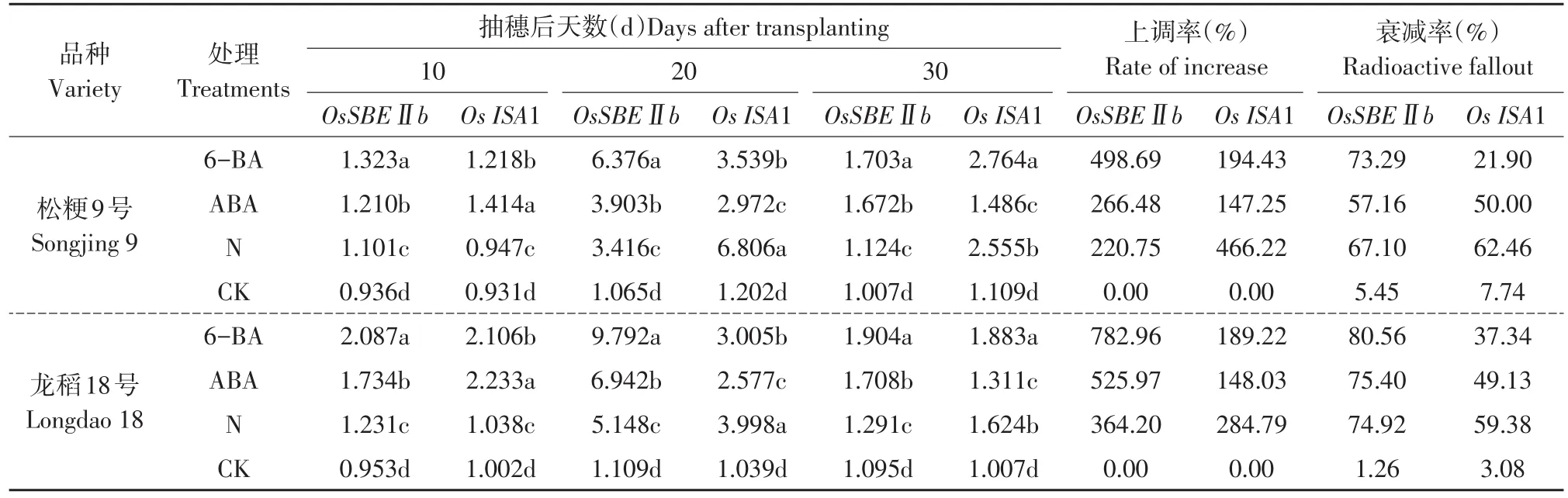
表4 灌浆不同时期处理间OsSBEⅡb和OsISA1基因转录表达量比较Table 4 Comparition of mRNA transcription expression quantity of OsSBEⅡb and OsISA1 in rice sword between different fertility treatments during grain filling stage
2.4 外源激素对灌浆不同时期籽粒OsRSR1基因转录表达量的影响
OsRSR1是调控淀粉合成关键酶基因转录表达的转录调控因子基因,不同类型品种灌浆不同时期籽粒OsRSR1基因转录表达量处理间多重比较结果见表5。
两个不同类型品种在灌浆不同时期,叶面喷施6-BA和ABA及增施氮肥处理的籽粒OsRSR1基因转录表达量均显著低于同时期喷水对照,灌浆不同时期OsRSR1基因转录表达量依次是CK>N>ABA>6-BA,且各时期不同处理间差异均达显著水平。从抽穗后20 d基因转录表达量下调率比较可知,松粳9号和穗数型优质品种龙稻18号均为6-BA处理的下调率最大,分别为27.93%和27.73%,其次是ABA处理,分别为11.00%和11.49%,增施氮肥处理的下调率最小,分别为3.86%和3.45%,平均下调率超级稻品种松粳9号为14.26%,优质品种龙稻18号为14.22%,两品种间几乎无差异(见表5)。说明叶面处理外源激素6-BA和ABA及增施氮肥处理显著下调灌浆过程中籽粒OsRSR1基因转录表达量,其受外源因素调控作用较大,且基因转录表达量下调程度不因品种类型发生较大变化。
在灌浆过程中供试的两个不同类型品种各处理籽粒OsRSR1基因转录表达量均随灌浆进程逐渐下降,抽穗后20 d达到最低谷后再逐渐上升,呈V字形变化动态,但上升速率和程度不同外源调控因素间差异较大。从抽穗后20d至30d基因转录表达量上升率比较可知,松粳9号和龙稻18号均为6-BA处理的上升率最大,分别为65.15%和72.37%,明显高于喷水对照的上升率44.87%和55.60%,增施氮肥处理的上升率分别为43.12%和46.28%,低于喷水对照,ABA处理的松粳9号为47.41%,高于喷水对照,龙稻18号为53.57%,低于喷水对照(见表5)。说明灌浆中后期籽粒OsRSR1基因转录表达量上升速率和程度与外源调控因素间关系密切,不同外源调控因素对灌浆中后期籽粒OsRSR1基因转录表达量调控作用机理差异明显。
2.5 外源激素对灌浆不同时期籽粒OsGS1;3基因转录表达量的影响
