
不同类型高寒草地群落结构与生产对施氮的响应及其敏感性
2020-07-31 08:20石培礼
生态学报 2020年12期
宗 宁, 石培礼
1 中国科学院地理科学与资源研究所生态系统网络观测与模拟重点实验室, 北京 100101 2 中国科学院大学资源与环境学院, 北京 100049
氮素是限制陆地生态系统生产最重要的元素[1]。近几十年来,工业发展和农业施肥导致的氮输入增加在一定程度上缓解了陆地生态系统的氮限制,也对生态系统的功能产生重要影响[2- 4]。虽然氮输入增加对不同生态系统的影响存在较大差别,一般来说氮输入增加会促进生产力提高,故长久以来施肥被作为一种改良退化草地养分状况的重要管理措施[5- 6]。施氮能显著改善草地植物群落结构,有助于草地生产力的恢复,改善土壤表层土壤肥力。但由于氮输入增加会促进一些机会主义物种的生长[7- 8],引起土壤理化性质的改变,因此会降低植物物种丰富度,这种物种丰富度的降低反过来会降低群落生产和稳定性[9- 12]。
但目前尚不清楚的是氮输入增加对群落组成和功能的改变是依赖于施氮剂量还是群落原有组成。已有研究表明,对于养分贫瘠的草地,氮输入增加会缓解生态系统氮限制,引起生态系统物种多样性的增加[11]。与之相反,对于成熟稳定的同类草地,氮输入增加会打破生态系统内部的物种共存,导致物种丰富度的降低[11]。量化不同类型草地对外源氮输入增加的响应显的非常有必要。但目前研究广泛认可的结论是生态系统对氮输入增加存在饱和现象[11,13- 16],即低氮输入会提高草地生产,而长期高氮输入不仅不会持续增加草地生产,还可能会引起生产力降低,转折点的氮输入剂量被称为饱和阈值。已有研究表明,内蒙古温带草地对氮输入的饱和阈值为105 kg N hm-2a-1[11],半干旱草地的氮饱和阈值为91.7 kg N hm-2a-1[15],而美国落基山地区高寒草地的氮饱和阈值为46 kg N hm-2a-1[13- 14],与青藏高原的其他研究结果接近(50 kg N hm-2a-1)[16]。目前关于羌塘高原不同类型高寒生态系统对氮输入响应的研究仍然十分缺乏。
青藏高原作为世界上海拔最高的地区,对气候变化十分敏感[17]。羌塘高原地处西藏高原腹地,是西藏高原的主体部分之一,平均海拔在4500 m以上。由于海拔高、气温低,植物生存环境条件比较严酷,各生态因子常常处于植物生存的阈值边缘,植物生长受到养分有效性的限制[18]。高寒草地占据了青藏高原面积的60%以上,是维持高原地区生产力和畜牧业发展的基础。羌塘高原自东向西水分逐渐降低,植被也呈现地带性分布特征。东部是以高山嵩草(Kobresiapygmaea)为优势种的高寒草甸,向西过渡到以紫花针茅(Stipapurpurea)和线叶苔草(Kobresiaduthiei)为优势种的高寒草甸草原[19]。高山嵩草作为地带性植被,是成熟稳定的高寒植被群落;而高寒草甸草原地处高寒草甸向典型高寒草原的过渡地带,植物种类兼具草甸和草原的特征,因此两者对外源氮输入的响应可能不同。本研究选取羌塘高原高寒草甸和高寒草甸草原为研究对象,设置长期施氮梯度试验,对比研究外源氮输入增加对不同类型高寒草地的影响,并利用施氮梯度试验研究不同类型高寒草地的氮敏感性,为青藏高原高寒草地的管理提供科学指导。
