
金沙江干热河谷新银合欢(Leucaena leucocephala)的光合与荧光特征干湿季对比研究
2020-07-31 08:21李登峰冯秋红胡宗达徐峥静茹
生态学报 2020年12期
李登峰,冯秋红,胡宗达,徐峥静茹
1 四川省林业科学研究院,四川卧龙森林生态系统定位站, 成都 610081 2 四川农业大学, 成都 611130
因深受亚热带季风及“焚风”效应影响,我国西南横断山区干热河谷的气候全年干旱炎热、干湿季分明,旱生的稀树草原及散生的肉质灌丛为该地区主要的植被类型[1]。由于干热河谷地区恶劣的自然环境,自然更新缓慢的植被特点,加之日益显著的人为干扰等因素的影响,导致该地区植被破坏严重,生态环境脆弱,生态恢复困难,干热河谷的生态屏障功效降低。因此,提升该地区生态环境的稳定性有利于保障中国西南地区乃至长江中下游地区的生态安全。目前,筛选出对干热河谷适应性强的树种是实现该地区生态恢复的主要措施之一[2- 3]。
新银合欢(Leucaenaleucocephala)是干热河谷地区的外来引进树种,其根系发达且具根瘤,固氮作用良好,生长迅速且耐旱耐瘠能力强,对干热河谷的环境具有较强的生态适应性,目前已成为该地区重要的植被恢复先锋树种[4]。叶片作为植物最主要的光合器官,其光合生理对环境变化十分敏感。在干旱胁迫下,叶片气孔开放程度降低、叶绿素含量减少、光合酶活性下降以及光合器官受损等原因都可能导致植物光合速率的下降[5],引起光合速率下降的原因可分为气孔限制及非气孔限制[6]。干热河谷的干湿季环境差异明显,植物光合生理特征的季节变化能够反映植物对该地区的适应性和抗逆性[7]。目前,对该地区新银合欢的光合生理方面研究包括气孔参数的坡位效应[8]、离体叶片的叶绿素荧光特征[9]等方面,但相关研究仍存在涉及的光合指标较为单一、研究对象多为盆栽幼苗、季相间变化的研究较少等问题。因此本文对干热河谷地区野生新银合欢在不同季节的光合生理特征及叶绿素荧光特征进行原位测定,分析和探讨干湿季间新银合欢光合机理变化的规律与原因,为其在干热河谷地区的推广奠定科学基础。
1 材料和方法
1.1 试验地概况
试验区域位于雅砻江和安宁河交汇处的攀枝花市米易县得石镇黑古田林场(21°44′04″N,101°51′08″E),海拔高度1048 m,土壤多为燥红土。该地属于典型的亚热带干热气候,全年气温较高,光照充足,年平均气温19.9℃,最冷月平均气温11.7℃,最热月平均气温25.2℃,极端最高气温41.2℃,极端最低气温-2.4℃,年平均日照总时数2397.4 h。但该地区气候干旱,干湿季分明,年平均降水量1101.2 mm,年平均蒸发量2178.5 mm,湿季为每年的5—9月,湿季降水量占全年的94%,干季为每年的10月至翌年4月。
1.2 试验材料与试验设计
在试验区域内,选择坡位(下坡)、坡向(南坡)、坡度(25°)等立地条件基本相同,长势基本一致,健康、无病虫害的新银合欢10株作为备测植株,并进行挂牌编号。选取枝条中上部的当年生、成熟、健康、向阳面叶片,分别于2017年7月上旬(湿季)和11月中旬(干季)进行相关指标的测定,每株设3—5个重复。