不同类型品种灌浆不同时期籽粒OsGS1;3基因转录表达量处理间多重比较结果见表6。
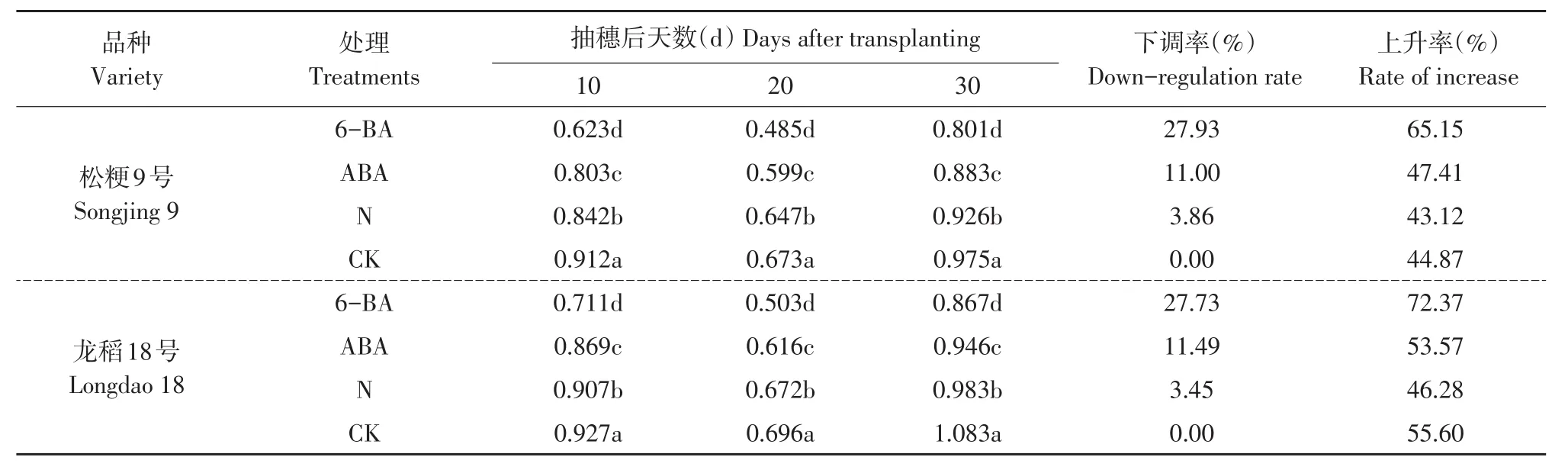
表5 灌浆不同时期处理间OsRSR1基因转录表达量比较Table 5 Comparition of mRNA transcription expression quantity of OsRSR1 in rice sword between different fertility treatments during grain filling stage
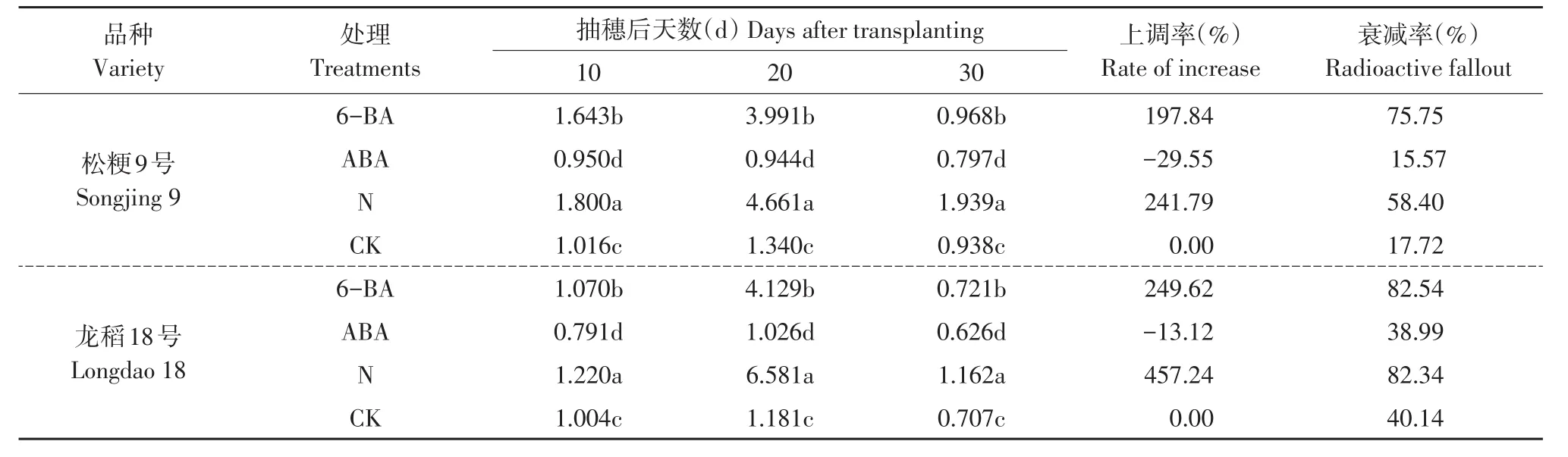
表6 灌浆不同时期处理间OsGS1;3基因转录表达量比较Table 6 Comparition of mRNA transcription expression quantity of OsGS1;3 in rice sword between different fertility treatments during grain filling stage
两个不同类型品种在灌浆不同时期,叶面喷施6-BA和增施氮肥处理的籽粒OsGS1;3基因转录表达量均显著高于同时期喷水对照,而ABA处理的转录表达量均显著低于同时期喷水对照,灌浆不同时期基因转录表达量依次是N>6-BA>CK>ABA,且各时期不同处理间差异均达显著水平。从抽穗后20 d基因转录表达量上调率和下调率比较可知,松粳9号和龙稻18号均为增施氮肥处理的上调率最大,分别为241.79%和457.24%,其次是6-BA处理的下调率分别为197.84%和249.62%,ABA处理的下调率分别为29.55%和13.12%,平均上调率龙稻18号为353.43%,显著高于超级稻品种松粳9号219.82%,品种间下调率差异并不大(见表6)。说明叶面处理外源激素6-BA和增施氮肥处理显著上调灌浆过程中籽粒OsGS1:3基因转录表达量,但ABA处理显著下调基因转录表达量,其受外源因素调控作用较大,且基因转录表达量上调或下调程度因品种类型及灌浆时期不同而差异显著,优质品种受外源调控因素影响高于超级稻品种。
在灌浆过程中供试的两个不同类型品种各处理籽粒OsGS1;3基因转录表达量均呈单峰曲线变化,随灌浆进程基因转录表达量逐渐上升,抽穗后20 d达到峰值,之后开始下降。但下降速率和程度不同外源因素间差异较大。从抽穗后20~30 d基因转录表达量衰减率比较可知,品种间和外源因素间衰减率差异较大,松粳9号6-BA和增施氮肥处理的衰减率分别为75.75%和58.40%,显著高于喷水对照的衰减率17.72%,ABA处理的衰减率为15.57%,低于喷水对照;优质品种龙稻18号6-BA和增施氮肥处理的衰减率分别为82.54%和82.34%,显著高于喷水对照的衰减率40.14%,ABA处理的衰减率为38.99%,低于喷水对照(见表6)。