1 研究区概况与实验方法
1.1 研究区概况
羌塘高原(29°53′―36°32′N, 78°41′―92°16′E)地处藏北高原腹地,是西藏高原的主体部分之一,平均海拔在4500 m以上。本实验分别在藏北羌塘高原高寒草甸和高寒草甸草原区开展。高寒草甸样地位于西藏自治区那曲县境内(31°34′N, 92°34′E, 海拔4570 m),气候为大陆性高原气候。研究区所在地点多年平均气温-0.9℃。多年平均降雨量444.9 mm,主要集中在6―8月。土壤为高寒草甸土,土壤有机质含量高。植被是以高山嵩草为优势种的高寒草甸,群落盖度在80%以上,并且伴生有紫花针茅、藏豆(Stracheyatibetica)等植物[19- 20]。高寒草甸草原样地位于西藏自治区班戈县境内(31°23′N, 90°14′E, 海拔4590 m),气候为大陆性高原气候。班戈县多年平均气温-1.0℃,多年平均降雨量335.4 mm,主要集中在6―8月。土壤为高寒草原土,土壤相对贫瘠。植被是以紫花针茅为优势种的高寒草甸草原,群落盖度在30%―50%,并且伴生有线叶苔草、弱小火绒草(Leontopodiumpusillum)、藏沙蒿(Artemisiaannua)、丛生棘豆(Oxytropiscaespitosula)等植物[19- 20]。
1.2 试验设计与样品采集
在高寒草甸和高寒草甸草原区分别选取植被均匀的区域进行长期施肥实验布设。样地布设采取随机区组设计,共设置5个区组,每个区组有5个重复。每个区组设置5种施肥处理,施肥梯度依次为对照(N0)、25(N25)、50(N50)、100(N100)、200(N200)kg N hm-2a-1。施肥小区大小为4 m × 4 m,相邻两个小区之间设置2 m的缓冲带。实验布设于2013年并开始进行养分添加处理,肥料种类为尿素,每年植物返青期(6月初)将肥料一次性施加进去。所有施肥区域用围栏围住,属于全年禁牧区,无牛羊放牧活动干扰。
植物群落特征调查采用样方法。2014―2017年每年植物生长旺盛期(8月中旬)进行,每年群落调查和样品采集一次。每种施氮处理随机选取三个重复进行调查和取样,每个样地内随机选取0.5 m × 0.5 m样方,将样方框用细绳均匀分为10 × 10个25 cm2的栅格,以植物群落所占栅格数确定群落总盖度及每个植物种的分盖度,并数出每种植物的株丛数。为了便于统计比较,本文中将所有植物物种划分为4个功能群:禾草、莎草、豆科和其他杂草。