
‘李光杏’花芽分化时期内源激素及碳氮比值的动态研究
2020-07-28 04:04陈翠莲张继强刘生虎朱祖雷朱燕芳王延秀
干旱地区农业研究 2020年3期
赵 通,陈翠莲,程 丽,张继强,刘生虎,郭 荣,朱祖雷,朱燕芳,王延秀
(1.甘肃农业大学园艺学院,甘肃 兰州 730070;2.甘肃省林业科学研究院,甘肃 兰州 730070;3. 甘肃省敦煌市林业技术推广中心,甘肃 敦煌 736200)
‘李光杏’ (A.vulgarisvar.glabraSun S. X)属蔷薇科杏属,因产地又名‘敦煌李光杏’,是甘肃省敦煌市特产,得益于当地气候地理条件,以味道鲜美可口而闻名,深受人们喜爱[1]。‘李光杏’抗寒、抗旱、耐瘠薄、适应性强,具有较高的经济效益和生态效益。但‘李光杏’开花时间早(3月中旬),花期持续时间长,由于花期常受低温冻害的影响,花瓣脱落、雌蕊退化等已成为制约‘李光杏’发展的主要因素[2]。已有不少学者对各种杏品种的花芽分化有过报道,如:极早熟品种‘骆驼黄’[3]、新疆新品种[4]及‘兰州大接杏’和‘龙王帽’[5]等。而对于‘李光杏’花芽分化的研究鲜有报道。
花芽分化是果树生殖过程的基础,与果实的产量和品质密切相关。大量的研究表明,内源激素水平对于调控植物的花芽分化过程有着重要作用[6-8]。吴月燕等[9]研究认为,高浓度的CTK和低浓度的GA对杜鹃成花有促进作用。高比值的ABA/IAA、ABA/GA和ZR/GA均有利于罗汉果的花芽分化[10]。樊卫国等[11]在研究刺梨花芽分化时发现,GA与IAA含量下降,ZR与ABA上升对成花有促进作用。而Mohanty[12]证实ABA对成花有促进或抑制的双重作用。此外,碳氮含量的积累和转运是影响花芽分化的重要因素之一,其在营养旺盛生长期和花芽分化初期叶片的含量决定了产量高低[13-14]。在营养生长期,矿质营养主要存于叶片,分化至生殖生长期矿质营养被转运到芽和花中,促进花芽分化[15]。碳氮物质的总含量之比决定植株成花,相对多碳少氮的内环境利于成花。较高的C/N能提早桃花花芽分化时间,有利于芽体形态分化的持续进行,而低水平的C/N导致了梨僵芽的发生[17-18]。花芽分化和开花是形成产量的基础,因此有必要对‘李光杏’成花进行研究。
本研究以‘李光杏’为试材,探讨‘李光杏’花芽分化的进程,定量分析了花芽分化期内源激素、全碳、全氮含量及C/N的动态变化,了解内源激素及营养物质变化与成花之间的关系,以期为外用生长调节物质调控花芽分化提供一定的理论依据。
1 材料和方法
1.1 试验地概况