说明灌浆中后期籽粒OsGS1;3基因转录表达量下降速率和程度与外源调控因素间关系密切,不同外源调控因素对灌浆中后期籽粒OsGS1;3基因转录表达量的调控作用机理差异明显。
3 讨论与结论
基因转录为基因表达调控的重要环节之一,其表达量受遗传因素控制,也受氮素营养、温度、水分等外源环境因素影响。在OsRSR1突变体中与淀粉合成有关的OsAGPL1、OsAGPL2、OsAGPS2、OsGBSSI、OsSSSI等15个基因表达均被上调[4],其转录水平受干旱、高盐[11]抑制,进而调控下游逆境相关基因表达,以应对不良环境。研究表明,适当增加氮肥浓度处理对水稻OsGBSS1、OsISAs和OsGS基因mRNA表达量起正向调控作用[12-13]。沈直等研究表明,增温处理使籽粒中OsAGPase、OsSSS和OsSBE同工型酶基因表达水平上调,而OsGBSS同工型酶基因表达水平下调[14]。干旱胁迫下水稻胚乳中Wx基因表达量显著低于对照[15],卢红芳等则认为干旱提高TaGBSS1表达[16]。由本试验结果可知,喷施6-BA和ABA及齐穗期增施氮肥处理均显著上调灌浆过程中籽粒OsGBSS1、OsAGPL2、OsSSSⅠ、OsSBEⅡb和OsISA1基因转录表达量,显著下调转录因子基因OsRSR1转录表达量,且喷施外源激素上调或下调效果均强于增施氮肥;喷施6-BA和增施氮肥使OsGS1;3基因转录水平显著上调,而喷施ABA使OsGS1;3转录水平显著下调。表明灌浆过程中籽粒碳氮代谢相关酶基因转录表达量受外源激素影响较大,其影响程度甚至超过增施氮肥处理,且不同作用性质的激素对基因转录表达量影响并不一致,既有正向调控激素,也有负向调控激素,同时不同基因对外源激素调控响应也不同,既有上调表达基因,也有下调表达基因。本试验中喷施外源激素时基因上调或下调表达效果显著,但一段时间后基因表达量显著衰减,表明基因转录表达量与植株体内外源激素浓度关系密切,保持适当浓度激素是基因持续上调或下调表达的必需条件。鉴于基因转录表达量与酶活性线性关系密切,基因转录表达量高,酶活性强,否则酶活性低。酶活性直接影响性状表现程度,酶活性高性状表现强,反之表现弱。可知基因转录表达量调控是外源激素和氮素营养等对生物性状调控作用的分子基础,如何调控基因转录表达量有待深入研究。