调查完成后齐地面、分物种剪取样方内植物地上部分,装入信封内带回实验室,置于65℃烘箱内烘干48 h至恒重,称重后换算到每平米作为地上生物量(精度为0.001 g)。在剪掉植物的样方四角和中间位置用土钻(直径3.8 cm)分别钻取20 cm深度的土壤,混合均匀作为一个土壤样品,每年在生长季旺盛期取样一次。过2 mm筛剔除植物根系和石块,风干后利用凯氏定氮法测定土壤全氮含量。不同样地的气象数据来自中国气象数据网(http://data.cma.cn/)。
1.3 数据计算与统计分析
群落物种综合α多样性指数采用如下公式进行计算。
(1)辛普森(Simpson)指数:
(2)香农-威纳(Shannon-Weiner)指数:

(3)Pielou均匀度指数:

式中:Pi代表第i个物种数量占群落总个体数量的比例,S代表样地内物种总数。
植物群落对施氮的敏感性用响应曲线来计算。采用Bowman等[13,16]的计算方法估算群落动态对外源氮输入的饱和阈值:将不同施氮剂量下群落生物量相对于不施氮处理改变的百分率进行回归拟合。有研究表明,群落生产随施氮量增加呈抛物线趋势,拟合曲线顶点对应的施氮量作为氮饱和阈值[13,16]。
以年份为重复测定因子,利用重复测定方差分析施氮对群落结构(物种数、多样性指数)与生物量(群落与各个功能群地上生物量)的影响。在每个采样年份,利用单因素方差分析研究施氮梯度对群落结构(物种数、多样性指数)与生物量(群落与各个功能群地上生物量)的影响。显著性水平为P<0.05,所有的统计分析在SPSS 16.0软件中进行,所有的图件用Origin Pro 8.0进行制作。
2 结果
2.1 群落结构与物种多样性
施氮对高寒草甸物种数的影响存在年际间差异(表 1,P=0.066)。2016年随着施氮量的提高,物种数逐渐增加,2017年呈现逐渐降低的趋势(图 1)。然而,施氮对高寒草甸物种多样性指数均无显著影响(图1; 表1,P>0.05)。高寒草甸草原呈现不同的规律。随着施氮量的提高,高寒草甸草原物种数和多样性指数均呈现逐渐降低的趋势(图1; 表 1,P<0.01),显著差异一般出现在N50或N100处理下。
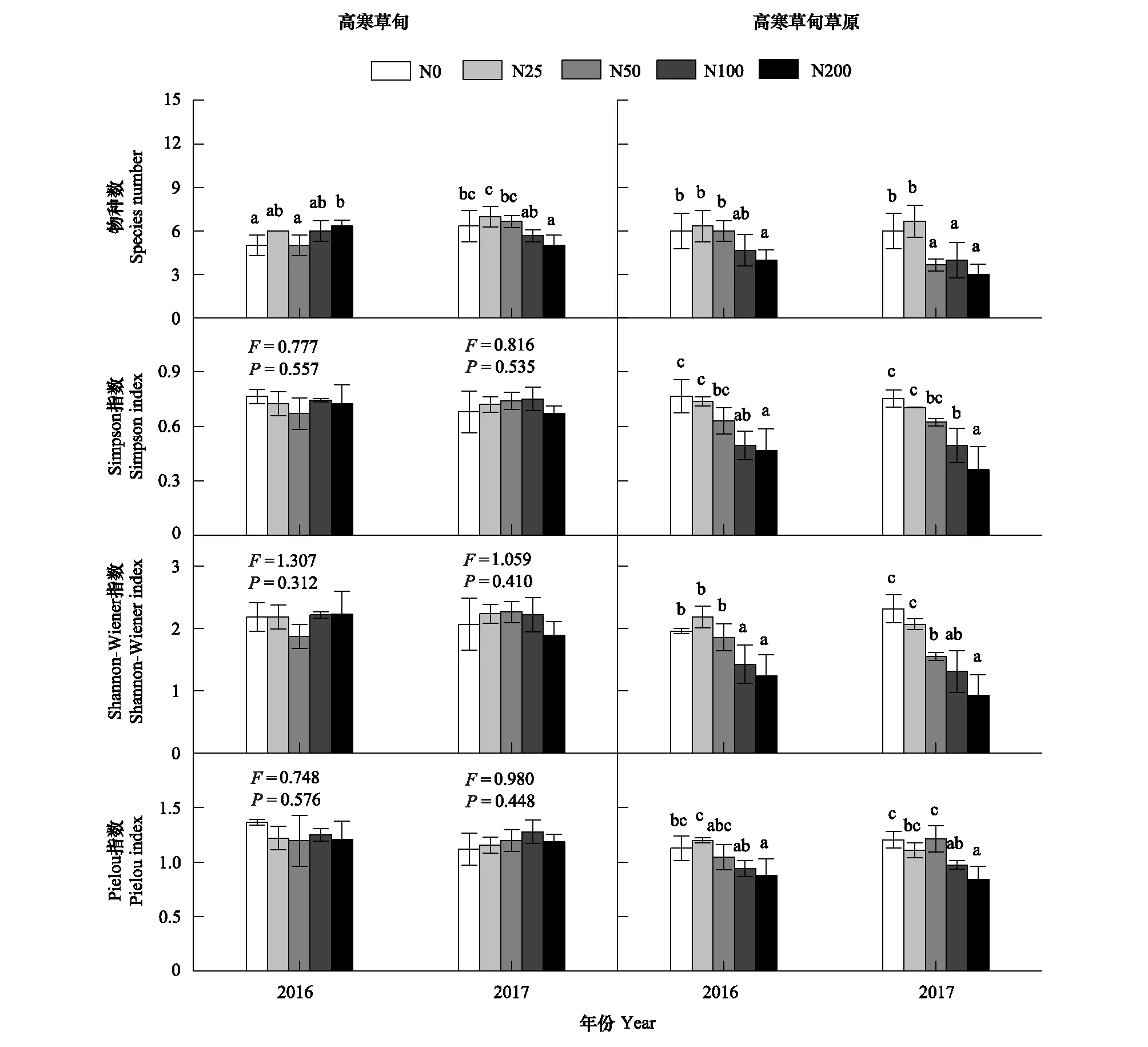
图1 施氮对高寒草甸和高寒草甸草原物种多样性、辛普森指数、香农威纳指数和Pielou多样性指数的影响Fig.1 Effects of nitrogen addition on plant species number, Simpson index, Shannon-Wiener index and Plelou index in alpine meadow and alpine meadow-steppe, respectivelyN0, N25, N50, N100, N200代表分别对照, 25, 50, 100, 200 kg N hm-2 a-1;图中数据代表每个处理4个重复的均值,误差棒代表标准差;相同年份的不同小写字母代表不同施氮梯度有显著差异(P<0.05),未标出的代表差异不显著(P>0.05)