本试验在甘肃省敦煌市合水村(92°21′~94°34′E,40°53′~42°27′N)进行。该地属于温带大陆性气候,年平均降水量40 mm,蒸发量2 522 mm,全年日照时数为3 246.7 h;年平均气温为9.4℃,月平均最高气温为24.9℃(7月),月平均最低气温为-9.3℃(1月),年平均无霜期142 d,土壤类型为灌淤土。
1.2 试验材料及试验设计
供试材料为8 a生结果的‘李光杏’,株行距2.0 m×3.0 m,主干直径达13 cm,树冠直径1.5~2.2 m。行间设置排水沟,田间采用常规管理。从2018年6月1日至10月31日,每隔 10 d于上午10∶00采取树冠外围(东、南、西、北)短枝顶芽30个,用FAA(70%乙醇90 mL+冰醋酸5 mL+福尔马林5 mL)固定,保存备用。石蜡切片后在显微镜下观察花芽形态分化的时期。
2018年于6月1日至10月31日每月初随机从6棵‘李光杏’树体采取树冠外围不同类型果枝(短果枝5~15 cm、中果枝15~30 cm、长果枝30 cm以上、花束状果枝)上的叶片及顶花芽,取样后用液氮速冻,立即置于干冰中带回实验室,置于-20℃以下低温冰箱中保存,用于测定花芽及叶片内源激素含量。
于6月1日至10月31日每月初随机从6棵‘李光杏’树体采取树冠外围不同类型果枝上的花芽,带回实验室置于105℃烘箱中杀青20 min,然后在75℃下烘干,碾碎干燥保存,用于测定全碳、全氮含量,试验测定设6次重复。
1.3 试验指标的测定
1.3.1 花芽形态分化的观察 采用常规石蜡切片法制片,将固定液中的芽取出,放入盛有50%乙醇的培养皿中,用镊子与解剖针小心剥去鳞片,之后移入装有蒸馏水的小瓶中,再用注射器吸干水,然后加入2~3滴爱氏苏木精整染,按照步骤(材料冲洗→脱水→透明→浸蜡→包埋→修蜡→切片与贴片→展片与封片)进行,并用Motic Images Advanced 3.2显微成像系统软件对封好的片子进行观察并拍照。每个分化时期最少制作15张切片,其中将1个芽进入某一分化期定为该时期始期,10个芽进入某一分化期定为该时期末期。
1.3.2 内源激素的测定 试剂盒采用双抗体夹心法酶联免疫吸附试验(ELISA)。往预先包被植物赤霉素(GA)、脱落酸(ABA)、细胞分裂素(CTK)、吲哚乙酸(IAA)、玉米素核苷(ZR)捕获抗体的包被微孔中,依次加入标本、标准品、HRP标记的检测抗体,经过温育并彻底洗涤。用底物TMB显色,TMB在过氧化物酶的催化下转化成蓝色,并在酸的作用下转化成最终的黄色。颜色的深浅和样品中的植物赤霉素(GA)、脱落酸(ABA)、细胞分裂素(CTK)、吲哚乙酸(IAA)、玉米素核苷(ZR)呈正相关。用酶标仪在450 nm波长下测定各种激素的吸光度(OD值),计算样品中各激素的浓度。
