
高粱OFP 转录因子家族生物信息学及 盐胁迫下的表达分析
2020-07-29 08:24梁红凯陈辉龙杨溥原殷丛培梁永波崔江慧常金华
河北农业大学学报 2020年3期
梁红凯,陈辉龙,杨溥原,殷丛培,梁永波,崔江慧,常金华
(1.河北农业大学 农学院,河北 保定 071000;2.华北理工大学 生命科学学院,河北 唐山 063210)
高粱(Sorghum bicolor(L.) Moench)是世界五大谷类作物之一,抗旱、耐盐碱、耐瘠薄,具有粮、饲、酿造等多种用途[1]。
OFP 卵形家族蛋白(Ovate family protein,OFP),是植物所特有的一类转录因子。人们发现OFP 基因广泛参与植物多个组织器官的生长发育,如参与次生细胞壁的形成,胚珠和维管束的生长以及油菜素内脂(Brassinolide,BRs)的信号传 导[2]。OFP 相关基因首次在番茄中克隆,该基因的一个点突变导致翻译提前终止,从而使圆形果实变成梨形果实[3]。后续在辣椒中CaOvate 蛋白通过抑制CaGA20ox1 表达,改变果实形状[4]。在香蕉中MaOFP1 与香蕉MuMADS1 蛋白互作,共同控制果实发育[5]。同样在瓜类植物中也发现OFPs 是控制果实形状的重要基因[6]。在拟南芥、水稻基因组中分别含有18、31 个OFP 家族成员[7-8]。拟南芥中AtOFP1在抑制细胞伸长生长中发挥重要作用[9],AtOFP8参与逆境胁迫过程,其对干旱胁迫和高温胁迫较为敏感[10],在水稻中过表达OsOFP6基因可提高水稻抗旱和抗冷能力[11]。目前对OFP 家族的研究主要集中在植物发育上,而在植物逆境调控中的研究尚不充分。因此本研究利用高粱基因组数据对高粱的OFP 家族基因进行筛选和鉴定,全面分析其基因序列特征及与水稻、玉米、谷子等物种间的共线性,并通过盐胁迫分析高粱OFPs 基因在盐胁迫条件下的表达特征, 为进一步研究高粱OFP 基因家族的功能和OFP 在植物逆境胁迫中的作用提供依据。
1 材料方法
1.1 数据的获取与鉴定
高粱(Sorghum bicolor(L.) Moench)、水稻(OryzasativaL.)、 谷子(Setaria italica)、 玉米(Zea maysL.)基因组数据从Ensamble 数据库(http://plants.ensembl.org/index.html)中获取,从Pfam 数据库(http://pfam.xfam.org/family/PF04844) 下载OFP 保守结构域蛋白序列的HMM 文件(PF04844),基于下载的HMM 模型使用HMMER 软件对高粱基因组蛋白质数据进行OFP 家族成员鉴定,选取E-value<0.001。获得候选的OFP 家族成员后经pfam(http://pfam.xfam.org/)、NBCI CDD(https://www.ncbi.nlm.nih.gov/cdd/) 和SMART(http://smart.embl.de/)等3 大数据库进一步确认高粱OFP结构域,含有OFP 结构域的成员才被认定为OFP家族成员。
1.2 蛋白信息和理化性质分析
利用ExPASy-ProtParam 在线工具(http://web.expasy.org/protparam/)分析高粱OFP 家族蛋白的氨基酸数目、分子量、等电点和亲水性[12]。
1.3 进化分析
利用MEGA7.0[13]中的ClustalW 对物种中的OFP 全长蛋白序列进行比对,基于邻接法(Neighbor-Joining),Bootstrap 值设为1 000,其余参数默认的条件下,绘制出OFP 家族的系统进化树。
1.4 基因在染色体上的位置