围绕肥料、外源激素、温度等环境因素对稻米蒸煮食味品质的影响国内外从生理生化及分子水平上已开展大量研究,但基因转录表达调控层面研究报道较少。刘海英研究结果表明,喷施6-BA加速籽粒中淀粉组分积累,调节支/直淀粉比例进而改善籽粒淀粉品质[17]。外源ABA调节籽粒灌浆过程,影响产量和稻米品质[18]。金正勋等研究结果表明,齐穗期增施氮肥因显著提高稻米蛋白质含量而降低稻米蒸煮食味品质[19]。OsGBSS1、OsAGPL2、OsSSSⅠ、OsSBEⅡb、OsISA1和OsGS1;3基因在水稻籽粒中特异性表达,是参与籽粒淀粉和蛋白质合成的关键酶基因。颗粒结合淀粉合成酶、可溶性淀粉合成酶、ADPG焦磷酸化酶、淀粉分支酶、淀粉去分支酶、谷氨酰胺合成酶和转录因子OsRSR1参与籽粒淀粉和蛋白质合成代谢,并影响淀粉组分含量和精细结构等。水稻OsGBSSI表达受阻,导致胚乳中几乎不含直链淀粉[20],OsGBSS1基因mRNA表达量与直链淀粉含量同步变化[21];SBE酶催化支链淀粉精细结构,高温导致水稻胚乳中SBE酶活性显著下降,稻米支链淀粉长B链比例有所降低[22]。而OsSBEⅡb和OsISA1表达影响支链淀粉含量与精细结构,精细结构和淀粉糊化特性影响稻米品质,表现为支链淀粉中短链含量高,有利于糊化、米饭口感好[23];谷氨酰胺合成酶活性提高有利于水稻籽粒灌浆过程[24],OsGS1;3基因mRNA表达量与籽粒谷氨酰胺合成酶活性关系密切[25]。由本试验可知,淀粉及蛋白质合成相关酶基因转录表达受外源激素和氮素营养调控影响较大,而基因转录为基因表达调控重要环节,表达量直接影响酶活性,影响表观性状或碳氮化合物合成积累。本试验选择的碳氮代谢相关酶基因是众多酶基因中的一小部分,但却是参与淀粉和蛋白质合成代谢的关键酶基因,酶活性变化直接影响淀粉和蛋白质含量及分子结构,影响稻米蒸煮食味品质。因此,外源激素或氮素营养对稻米蒸煮食味品质的影响通过籽粒碳氮代谢相关酶基因的转录表达量调控实现。由于淀粉精细结构对稻米蒸煮食味品质影响非常大,深入研究外源激素与各种基因表达调控及淀粉精细结构间关系,对提高稻米蒸煮食味品质及建立环保型高效优质水稻栽培技术具有重要实践意义。
猜你喜欢
中国化肥信息(2022年8期)2022-11-30
中国化肥信息(2022年9期)2022-11-23
中国化肥信息(2022年9期)2022-11-23
热带作物学报(2022年7期)2022-08-06
舰船科学技术(2022年11期)2022-07-15
今日农业(2021年13期)2021-11-26
今日农业(2021年12期)2021-10-14
建材发展导向(2021年10期)2021-07-16
农村农业农民·A版(2017年7期)2017-07-17
园艺与种苗(2015年5期)2015-12-09