2.2 群落及各植物功能群地上生物量
不管在高寒草甸还是高寒草甸草原,施氮对群落总地上生物量均产生显著影响(表1,P<0.001),且存在年际间差异(表1,P<0.001)。在高寒草甸,施肥第二年(2014年)随着施氮量提高,地上生物量呈现逐渐增加趋势(图2)。随着施肥时间的延长(2015年以后),地上生物量呈现先增加后降低的趋势,最大生物量出现在N50处理中(图2)。在高寒草甸草原,随着施氮量提高,地上生物量均呈现先增加后降低的趋势(图2)。
不管在高寒草甸还是高寒草甸草原,施氮对禾草植物地上生物量产生显著影响(表1,P<0.001),且存在年际间差异(表1,P<0.001)。开始施氮几年(2015年之前),随着施氮量提高,高寒草甸禾草植物地上生物量逐渐提高。随着施氮时间的延长(2016年以后),禾草植物地上生物量呈现先增加后降低的趋势,最大生物量出现在N50处理中(图2)。高寒草甸草原呈现不同的趋势。开始施氮几年(2015年之前),随着施氮量提高,禾草植物地上生物量呈现先增加后降低的趋势,最大生物量出现在N50或N100处理中。随着施氮时间的延长(2016年以后),高寒草甸草原禾草地上生物量逐渐提高(图2)。
不管在高寒草甸还是高寒草甸草原,施氮对豆科植物地上生物量的影响存在年际间差异(表1,P<0.001)。施氮对高寒草甸豆科植物地上生物量有显著影响(表1,P=0.001),施氮前几年(2015年之前)对豆科植物地上生物量的影响存在不一致规律。随着施氮时间的延长和施氮剂量的增加,豆科植物地上生物量呈现逐渐降低的趋势(图2)。重复测定方法分析显示,施氮对高寒草甸草原豆科植物地上生物量无显著影响(表1,P=0.112),从每年的分析来看,施氮对豆科植物地上生物量的影响只在施肥的后几年出现(图2)。
不管在高寒草甸还是高寒草甸草原,施氮对莎草植物地上生物量产生显著影响(表1,P<0.05),且存在年际间差异(表1,P<0.001),但对两者影响不同。随着施氮时间的延长,高寒草甸莎草植物地上生物量呈现不同的趋势。随着施氮剂量的提高,莎草植物地上生物量由施氮开始时(2014年)的逐渐增加转变为先增加后降低的趋势(2015年),最后变为逐渐降低的趋势(2017年)(图2)。这说明随着施氮时间的延长,施氮不利于莎草植物的生长。在高寒草甸草原,施氮对莎草植物地上生物量的影响呈现相对一致的趋势:随着施氮剂量的提高,莎草植物地上生物量呈现先增加后降低的趋势,最大生物量出现在N25或N50处理(图2)。
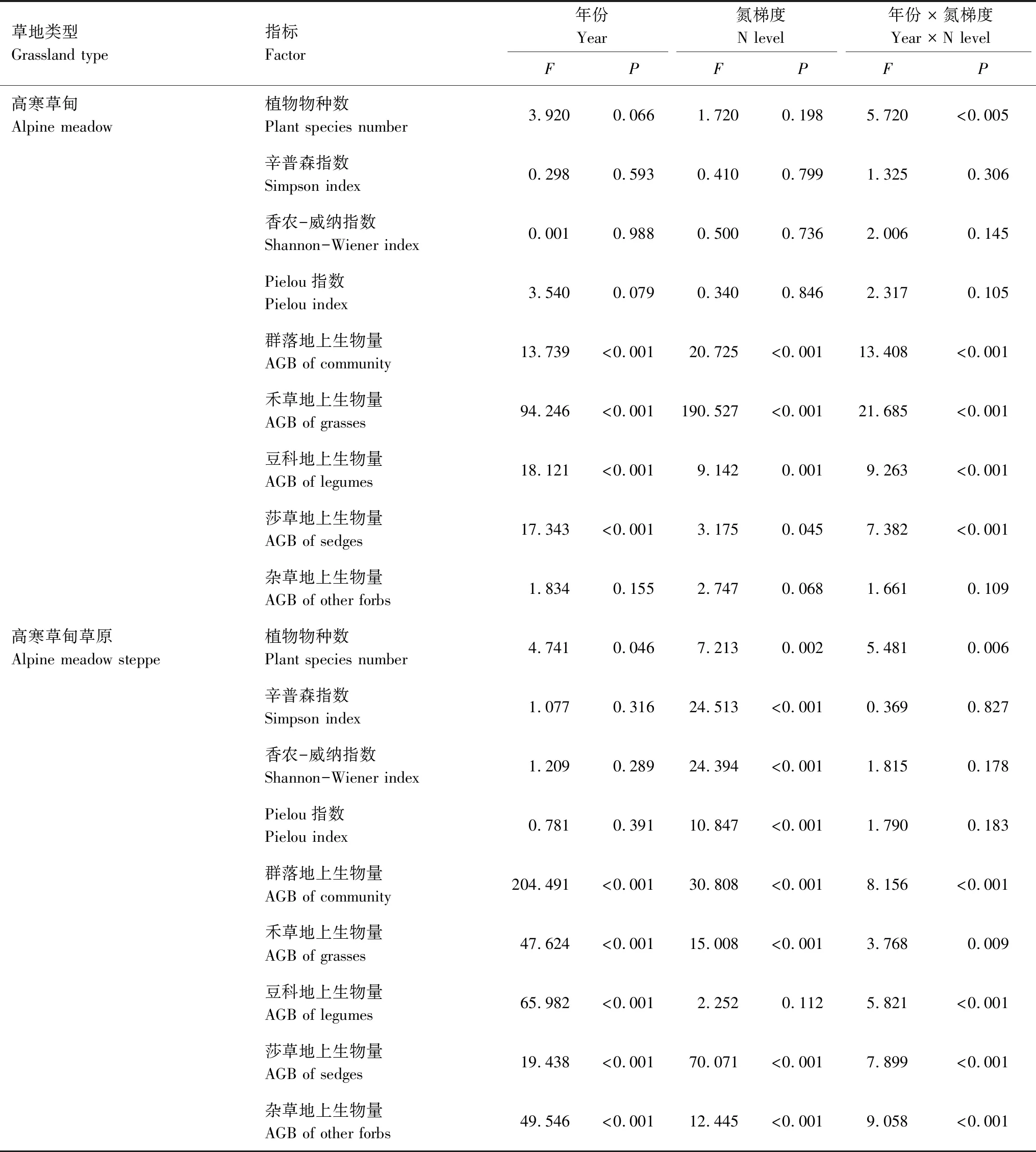
表1 利用重复测定分析方法对高寒草甸和高寒草甸草原植物物种数、辛普森、香农威纳、Pielou多样性指数,群落禾草、豆科、莎草和其他杂草植物地上生物量分析