在高粱基因组序列DNA 注释文件中获取高粱OFP 家族基因在染色体上的信息,使用软件MapChart2.32[14]绘制基因染色体分布图。
1.5 基因结构、motif 及保守结构域分析
利用CFVisual 软件根据高粱OFP 家族GFF 文件和CDD 网站进行结构域预测,结果文件分别绘制基因结构图和结构域分布图,利用MEME[15]软件对高粱OFP 家族成员进行保守基序分析,Motif 数量设置为8 个。
1.6 顺式作用元件分析
提取高粱OFP 家族基因上游1 500 bp 长度的序列作为启动子序列,并提交到Plant CARE[16](http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)数据库,对高粱OFP 家族基因进行顺式作用元件分析。
1.7 共线性分析
使用python 版本的MCScanX[17]软件,分别对高粱内部和高粱与水稻、玉米和谷子的物种间共线性进行分析,并利用Circos-v0.69 软件和绘图程序将结果可视化。
1.8 OFPs 基因在盐胁迫下高粱中的表达模式分析
根据本课题组在NaCl 胁迫高粱幼苗获得的转录组数据,将耐盐高粱品种‘高粱蔗’(GZ)和盐敏感品种‘河农16’(HN)在不同盐胁迫时间下SbOFPs基因的表达,通过表达量的log2 比值进行聚类,利用TBtools 软件显示聚类图,分析SbOFPs基因在不同品种中的表达模式。
2 结果分析
2.1 高粱OFP 转录因子家族鉴定和理化性质分析
基于HMM 模型在高粱基因组中搜索鉴定得到37 个高粱OFP 基因家族成员,通过生物信息学分析,这37 个OFP 转录因子的蛋白等电点在4.49 ~11.62 之间,蛋白质大小在139 ~543 个氨基酸之内,氨基酸平均个数为302 个,分子量大小 在15.379 88 ~60.055 64 kD 之间。总平均亲水性(GRAVY)的范围在-0.768 和-0.043 之间,均为负值,表明这些蛋白均为亲水蛋白。
2.2 禾本科OFP 家族系统发育树分析
利用37 个高粱OFP 家族成员和粳稻30 个、谷子27 个、玉米的43 个OFP 基因全长蛋白序列,构建系统发育树(图1)。根据发育树可将OFP 基因家族分为group1、group2、group3 三大类,同时可以看出,系统树的大部分分支上是1 个高粱基因对应1 个谷子、1 个水稻、1 个玉米的比例分布,表明该家族进化非常保守。谷子和水稻的OFP 家族成员大多数处于一个单独分支上,高粱和玉米OFP 家族成员聚集在一起较多,这一现象与禾本科物种进化相符[18]。
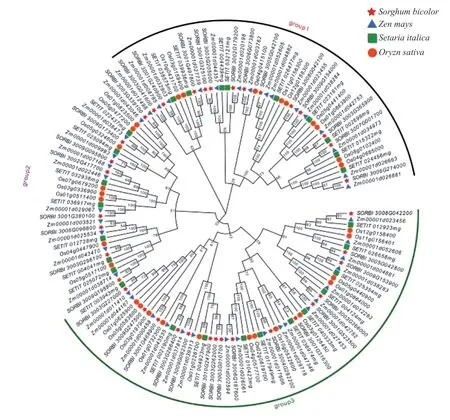
图1 高粱、水稻、谷子和玉米OFP 基因家族成员的系统发育树Fig.1 Phylogenetic tree of OFP genes in Sorghum bicolor, Oryza sativa, Setaria italica and Zea mays
2.3 高粱OFP 转录因子染色体定位分析
根据基因的染色体定位信息(图2),发现高粱的10 条染色体上包含了其OFPs 家族的所有37个基因,且呈不均匀分布,从单条染色体来看,3号染色体上分布最多,含有10 个SbOFPs基因,其次是1 号和9 号均为7 个SbOFPs基因分布,2 号、5 号、8 号均含有2 个SbOFPs基因,4 号、7 号和10 号所含SbOFPs基因最少均只有1 个。总体来看,大多数的高粱OFP 基因分布在染色体的两端区域且表现聚集趋势。

图2 高粱OFP 基因染色体定位Fig.2 Mapping of the OFP transcription factor gene family on Sorghum bicolor chromosomes