1.3 测定指标及方法
1.3.1 环境条件测量
在研究区域2 km范围内布设三台型号为NH121-NHSF48的小型“气象-土壤”监测仪(ZhongkeNenghui Tech.,China),监测离地1.2 m高度的气温(Air temperature,Tair)、气湿(Relative air humidity,RH)等气象环境指标,以及地下50 cm深度的土壤温度(Soil temperature,Tsoil)、土壤体积含水量(Soil volumetric water content,VWC)等土壤环境指标,该监测仪每30 min自动记录一次数据。
1.3.2气体交换参数和Pn-Ci响应曲线的测定
选择枝条上部、当年生、饱满、健康并位于向阳面的叶片,在晴朗天气的上午9:00—12:00和下午3:00—5:00两个时间段内,采用Licor- 6400便携式光合作用测定仪(Licor,USA)进行相关光合生理指标的测定。首先对叶室内的待测叶片进行活化和适应,将叶室内温度设定为25℃±1℃,湿度控制在75%±5%,利用CO2小钢瓶注入系统将环境CO2浓度(Ambient CO2concentration,Ca)设定为400 μmol/mol(模拟外界大气的CO2浓度),光合有效辐射(Photosynthetic active radiation,PAR)设定为1000 μmol m-2s-1,适应30 min。然后将PAR升至新银合欢的饱和光强1800 μmol m-2s-1(前期实验测得),适应10 min,测定此时的饱和光下净光合速率(Light-saturated net photosynthetic rate,Pnmax),以及气孔导度(Stomatal conductance,Gs)、蒸腾速率(Transpiration rate,Tr)、胞间CO2浓度(Intercellular CO2concentration,Ci)和叶片饱和水汽压差(Vapor pressure deficit,VPD)等相关气孔参数;并计算气孔限制值(Stomatal limitation,Ls),计算公式为Ls=(1-Ci/Ca)×100%,以及瞬时水分利用效率(Water use efficiency,WUE),计算公式为WUE=Pnmax/Tr。接着利用CO2小钢瓶注入系统将叶室内的Ca浓度梯度分别设置为:400、200、150、100、50、200、400、600、800、1000、1200、1400、1600、1800、2000 μmol/mol,其他条件不变,每个梯度浓度均在样本室与参比室达到平衡后适应3 min,待读数稳定后记录此时的净光合速率(Net photosynthetic rate,Pn)。