1.3.3 矿质元素的测定 样品中全碳含量采用燃烧滴定法,全氮含量测定用凯氏法[19]。
1.4 数据处理与分析
应用Excel 2013及Origin 2017软件进行数据处理及作图,运用SPSS 22.0进行数据方差分析。统计分析采用单因素ANOVA的LSD比较处理间差异显著性。
2 结果与分析
2.1 ‘李光杏’花芽形态分化时期的划分及观察
‘李光杏’花芽形态分化开始早,延续时间长,从6月20日开始至10月初结束,约需80多天,且各期均有重叠现象,生理分化不稳定,是内部生理状态的改变,而无形态上的差异,显微结构上观察到的分化表明其已经通过生理分化,进入形态分化。‘李光杏’花芽形态分化自6月下旬开始,7月下旬大部分花芽已进入萼片分化期,但仍有少数花芽处于分化初期。李光杏进入初分化期的时间虽然相差较大,但在落叶前雌蕊分化均已完成。根据对‘李光杏’花芽纵剖面切片的观察,将‘李光杏’花芽形态分化划分为7个时期,分别是:未分化期、分化初期(花序原基分化期)、萼片分化期、花瓣分化期、雄蕊分化期、雌蕊分化期、花粉细胞期。
2.1.1 未分化期 此时‘李光杏’的芽体较小,已有叶原基分化,且鳞片紧抱,芽的生长椎尖狭小,细胞体积较小(图1A)。
2.1.2 分化初期 生长点开始变得肥大,向上隆起呈馒头状。观察发现,此时生长锥外侧出现花序原基,且两侧可见到一些凸起。分化初期最早出现在6月20日,但可以看出8月10日左右仍有少数花芽处在分化初期(图1B)。
2.1.3 萼片分化期 花原基顶部先变平坦,然后其中心部分相对凹入而四周产生突起,即萼片原始体。萼片分化期在7月20日到8月底,且分化盛期在8月10日左右(图1C)。
2.1.4 花瓣分化期 可以看到随着萼片的伸长生长,萼片内侧基部发生突起体,即为花瓣原基。从8月10日开始进入花瓣分化期,分化盛期一直可延续到9月初(图1D)。
2.1.5 雄蕊分化期 可以看出花瓣原基内部会出现两轮凸起(多排列为上、下两层),此部分为雄蕊原始体,进入雄蕊分化期。最早出现在8月20日,分化盛期在8月底,也可延续到9月10日左右(图1E)。
2.1.6 雌蕊分化期 明显可见,此期花序原基上的花粉囊呈锯齿状,且花粉囊已突出于花序原基,花蕾原始体的中心部出现一个向上生长的突起,即为雌蕊原基。雌蕊分化期在8月20日开始,分化盛期在9月10日左右,一直可延续到9月底完成(图1F,图1G)。