图2 施氮对高寒草甸和高寒草甸草原群落总地上生物量的影响Fig.2 Effects of nitrogen addition on aboveground biomass of total community in alpine meadow and alpine meadow-steppe, respectively
重复测定方差分析显示,施氮对高寒草甸其他杂草植物地上生物量无显著影响(表1,P=0.068),年际之间也无显著差异(表1,P=0.155)。通过对每年施氮影响的单独分析发现,显著影响只出现在2016年,随着施氮剂量的提高,杂草植物地上生物量逐渐增加(图2)。施氮对高寒草甸草原其他杂草植物地上生物量存在显著影响(表1,P<0.001),年际之间也无显著差异(表1,P<0.001)。随着施氮剂量的提高,杂草植物地上生物量呈现先增加后降低趋势(图2)。
2.3 不同类型草地敏感性及其影响因子分析
禾草植物是对氮输入最敏感的植物功能群,所以利用禾草植物的反应进行氮饱和阈值的估算。不管是高寒草甸还是高寒草甸草原,随着施氮剂量的增加,禾草植物的响应比呈现先增加后降低的单峰趋势(图3)。高寒草甸和高寒草甸草原的饱和阈值分别是109.5、125.8 kg N hm-2a-1,高寒草甸草原对施氮的敏感性低于高寒草甸(图3)。

图3 不同高寒草地对施氮的敏感性分析Fig.3 Analysis on the sensitivity of different alpine grasslands to nitrogen addition H代表拟合抛物线顶点对应的氮梯度
通过对高寒草甸和高寒草甸草原降雨量分析发现,2013―2017年平均降雨量分别是498.3、343.6 mm,高寒草甸比高寒草甸草原降雨量高45%(图4,P=0.005)。同时,通过对两种高寒草地土壤总氮含量分析发现,高寒草甸总氮含量显著高于高寒草甸草原(图4,P<0.001)。进一步分析发现,两种高寒草地氮饱和阈值与降雨量、土壤基地养分含量均呈负相关关系(图4),即随着降雨量的降低高寒草地对外源氮输入越来越不敏感。这也说明高寒草地对氮输入的响应不仅受气候因子的调控,还受与气候因子相关的土壤基底养分含量的影响。
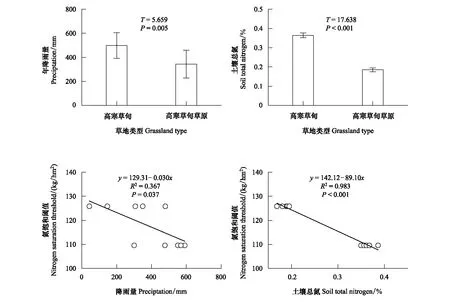
图4 不同高寒草地2013―2017年均降雨量与土壤总氮含量比较及其与氮饱和阈值的关系Fig.4 Comparison of precipitation from 2013 to 2017 and soil total nitrogen in different grassland types, as well as the correlations between these two factors and nitrogen saturation threshold
3 讨论