最后利用直角双曲线的修正模型[10]对叶片的Pn-Ci响应曲线进行拟合,同时利用该模型计算出初始羧化效率(Initial carboxylation efficiency,CE)、光合能力(Photosynthetic capacity,Amax)、饱和胞间CO2浓度(Saturated intercellular CO2concentration,Cisat)、CO2补偿点(CO2compensation point,Γ)、光呼吸速率(Photorespiration Rate,Rp) 等光合生理参数;并利用基于Farquhar光合生化模型的光合助手[11- 12],求得最大羧化速率(Maximum carboxylation rate,Vcmax)、最大电子传递速率(Maximum electron transport rate,Jmax)、磷酸丙糖利用率(Triose phosphates utilization rate,TPU)、Jmax/Vcmax等光合生化参数。暗呼吸速率(Dark respiration rate,Rd)在夜间22:00—24:00用Licor- 6400测定,关闭叶室光源,Ca设定为400 μmol/mol,其他环境控制条件与白天一致。
1.3.3 叶绿素荧光参数的测定
选择与Pn-Ci响应曲线测定的相同叶片,先用锡箔纸完全包裹待测叶片24 h进行暗适应处理,然后将待测叶片迅速放入Licor- 6400荧光叶室(Licor,USA)中进行相关指标的测定,设置环境参数如下:温度设定为25℃±1℃,湿度控制在75%±5%,利用CO2小钢瓶注入系统将Ca设定为400 μmol/mol,PAR设定为0 μmol m-2s-1。叶片适应15 min后,待读数稳定,测定其PSII反应中心的最大光化学效率(Maximum photochemical efficiency of PSII,Fv/Fm)。接着打开光源,将PAR设定为1800 μmol m-2s-1,即饱和光强,活化30—45 min后,待读数稳定后计数,得到开放的PSII反应中心的激发能捕获效率(Excitation capture efficiency,Fv′/Fm′)、光化学猝灭系数(Photochemical fluorescence quenching,qP)、非光化学猝灭系数(Non-photochemical fluorescence quenching,NPQ)、PSII实际光化学效率(Actual photochemical efficiency of PSII,ФPSII)和电子传递效率(Electron transport rate,ETR)等叶绿素荧光参数。
1.3.4 叶绿素SPAD值测定
选择靠近主叶柄部、远离主叶柄部以及中间部位的叶片,利用SPAD- 502叶绿素含量测定仪(Konica Minolta,Japan)对待测叶片进行叶绿素SPAD值(Chlorophyll SPAD value,Chl-SPAD)测定,每个部位重复测定3次。
1.4 数据分析
利用Excel 2016进行数据整理与绘图,用SPSS 25.0对相关数据进行干湿季间的差异显著性分析及指标间的相关性分析。