2.4 高粱OFP 基因家族保守基序和基因结构分析
对高粱的OFP 家族蛋白序列进行保守基序分析(图3B)。结果显示,除SORBI_3003G262000只有Motif1 保守基序以外,其它OFP 家族蛋白保守结构域均含有Motif1 和Motif2,另外还有其它的Motif 类型分别存在于26 个基因中。值得注意的是,SORBI_3009G095800、SORBI_3009G173400和SORBI_3003G3391003 个基因不仅具有OFP 家族共有的保守结构域,还具有DNA_binding_2 结构域。另外,对37 个高粱OFP 基因家族成员的结构分析显示(图3C),基因的内含子数和外显子数在高粱OFP 基因家族中存在很大的差异,SORBI_3009G247500、SORBI_3010G247900、SORBI_3003G262000、SORBI_3006G096600、SORBI_3002G417700、SORBI_3006G073800和SORBI_3003G365900含有1 个内含子,其长度相对较短,SORBI_3008G042100含有2 个内含子,其余成员则不包含内含子。

图3 高粱OFP 基因家族保守基序和基因结构分析Fig.3 Conservative motifs and gene structure analysis of the Sorghum OFP gene family
2.5 启动子顺式作用元件预测分析
对37 个高粱OFPs 家族上游1 500 bp 的启动子顺式作用元件的分析结果发现(图4),与生长发育、激素调控和抗逆等相关元件在37 个OFPs家族成员中都有分布,SORBI_3001G316300,SORBI_3003G205000,SORBI_3009G082300,SORBI_3009G095800,SORBI_3008G042100,5 个基因含有防御和胁迫反应元件(TC-rich repeats);SORBI_3001G468900,SORBI_3001G523500,SORBI_3001G316300,SORBI_3003G286400,SORBI_3003G296100,SORBI_3003G227000,SORBI_3005G042800,SORBI_3006G187600,SORBI_3007G001700,SORBI_3009G082300,SORBI_3009G173400,SORBI_3010G247900,SORBI_3009G154000,SORBI_3006G274000,SORBI_3003G041600,SORBI_3003G339100,SORBI_3009G247500,SORBI_3004G282200,SORBI_3003G365900,18 个基因均含有干旱诱导响应元件(MYB binding site,MBS), 其中SORBI_3003G286400中含有最多,有4 个干旱诱导响应元件,其次是SORBI_3009G154000含有3个。32 个基因具有脱落酸响应元件(Abscisic acid responsiveness,ABRE),29 个基因具有茉莉酸甲酯(MeJA)响应元件(TGACG-motif 和CGTCAmotif),11 个基因有水杨酸响应元件(TCA-element)和12 个基因具有赤霉素响应元件(TATC-box,P-box 和GARE-motif)。其中,同时具有干旱诱导响应元件(MBS)、脱落酸响应元件(ABRE)和MeJA 响应元件(CGTCA-motif 和TGACG-motif)的共有13 个基因,分别为SORBI_3001G468900、SORBI_3003G296100、SORBI_3001G523500、SORBI_3003G227000、SORBI_3003G286400、SORBI_3005G042800、SORBI_3009G173400、SORBI_3006G187600、SORBI_3007G001700、SORBI_3003G339100、SORBI_3009G247500、SORBI_3004G282200和SORBI_3003G365900, 这13个基因可能在干旱胁迫等非生物胁迫中起主要作用。