我们的研究的表明,外源氮输入对高寒草甸物种丰富度存在年际间差异:2016年随着施氮量的提高,物种丰富度逐渐增加,2017年呈现逐渐降低的趋势,且施氮对高寒草甸物种多样性指数均无显著影响。高寒草甸作为成熟稳定的高寒生态系统类型,对施氮的响应较慢。高寒草甸的优势物种为高山嵩草,属于寒中生密丛型多年生草本植物,须根系相当发达,与其他植物根系交织一起形成草毡层,对外源氮输入增加的响应不敏感。高寒草甸草原呈现不同的规律,随着施氮量的提高物种丰富度和多样性均呈现逐渐降低的趋势,这与多数已有研究结论一致。Bai等[11]研究了氮素添加对内蒙古成熟和退化草原生物多样性和生态系统功能的影响,发现氮素添加明显降低了成熟草原的物种丰富度。Ren等[21]通过添加氮、磷、钾和水等营养物质研究了不同组合的资源添加对高寒草甸植物群落物种丰富度和生产力的影响,结果表明,随着施加资源组合数量的增加物种多样性降低。一般来讲,氮肥作为速效养分,极易在短时间内被喜氮植物吸收利用,使其生物量极大增加。施肥提高了土壤中可利用资源的含量,使植物之间由地下部分对矿质资源的竞争转化为地上部分对光的竞争。物种之间对光的竞争降低了植物群落的物种多样性。Hautier等[22]认为,在施氮后的草原群落植被下层增加光照,可以有效地缓解植物多样性的降低。Ren等[21]认为物种之间对群落下层光的竞争是资源添加导致物种多样性降低的主要原因之一。同时,施氮导致的土壤理化性质的改变也是引起物种多样性降低丧失的重要原因[23- 24]。
施肥第二年,随着施氮量提高,高寒草甸地上生物量呈现逐渐增加趋势,随着施肥时间的延长地上生物量呈现先增加后降低的趋势;随着施氮量提高,高寒草甸草原地上生物量均呈现先增加后降低的趋势。这与已有的多数研究结论一致,即生态系统生产对氮输入增加存在饱和现象,即低氮输入会提高草地生产,而过量氮输入不仅不会持续增加草地生产,还可能会引起生产力降低[11,13- 14,16]。但高寒草甸在施肥开始初期表现出地上生物量逐渐增加的趋势,这说明高寒草甸生产受氮限制,高剂量施氮依然会促进群落生产。研究表明,生态系统对氮输入的响应并不总是线性的,氮输入导致的氮饱和会使得生态系统向不同的方向发展,氮饱和发生在当外源氮输入超过植物和微生物的需求时[25]。
对于外源氮输入增加,高寒草地中不同植物功能群的响应存在差异。随着施氮剂量的增加,高寒草甸禾草植物虽然表现出先增加后降低的趋势,但其在群落中的比例随着氮输入年份的增长逐渐增加,而莎草植物比例随着施氮时间的增长呈现逐渐降低的趋势。高寒草甸草原中莎草植物所占比例较低,随着施氮剂量的增加,莎草植物虽然表现出先增加后降低的趋势,所以高剂量施氮不利于莎草植物生长。与之相反的是,禾草比例随着施氮时间的延长,沿施氮梯度的增加逐渐升高。杂草植物比例随着施氮时间的延长,沿施氮梯度的增加逐渐降低。禾草是高寒草甸草原的植物优势功能群,长期氮输入使得禾草植物在群落中优势度进一步增加。