2 研究结果
2.1 干湿季的环境条件比较
由表1可知,干湿两季间的气象和土壤环境条件均差异明显。干季的Tair、Tair(max)、Tair(min)、RH等气象环境指标较湿季均出现了不同程度的下降,其中Tair和Tair(min)降幅较大,分别为31.04%和60.84%。干季的Tsoil、Tsoil(max)、Tsoil(min)较湿季也分别下降了15.38%、14.66%、17.86%,干季土壤的VWC较湿季下降明显,降幅达到40.54%。

表1 干湿季的气象和土壤环境特征
2.2 干湿季新银合欢的气体交换参数特征比较
由表2可知,相较于湿季,新银合欢的干季Pnmax、Gs、Ci、Tr、WUE出现了显著上升,分别上升了26.50%、100.00%、20.35%、10.34%、15.63%(P<0.05),而Ls、VPD则分别显著下降了29.24%、65.91%(P<0.05)。

表2 干湿季新银合欢的气体交换参数
2.3 干湿季新银合欢的Pn-Ci响应曲线
由图1可知,湿季和干季新银合欢的Pn对Ci响应趋势基本一致。当Ci<300 μmol/mol,Pn随着Ci的上升迅速增加;随着Ci的继续上升,Pn的增速逐渐放缓;最后在较高Ci时,Pn的增速趋于平缓,但在本次试验梯度内未出现下降趋势。当Ci<150 μmol/mol,新银合欢干季Pn高于湿季;而Ci>150 μmol/mol,干季Pn则低于湿季。在相同Ca水平下,干季新银合欢的Ci高于湿季。直角双曲线修正模型对干湿季新银合欢Pn-Ci响应曲线有较高的拟合度,湿季和干季拟合模型的决定系数R2分别为0.9979和0.9991。
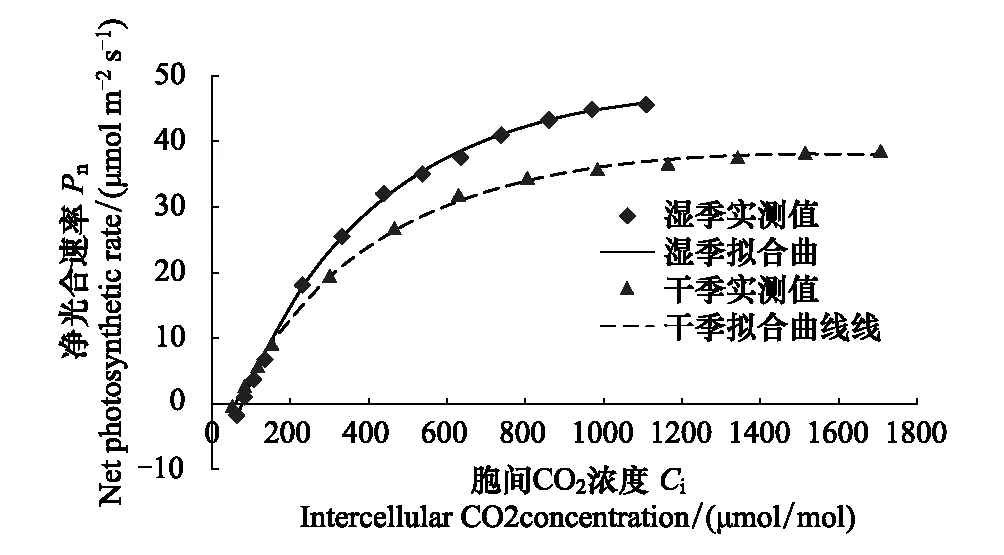
图1 干湿季新银合欢的Pn-Ci响应曲线 Fig.1 Pn-Ci response curves of Leucaena leucocephala in wet and dry season
2.4 干湿季新银合欢的光合生理生化特征及呼吸速率特征比较
通过对由直角双曲线修正模型和Farquhar光合生化模型计算出的相关光合生理生化参数的分析发现(表3、表4):新银合欢的干季CE、Amax、Γ、Vcmax、Jmax、TPU、Jmax/Vcmax较湿季均显著降低(P<0.05),下降幅度分别为28.57%、18.91%、22.89%、53.05%、62.36%、13.90%、19.51%;Cisat略有增加,但差异并不显著。呼吸速率方面,在干季,新银合欢的Rp较湿季显著下降了41.85%(P<0.05),但其Rd较湿季显著上升了96.55%(P<0.05)。
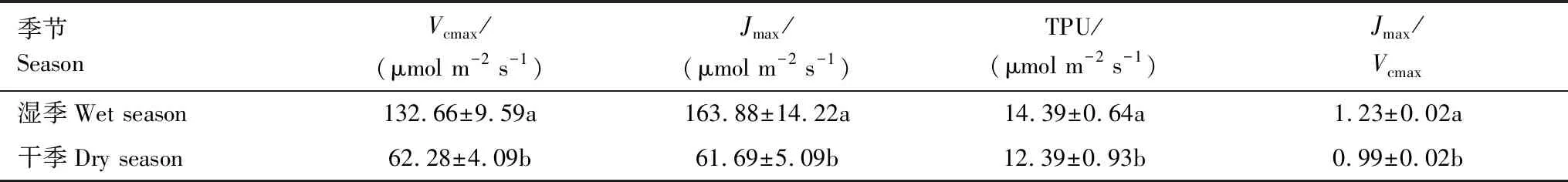
表4 干湿季新银合欢的光合生化参数

表3 干湿季新银合欢的光合生理参数及呼吸速率
2.5 干湿季新银合欢的叶绿素荧光参数及叶绿素含量特征比较