2.1.7 花粉细胞期 自10月份以后花粉囊壁内出现花粉母细胞,且排列紧密,呈多角形,整个花蕾体积不断增大,同时膨大的子房体内部珠心原始体开始分化(图1H,图1I)。
2.2 ‘李光杏’花芽形态分化时期内源激素含量的变化
由图2可知,花芽中IAA含量的变化呈逐渐上升趋势(图2A),10月之后无显著变化,此时大多数花芽形态分化已经完成,6月份(未分化期)含量最低,为129.6 ng·g-1,9月份(雌蕊分化期)最高为239 ng·g-1,相比6月份高出77.5%。花芽中CTK含量变化呈先降后升再降趋势(图2 B),6月份未进入形态分化期,此时CTK含量最高(448 ng·g-1),之后显著下降,7月份出现最低值(349.2 ng·g-1),相比6月份(未分化期)降低了22.1%,9月开始上升,此时大多数花芽已进入雌蕊分化期,CTK含量为439.49 ng·g-1,与6月份(未分化期)相比升高2.3%。
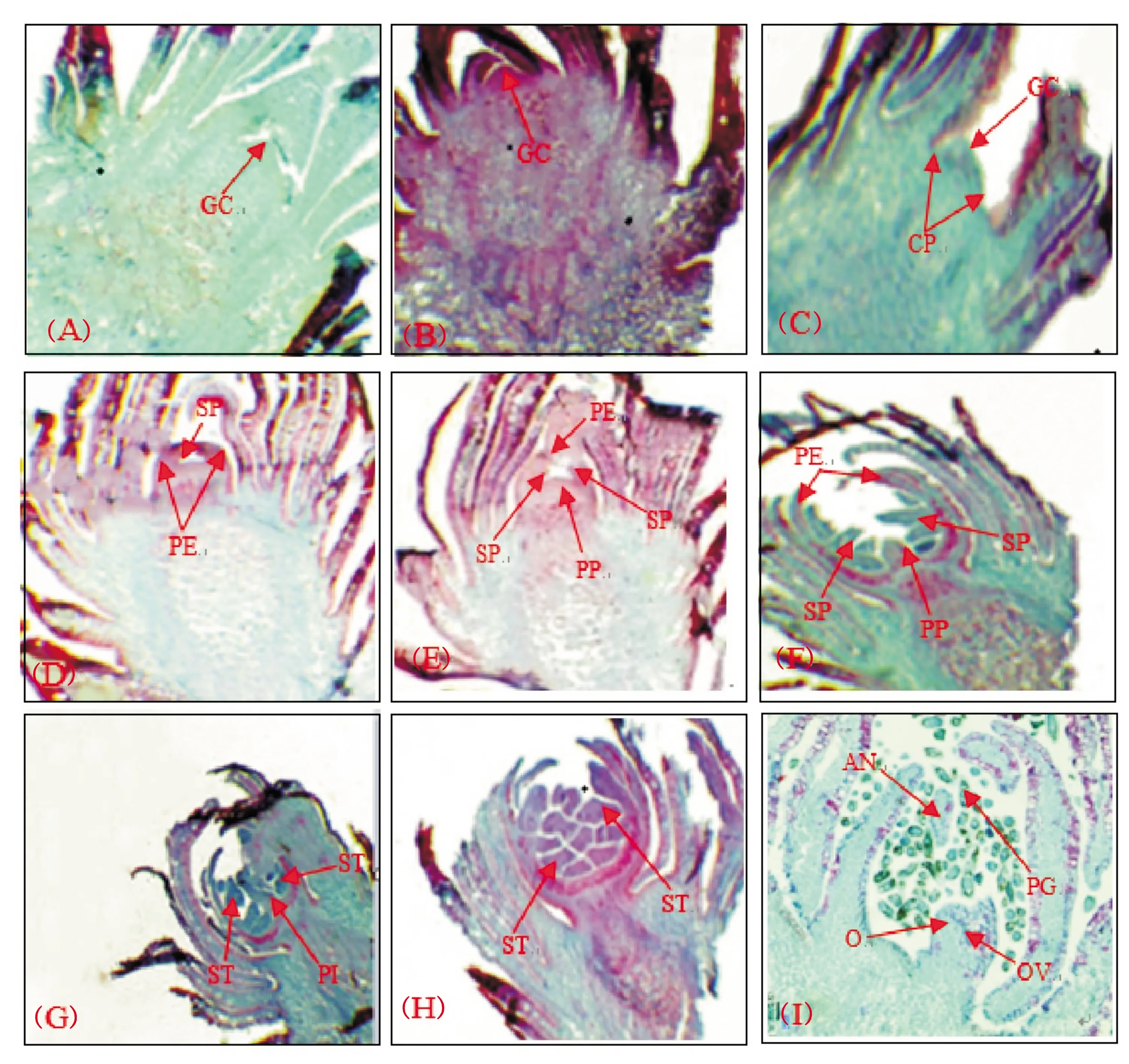
注:(A):未分化期(×10);(B):花芽分化初期(×10);(C):萼片原基分化期(×10);(D):花瓣原基分化期(×10);(E):雄蕊原基分化期(×10);(F)、(G):雌蕊原基分化期(×10);(H)、(I):花粉细胞期(×10);GC:生长锥;CP:花萼原基;PE:花瓣原基;SP:雄蕊原基;PP:雌蕊原基;ST:雄蕊;PI:雌蕊;AN:花药;PG:花粉;O:子房;OV:胚珠Note: (A): Undifferentiated stage (×10); (B): Flower bud differentiation stage (×10); (C): Sepal primordium differentiation stage (×10); (D): Petal primordium differentiation stage (×10); (E): Stamen primordium differentiation stage (×10); (F) and (G): Pistil primordium differentiation stage (×10); (H) and (I): Pollen cell stage (×10); GC: Growing tip; CP: Calyx primordial; PE: Petal primordial; SP: Stamen primordial; PP: Pistil primordial; ST: Stamen; PI: Pistil; AN: Anther; PG: Pollen; O: Ovary; OV: Ovule图1 ‘李光杏’花芽形态分化时期的划分及观察Fig.1 Division and observation of flower-bud morphological of Li-Guang Apricot at different periods