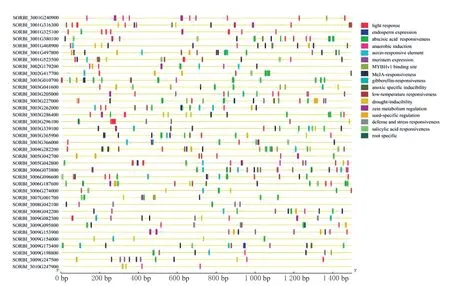
图4 高粱OFP 家族基因启动子顺式作用元件功能Fig.4 Functions of promoter cis-acting elements in OFP family genes of Sorghum bicolor
2.6 共线性分析
基因家族加倍可能是由于同一条染色体上由基因的自我复制产生两个或者多个基因距离很近的串联[19-20]或物种进化过程中染色体的结构变异导致[21]。基因复制在基因扩张和基因的功能分化中发挥重要的作用,经对高粱OFP 家族片段重复基因的共线性分析发现(图5),其中发生20 对基因的重复片段较高,SORBI_3006G187600分别与SORBI_3001G316300、SORBI_3001G523500和SORBI_3004G282200;SORBI_3001G523500分 别与SORBI_3006G187600、SORBI_3001G316300和SORBI_3004G282200;SORBI_3001G316300分 别与SORBI_3001G523500、SORBI_3006G187600和SORBI_3004G282200;SORBI_3004G282200分 别与SORBI_3006G187600、SORBI_3001G316300和SORBI_3001G523500重复较多。另外在对高粱和水稻、谷子、玉米OFP家族基因进行共线性分析发现(图6),有27 个高粱OFP 家族基因与27 个水稻OFP家族基因显示出共线性,与27 个谷子的全部OFP家族基因显示出共线性,28 个高粱OFP 家族基因与32 个玉米OFP 的基因显示出共线性,表明该基因家族在进化过程中物种间相对保守。
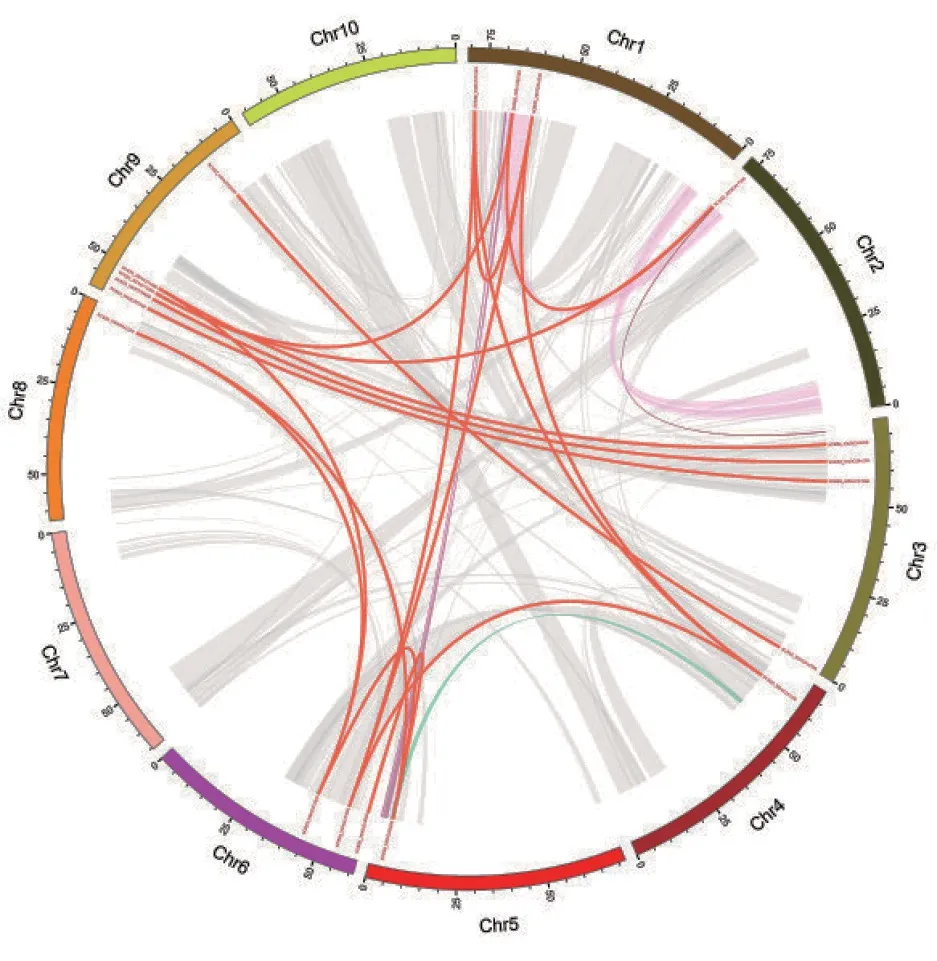
图5 高粱OFP 基因家族片段重复基因的circos 分析Fig.5 Circos analysis of fragment repeat genes of SorghumOFP gene family

图6 高粱与水稻、谷子和玉米的共线性图谱Fig.6 Synteny analysis of OFP genes between Sorghum bicolor and Oryza sativa, Setaria italica, Zea mays
2.7 OFP 基因在不同耐盐性高粱品种中的表达分析
利用转录组数据,对不同耐盐性高粱品种在NaCl 胁迫48 h 和72 h 后,高粱OFP 家族基因的表达进行了分析,2 个品种经盐胁迫处理后表达差异较大的基因(图7)主要有SORBI_3008G042200、SORBI_3005G042800、SORBI_3001G316300、SORBI_3003G227000、SORBI_3003G010700、SORBI_3003G366000、SORBI_3006G187600、SORBI_3010G247900。 其中SORBI_3003G227000在2 个品种盐胁迫后,表达量均表现上升趋势,SORBI_3003G010700在GZ 当中先升后降,但受盐胁迫后48 h 和72 h 的表达量均高于对照,而在HN 品种中表现上升趋势。SORBI_3003G366000和SORBI_3006G187600在2 个品种中的表达量均先升后降,而在耐盐的GZ 中的表达量均高于盐敏感品种HN,与2 品种的耐盐性特征相一致。这些基因随盐胁迫的变化特征暗示这些基因可能参与高粱盐胁迫的响应。