禾草对氮输入反应敏感,氮肥作为速效养分,极易在短时间内被对氮素需求强烈的禾草植物吸收利用,从而使其生物量增加[26- 27]。同时,由于禾草植物高度比较高,处于植物群落的上方,不会受到光资源的限制[22],故其生产受到施肥处理的显著促进。长期氮输入会改变高寒草地的群落结构,使得禾草植物在群落中优势度逐渐增加。这虽然会增加可食性牧草所占的比例,但单一功能群植物的提高会造成生物多样性的丧失和群落稳定性的降低,不利于高寒草地对气候异常等外界干扰的抵抗。
同时利用禾草和群落总生物量的反应估算的高寒草甸和高寒草甸草原的氮饱和阈值分别是109.5、125.8 kg N hm-2a-1,高寒草甸草原对施氮的敏感性低于高寒草甸。高寒草甸地处羌塘高原东部半湿润区,对氮输入的反应比较敏感,但由于群落结构的差异,本研究高寒草甸的研究结果高于在青藏高原当雄高寒草原化草甸的研究结果(50 kg N hm-2a-1)[16]。高寒草甸草原的饱和阈值接近于内蒙古温带草原的研究结果,Bai等[11]研究发现,能引起温带草原地上生物量、物种丰富度和植物功能群组成产生显著变化的阈值是105 kg N hm-2a-1。已有研究表明,相对于较少降雨的干旱地区,高降雨量地区的施肥会引起更大群落生产力的变化。一般来讲,天然草地的植被生产力通常受水分和养分的共同限制,这在本文的相关分析中得到验证(图4)。降雨较少的干旱地区(如高寒草甸草原区)群落生产的主要限制因子是水分,低的水分可利用性导致添加的养分不能变成植被易于吸收利用的形态。降雨较多的地区(如高寒草甸区)水分不是群落生产的主要限制因子,而养分多寡变成主要的限制因素,故对外源养分输入更加敏感。高寒草甸草原地处高寒草甸向典型高寒草原的过渡地带,植物种类兼具高寒草甸和草原的特征,植物群落结构对外源氮输入响应更加显著。
确定不同类型高寒草地氮的饱和阈值对于不同类型草地的管理十分重要。受气候变化和人类过度放牧的共同影响,西藏高原高寒草地呈现大面积退化趋势[28]。对于退化草地的恢复改良来说,合理、平衡的施肥对退化草地的恢复与改良具有积极的作用[5- 6],如果施肥量高于氮饱和阈值,不仅不会改善草地质量,反而会引起群落功能降低、土壤酸化等一系列的环境问题,故施肥量的选择应该在不同类型高寒草地的氮饱和阈值内。
猜你喜欢
成都信息工程大学学报(2022年4期)2022-11-18
青海草业(2022年2期)2022-07-23
中国土壤与肥料(2021年5期)2021-12-02
海外文摘·文学版(2021年3期)2021-05-19
畜牧兽医科学(2019年24期)2019-09-10
活力(2019年21期)2019-04-01
南方农业·下旬(2017年11期)2018-01-09
中国饲料(2017年15期)2017-08-22
农业环境科学学报(2017年2期)2017-03-20
杂草学报(2015年2期)2016-01-04