花芽中GA含量变化呈先升后降趋势(图2C),6、7、8月无明显的变化,分别为6月(232.4 ng·g-1),7月(221.9 ng·g-1),8月(222.1 ng·g-1),9月份显著上升,此时大多数花芽处于雄蕊、雌蕊分化期,出现最高值,为317 ng·g-1,10月份花芽形态分化完成后逐渐下降。花芽中ABA变化呈先降后升趋势(图2D),6月份(未分化期)ABA含量最高,为491.7 ng·g-1,8月份ABA含量最低(243 ng·g-1),与6月份相比下降50.5%,在花芽形态分化完成之后(10月初)ABA含量逐渐上升,但略低于6月份。花芽中ZR的含量在6、7、8月无显著差异(图2E),但9月份雌蕊、雄蕊分化期升至最高,为64.8 ng·g-1,与6月份(52.6 ng·g-1)相比升高了23.2%。
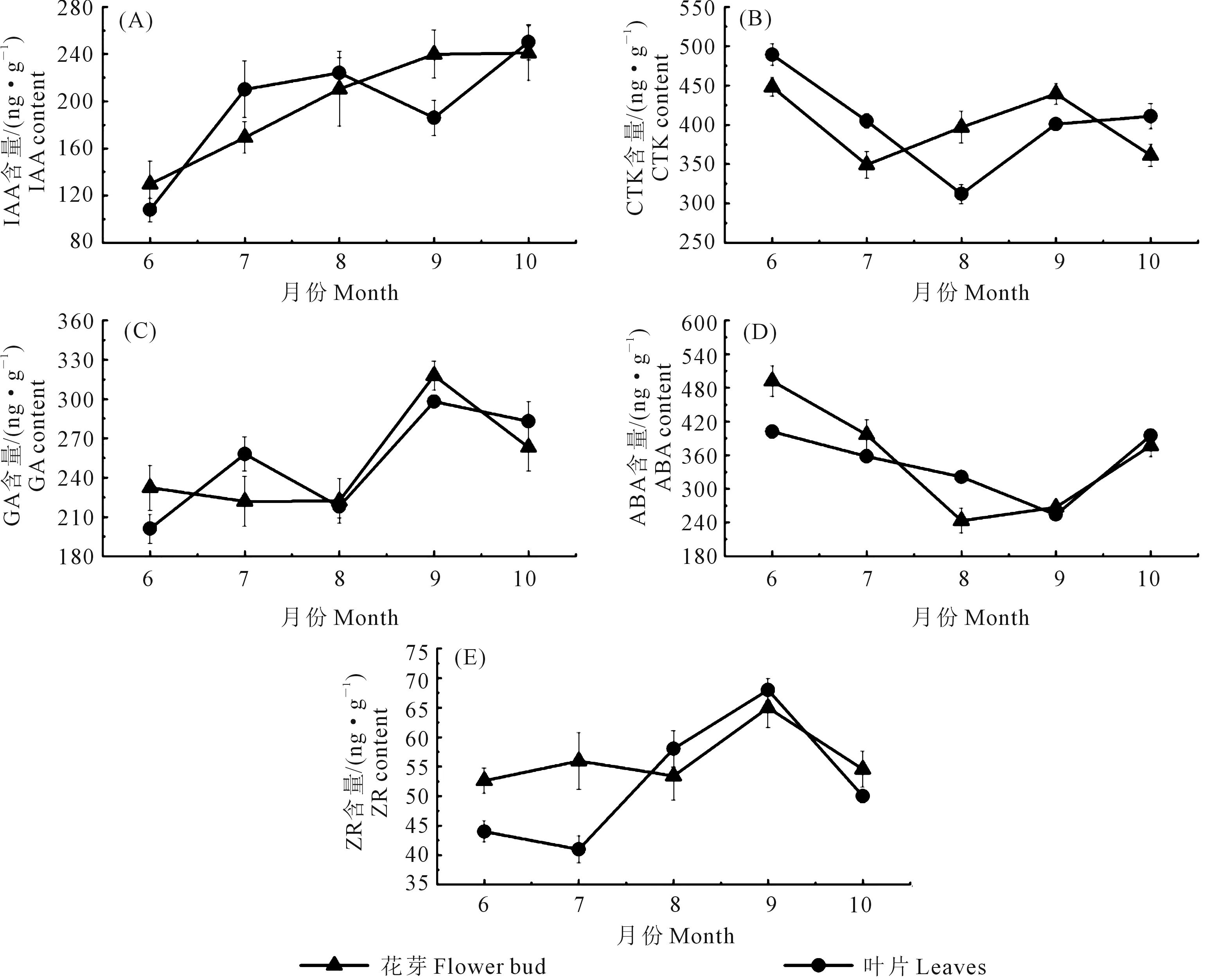
图2 不同时期‘李光杏’内源激素含量的变化Fig.2 Changes of endogenous hormones in Li-Guang Apricot at different periods
叶片中IAA含量6月份最低(108 ng·g-1),9月份显著高于花芽的;CTK变化呈先降后升趋势,分化前期叶片中略高于花芽,进入分化盛期显著低于花芽(8月份为312 ng·g-1);GA和ABA变化趋势与花芽的无显著差异;ZR含量在分化前期低于花芽(6、7月份),之后逐渐升高,9月份达最高值(68 ng·g-1)。
2.3 ‘李光杏’花芽形态分化期不同激素比值的变化
内源激素对花芽分化的调控取决于不同激素比值的变化。由图3可以看出,6月份(未分化期)ABA/IAA、ABA/GA、ZR/IAA比值显著高于其他时期,大多数花芽进入雌蕊分化期(9月)后逐渐下降,分别为ABA/IAA(1.11)、ABA/GA (0.84)、ZR/IAA (0.25),10月份之后(花粉细胞期)逐渐上升,此时大多数花芽形态分化已经完成。ZR/GA的变化呈先升后降趋势,7月份出现最高值(0.25),9月份(雌蕊分化期)最小,为0.21。GA/IAA在6月份最高(1.79),进入形态分化期后逐渐下降。ZR/ABA的变化呈先升后降趋势,9月份最高(0.24),此时大多数花芽处于雌蕊分花期,之后逐渐下降。由图2可知,6月份ABA、CTK这两大促花激素的含量显著升高,但IAA、GA处于较低水平,CTK、ZR、ABA与IAA、GA、比值的高低造成生理分化,然后进入形态分化。