由表5可知,相较于湿季,新银合欢干季的ФPSII、qP、ETR均出现显著下降(P<0.05),下降幅度分别为33.33%、26.83%、30.89%;NPQ则出现显著上升(P<0.05),上升幅度为21.24%;Fv′/Fm′、Fv/Fm和Chl-SPAD变化并不显著,表明这三个指标对季节的变化并不敏感。

表5 干湿季新银合欢的叶绿素SPAD值及叶绿素荧光参数
2.6 新银合欢光合生理生化参数和叶绿素荧光参数的Pearson相关性分析
由表6可知,新银合欢的光合生理参数和光合生化参数有较好的相关性。Amax与CE、Γ、Rp、Vcmax、Jmax呈极显著正相关(P<0.01),与TPU、Jmax/Vcmax呈显著正相关(P<0.05);CE与Amax、Γ、Rp、Vcmax、Jmax、TPU、Jmax/Vcmax呈极显著正相关(P<0.01)。
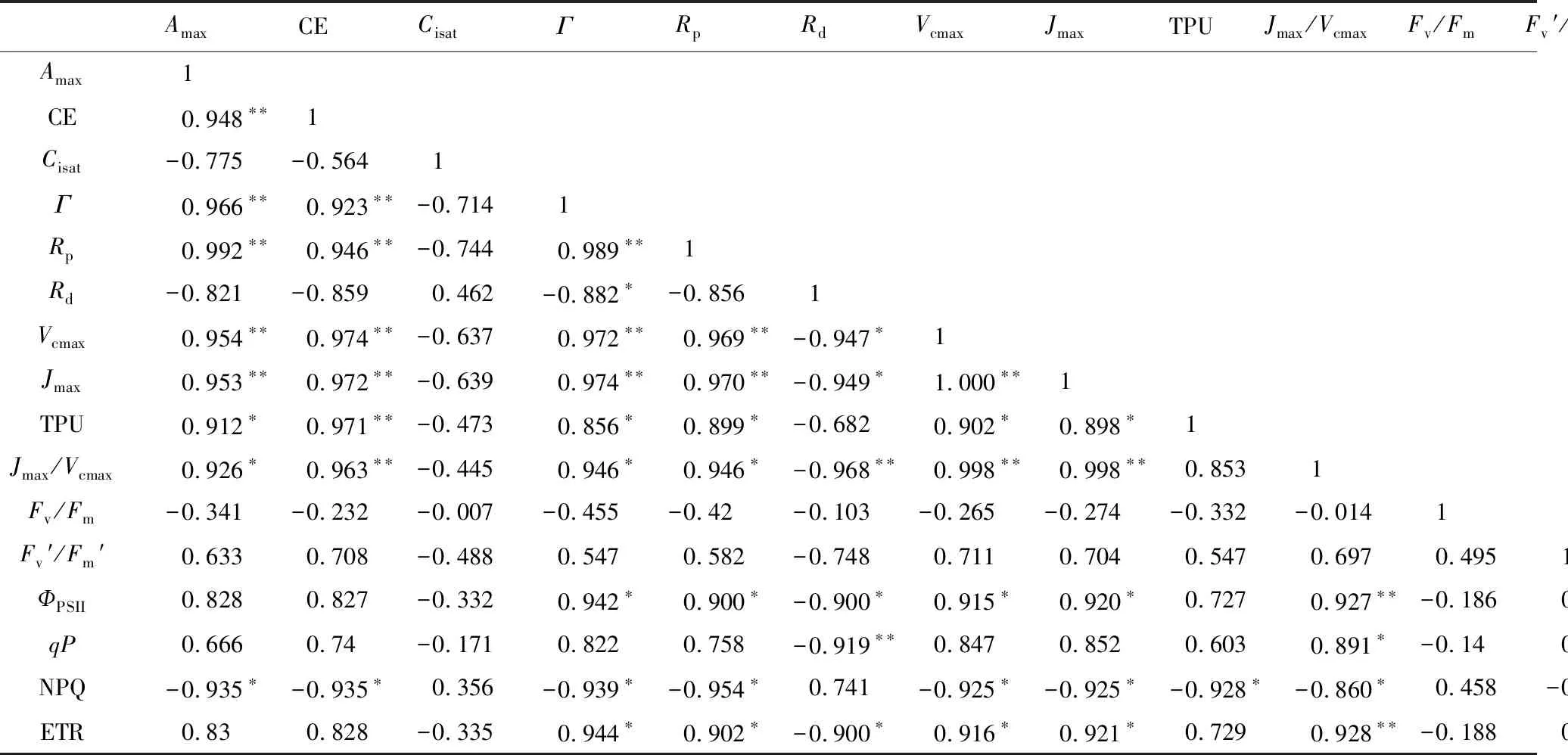
表6 光合生理生化参数和叶绿素荧光参数的Pearson相关性分析
Rp、Rd分别与光合生理生化参数呈相反的相关性。Rp与Amax、CE、Γ、Vcmax、Jmax呈极显著正相关(P<0.01),与TPU、Jmax/Vcmax呈显著正相关(P<0.05)。而Rd与Γ、Vcmax、Jmax为显著负相关(P<0.05),且与Jmax/Vcmax呈极显著负相关(P<0.01)。
叶绿素荧光指标中Fv′/Fm′、Fv/Fm两个指标与各光合生理生化指标相关性较弱,但NPQ与Amax、CE、Γ、Rp、Vcmax、Jmax、TPU、Jmax/Vcmax呈显著负相关(P<0.05)。ФPSII和ETR均与光合生理生化参数中的Γ、Rp、Vcmax、Jmax显著正相关(P<0.05),与Jmax/Vcmax极显著正相关(P<0.01),与Rd呈显著负相关(P<0.05)。
3 讨论
研究地区干季的气温较湿季下降迅速,极端高温环境有所缓解,日温差增加,气候环境由湿季的相对湿热转为相对干凉,土壤水分亏缺程度加深(表1)。
气孔是植物叶片与外界环境进行水气交换的重要通道,对环境变化极为敏感,在干旱胁迫下,通过降低气孔开放程度以减少水分散失是植物的一种常见干旱适应策略,但这种气孔调节策略将限制植物获取大气中CO2的能力,将可能导致光合速率下降[13]。但叶片的Gs不仅受土壤水分的影响,还与VPD等因素相关[14- 15]。VPD是反映大气水分条件的重要指标,也是驱动植物叶片蒸腾的主要动力[16]。韩路等[17]与王海珍等[18]的研究均发现,较高的VPD会引起灰胡杨(Populuspruinosa)的Gs降低。在湿季,新银合欢的VPD较高,其光合速率的气孔限制作用明显,原因可能为在干热河谷的干旱环境下,生长季植物需水旺盛,过高的VPD引起了叶片气孔阻力增加,以限制过度蒸腾,防止木质部栓塞和空穴化现象的产生[19],保证叶片内的水分平衡。而在干季,新银合欢的VPD大幅下降,Gs显著增加,Ci随之上升,Ls降低,植物可利用的CO2增加,叶片的Pnmax增加且增幅高于Tr的增幅,导致其WUE显著增加,表现出对干热河谷较强的适应性。相较于土壤水分的影响,新银合欢的气孔调节策略对VPD的变化更加敏感。