图7 高粱OFP 基因在GZ 和HN 品种中不同时间盐胁迫的表达Fig.7 Sorghum OFP gene expression in GZ and HN varieties at different times under salt stress
3 讨论
OFP 基因家族是一种新型的植物特异性转录因子,在植物的生长发育、生物和非生物胁迫中发挥着重要的调控作用[22-23]。在OsOFP6过表达的转基因水稻植株中,发现OsOFP6可以防止干旱胁迫对水稻的伤害[11]。在苹果受NaCl、PEG、高温和低温4 种胁迫处理后,MdOFP不同程度地响应盐、干旱和温度胁迫,推测MdOFP可能参与调控苹果的盐、干旱与温度逆境响应过程[24]。可见,OFP家族在应对胁迫中发挥着重要作用。
高粱具有对干旱、盐碱等多种不良环境的耐受性,为了更好的认识高粱OFP 基因家族及其在应对环境胁迫中的作用,本文对高粱OFP 基因家族进行了系统分析。在高粱的亲缘物种水稻、谷子和玉米中分别鉴定出30 个、27 个和43 个OFP 基因,从四个物种OFP 成员数量上看,OFP 基因家族并没有发生大规模的扩增,其中玉米的OFP 家族成员较多的原因,可能与玉米的第二次加倍事件有关[18,25]。依据高粱OFP 的蛋白质序列同水稻、谷子和玉米的OFP 蛋白序列构建的进化树将其分为3 类,其分类结果与玉米OFP 基因家族分组情况类似,通过基因结构分析显示,高粱OFP 多数基因缺少内含子结构,与前人[26-27]研究一致,推测OFP 基因家族在进化过程中相对保守。在盐胁迫条件下高粱OFP 基因的表达分析显示,SORBI_3005G042800、SORBI_3003G227000、SORBI_3003G010700和SORBI_3006G187600的表达量变化与盐胁迫及品种的耐盐性具有一定的相关性,这些基因在功能启动子顺式元件中预测到同时具有3 个干旱响应元件,暗示着这些基因可能同时在响应盐胁迫中起着重要作用。盐害主要是由渗透胁迫和离子毒害造成的,从渗透胁迫角度来看,植物的耐盐和耐旱密切相关,增强抗旱能力的同时一般也会提高耐盐性[28]。 Agarwal[29]等研究表明让盐信号经ABA 识别后传给MYC/MYB 转录因子,MYB/MYC 转录因子进而调节抗逆基因的表达应答盐胁迫。因而认为SORBI_3005G042800、SORBI_3003G227000、SORBI_3003G010700和SORBI_3006G187600可能同时在高粱应对干旱胁迫和盐胁迫中发挥着重要作用,具体如何参与逆境胁迫的响应有待进一步验证。
本研究对高粱OFP 家族基因进行了综合分析,为高粱OFP 基因家族的进化特征提供了有价值的线索,研究结果为进一步研究单个SbOFP基因的生物学功能,挖掘OFP 家族的应用潜力奠定了基础。
猜你喜欢
湖北农业科学(2022年11期)2022-07-18
河北农业大学学报(2022年2期)2022-04-26
当代陕西(2021年21期)2022-01-19
广西植物(2021年1期)2021-03-24
科学与财富(2021年3期)2021-03-08
实用肿瘤学杂志(2020年4期)2020-12-08
温州大学学报(自然科学版)(2019年2期)2019-06-04
现代商贸工业(2019年5期)2019-02-18
延河(2018年6期)2018-06-11
医学综述(2011年12期)2011-12-09