图3 不同时期‘李光杏’各激素比值的变化Fig.3 Changes of hormone ratios of Li-Guang Apricot at different periods
叶片中ABA/IAA、GA/IAA和ZR/ABA的变化趋势与花芽中的大致相同,但ZR/IAA的变化在7月份最低,显著低于花芽中的含量。ABA/GA和ZR/GA的变化与花芽的无显著差异。这些结果表明,‘李光杏’成花诱导期需要较高的ABA/IAA、ABA/GA及ZR/IAA比值,花芽分化完成后则比值降低,这一变化有利于营养生长向生殖生长转变。
2.4 ‘李光杏’花芽形态分化时期全碳、全氮及C/N的变化
植物C/N值反映植物的营养状况,果树体内氮和碳的比例适当,供应充足,花芽分化旺盛,有利于花芽分化的进行。由表1可知,2017年全氮含量在8月份最低(7.42 g·kg-1),全碳含量的变化呈先升后降趋势,且C/N比最高为59.43(8月)。2018年全氮含量呈先降后升趋势,8月初最低(8.15 g·kg-1),之后显著上升,10月份花芽形态分化完成,达到12.66 g·kg-1;全碳含量变化呈先降后升趋势(2018),7月份最低,为442.11 g·kg-1,之后逐渐上升;C/N值变化呈先升后降的趋势,8月份达到最高值,为54.79,之后显著下降,10月份为37.31。碳氮代谢作为植株最基本的代谢过程,对植物生长发育和作物产量形成至关重要,碳氮平衡即C/N反映了植物碳氮代谢的相对强弱,其协调程度,对果树的花芽分化及生长发育具有极其重要的调节作用。
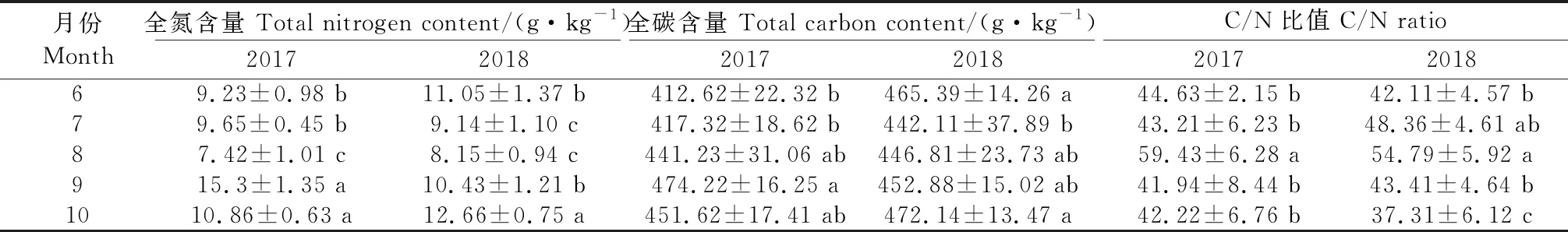
表1 不同时期‘李光杏’全氮、全碳及碳氮比值的变化
3 讨 论
3.1 ‘李光杏’花芽分化时期的确定
花芽分化是一个复杂的过程,生理分化完成后,植株受体内激素和外界条件影响,叶原基的物质代谢及生长点组织形态开始发生变化,逐渐分化出花芽和叶芽,进入形态分化期,并逐渐发育形成花萼、花瓣、雄蕊、雌蕊,从而完成整个花器官的发育[20]。本试验中,‘李光杏’分化初期最早出现在6月20日,而8月10日左右仍有少数花芽处在分化初期;8月初至9月中旬达到分化高峰期,9月中旬后,花芽内花瓣、萼片等器官继续增大,雌蕊原基和雄蕊原基等组织进一步分化;10月后,花粉囊壁内出现花粉母细胞,且排列紧密,呈多角形,整个花蕾体积不断增大,同时也可看到膨大的子房体内部珠心原始体的分化。