植物叶片的光合作用受外界环境和自身特性的影响,其Pn-Ci响应曲线及相关光合生理生化参数能较好地反映逆境条件下植物叶片的光合潜能、碳同化能力、光合电子传递效率、光合磷酸酶活性及光抑制水平高低等特性。本研究中,新银合欢的湿季和干季Pn-Ci响应曲线都比较符合经典形式(图1)[20],当Ci<150 μmol/mol,新银合欢的湿季Pn低于干季,但湿季的CE显著高于干季,表明湿季较高的Rp和Γ是导致该阶段Pn值较低的主要原因。CO2和O2作为Rubisco不同底物,在同一活性点位与其发生竞争性结合,产生竞争性的羧化反应和氧化反应。在饱和光强下,光下暗呼吸速率一般较低,此时的呼吸作用以光呼吸(Rp)为主;Γ是植物叶片光合作用和呼吸作用两个过程彼此平衡时的Ci浓度,代表Pn-Ci曲线与坐标系横轴的交点[21]。由于湿季新银合欢的气孔限制作用明显,在同等Ca水平下,其Ci低于干季,可用于羧化反应的CO2底物较少,同时湿季的高温(表1)也会调节Rubisco的动力学常数,提高氧合速率[22]。最终导致湿季Rp增加,Γ升高,Pn-Ci曲线与坐标系横轴的交点右移,使得在低Ci水平时(Ci<150 μmol/mol),湿季的Pn相对较低。但光呼吸并不是一个无意义的消耗过程,它能在胁迫环境下保护光合器官[23]。相关性分析表明,新银合欢的Rp与多个代表光合器官活性的光合生理指标呈显著正相关(表6),说明湿季较高的光呼吸对光合器官的保护作用显著,这与李环和吴沿友[24]的研究结果一致。在叶片气孔限制程度较高的湿季,新银合欢可能通过较高的光呼吸以甘油酸的形式回收部分由于RuBP氧化造成的碳损失[25],同时对过剩的光能进行快速耗散,减轻湿季高温环境对光合器官的损害[26]。因为光呼吸占新银合欢总光合速率的比例较高,在干季新银合欢的Rp显著降低可能是引起其Pnmax增加的另一个重要原因。而随着Ci的升高,Pn逐渐增加,羧化反应逐渐替代氧化反应成为主导反应[27]。Jmax/Vcmax值代表了Rubisco活性和电子传递速率的功能平衡,以及N素在光合系统内部间的分配[28- 29]。在本研究中新银合欢干季的CE、Amax、Vcmax、Jmax、TPU以及Jmax/Vcmax较湿季均出现显著下降(P<0.05);在Ci>150 μmol/mol阶段,干季的Pn值也明显低于湿季(图1)。表明在干季,叶片的光合酶活性和数量降低,羧化效率、光合电子的传递及RuBP再生都受到了极大的影响,分配给羧化反应的N素也出现下降,叶片的光合器官活性降低,CO2利用率下降,光合速率受此影响显著,最终导致Amax降低,这与王海珍等[30]的研究结果一致。同时,新银合欢叶片的干季Rd较湿季显著增加,也从另一个角度说明,新银合欢在干季通过增加消耗来促进其抗逆性,进而维持生息。