这些结果说明‘李光杏’花芽分化延续时间长,各时期均有重叠现象,不同分化期均有分化高峰期。王进[21]等认为花芽形态分化期受多种因素(如温度、光周期、地理、品种)的影响,使各阶段出现时间的早晚及持续期的长短略有差异。这也正是‘李光杏’花粉母细胞分化期、花粉粒形成期较长的原因之一。
3.2 内源激素含量变化与花芽分化的关系
植物内源激素是果树花芽分化的关键诱导因子之一,对花芽分化的顺利进行起着重要的调控作用。目前,激素调节成花的观点颇受关注,研究报道也很多,5大类激素对花芽分化的单独作用已基本清楚[22]。但由于内源激素在植物体内分布范围广、含量低,且容易氧化分解以及取材的时期不同,所得结果仍有许多不足之处。大量研究表明,CTK能够激活花芽细胞分裂以促进花的发育[23]。本试验中,CTK含量在分化初期(7月)最低,而分化盛期(8—9月)显著升高,花芽分化完成后逐渐下降,说明CTK对花芽分化有促进作用。这与Luckwill在苹果花芽分化中的研究一致[24]。ZR具有促进细胞分裂和芽的分化、延迟休眠、打破种子休眠等功能,研究发现,较高水平的ZR含量有利于成花植株的花芽分化[25]。本研究表明,ZR的变化在6—8月无较大的波动,而9月份显著升高,此时期正值雌蕊分化期,说明ZR升高有利于‘李光杏’花芽分化的进行。这与前人的研究结果相似。ABA是一种促进成花的内源激素,Garcia等[26]在油桃花芽分化的研究中发现,生理分化期ABA含量逐渐上升,生理分化完成进入形态分化期后,ABA含量逐渐下降,在分化末期再上升。‘李光杏’在6月份(未分化期)ABA含量显著升高,进入形态分化期以后,ABA含量显著下降(7—8月),但分化末期再上升(10月),说明ABA对果树成花的有利作用可能是引起枝条停长,从而使CTK、碳水化合物积累,使分化组织有一个适宜的生长速度,促进成花。
内源GA是果树成花的抑制因素,果实中发育着的种子是GA的重要来源,但这种抑制不仅仅决定于种子内形成的GA含量,还要视其向外扩散的量、扩散的时期及GA的种类。研究发现 GA 的合成受到吲哚乙酸(IAA)的调控,在多年生植物中IAA与GA有着明显的互作关系[27]。IAA是影响植物成花的重要激素之一,果树要顺利进行花芽分化,需降低营养芽内的IAA含量。Stone等[28]指出,降低IAA含量能够减少糖的消耗,增加淀粉的积累,为成花做准备。花芽诱导完成后,由于花芽分化的不可逆转性,此时IAA已不会影响花芽分化完成的数量。