植物叶片的叶绿素荧光参数与光合作用密切相关,能很好地揭示光合进程的“内在性”[31]。Fv/Fm反映PSII反应中心利用光能的最大能力,在无胁迫条件下一般在0.8以上[32- 33],新银合欢在干湿季的Fv/Fm均低于0.8,说明其光合器官活性在干湿两季都受到一定程度的抑制。ФPSII反映在PSII反应中心部分关闭的情况下的实际光化学效率,与植物叶片碳同化能力和光呼吸相关[34],ETR则代表实际光强条件下的表观电子传递效率,qP为光化学猝灭系数,表示PSII反应中心通过将天线色素捕获的光能作用于光化学反应的比例[35- 36]。相较于湿季,新银合欢干季的ФPSII、qP、ETR出现了显著下降,相关研究认为,qP的降低说明其PSⅡ反应中心开放程度下降,PSⅡ的原初电子受体QA重新氧化的能力降低,PSⅡ的电子传递活性减弱,致使还原态QA携带的电子不能快速向后传递,阻碍了整个光合电子传递过程,从而导致ФPSII的下降,光合器官活性降低[37- 38],这与李泽等[39]和黄纯倩等[40]的研究结果类似。在干季,新银合欢在光化学效率降低、光合电子传递受阻的情况下,过剩的光能如不能及时耗散,将对光合系统造成破坏[41]。NPQ代表非光化学猝灭系数,反映PSⅡ天线色素吸收的光能不能用于光合电子传递而以热能形式耗散掉的光能部分,是光保护机制中热耗散作用的体现[42]。新银合欢在干季通过增加NPQ,加强热耗散,以保护光合器官,这与魏清江等[43]的研究结果一致。但有学者认为植物叶片在胁迫条件下qP的下降和NPQ的上升是光合机构被破坏和能量耗散过程加强运转的共同结果,这是一种自我消耗性的,被动的适应方式,并不利于植物的长期生长[44- 45]。通过对叶绿素荧光参数和光合生理生化参数间的相关性分析发现,新银合欢的ФPSII、NPQ、ETR三个指标均与Vcmax、Jmax、Jmax/Vcmax等光合生理指标呈显著性相关(P<0.05),说明这三个荧光参数对干热河谷地区新银合欢光合生理特征具有较好的指示作用。
4 结论
综上所述,在干热河谷相对湿热的湿季,新银合欢的气孔开放程度较低,光呼吸对光合器官保护作用显著,光合器官活性较高。而在相对干凉的干季,土壤水分亏缺程度增加,新银合欢叶片的光合器官活性降低,光抑制程度加深,光合能力下降,热耗散的光保护作用得到增强;气孔导度的增加和光呼吸的下降对其光合进程具有一定的补偿作用,表现出对干热河谷较强的适应性。影响干季光合速率的主要限制因素由湿季的气孔限制转变为非气孔限制。叶绿素荧光参数中的ФPSII、NPQ、ETR三个指标能够较好地反映出干热河谷地区新银合欢叶片的光合能力的季节变化。
猜你喜欢
干旱地区农业研究(2022年1期)2022-01-28
阅读(科学探秘)(2020年8期)2020-11-06
装备制造技术(2019年12期)2019-12-25
音乐研究(2019年5期)2019-11-22
湖南饲料(2019年4期)2019-10-17
中国生物医学工程学报(2019年5期)2019-07-16
科技创新与品牌(2019年12期)2019-02-06
中国果业信息(2019年1期)2019-01-05
生物学教学(2017年9期)2017-08-20
Coco薇(2017年5期)2017-06-05