郭文丹等[29]在油桐花芽分化过程中测定了其内源激素的动态变化,发现花芽生理分化期IAA含量迅速下降,到花芽形态分化开始前降到较低水平;花芽形态分化开始后,IAA含量呈现出前期上升、中期波动、末期上升的变化规律,说明低含量IAA有利于促进油桐花芽分化。本研究表明,GA含量的变化在6—8月处于较低水平,9月份达峰值,之后显著下降。IAA含量在未分化期(6月)最低,之后逐渐上升,9月之后无显著变化,说明较低的IAA、GA对李光杏花芽分化由生理分化转变为形态分化有着重要作用,同时形态分化期开始后两种激素水平升高并在一定范围内的波动可促进花芽形态分化。
3.3 内源激素平衡与花芽分化的关系
内源激素对花芽分化的调控并不决定于单一的激素,而是依赖于不同激素的动态平衡,即不同激素的比值变化。激素平衡不仅较真实地反映了成花机制,也能解释一些花芽分化的现象。ABA、ZR、CTK这三大促花激素的显著升高,以及它们与IAA、GA比值的升高造成生理分化,然后进入形态分化[30]。果树花芽分化取决于来自短枝叶片的促花因素(CTK,ABA,ZR)和来自果实种子的抑花因素(GA,IAA)之间的平衡关系[31]。李志平等[30]在板栗梨的研究中发现,各时期花芽中ZR/IAA、CTK/GA、ABA/GA比值增高,这些对促进花芽分化都是有利的。本试验研究结果表明,6月份ABA/IAA、ABA/GA、ZR/IAA比值显著高于其他时期,各短枝内其比值越大,花芽孕育数就越多,与前人研究结果相一致。这些结果表明,成花诱导期植物营养体需要一个较高的ABA/IAA、ZR/IAA及ABA/GA的比值,一旦进入花芽孕育期,则比值逐渐降低,这一变化有利于营养生长向生殖生长转变。花芽分化还与 CTK/IAA、ABA/IAA、CTK/GA、IAA/GA、ABA/GA 等平衡有着密切关系,内源激素间的平衡与综合作用促进了‘李光杏’的花芽分化,这对花期的调控具有重要意义。
3.4 碳氮含量与花芽分化的关系
在营养生长向生殖生长转变的过程中内源激素导致的碳水化合物和氮代谢的变化起着重要作用[31]。碳氮比学说认为,果树体内碳和氮的比例适当,供应充足,花芽分化旺盛,开花、结果也多。若碳欠缺,花芽不能形成,氮欠缺,碳相对过剩,虽能形成花芽,但结果不良。本试验结果表明,‘李光杏’花芽形态分化期内C/N的变化呈先升后降趋势,且在花芽分化盛期(8月)显著高于未分化期(6月)。花芽中高的C/N状态,能够达到促花的效应,促进花芽的形成。碳和氮是花芽分化的前提和基础,也是花芽分化的重要营养和能量来源。当C/N高时有利于花芽分化。碳氮比学说为成花生理过程的研究起到了推动作用,C/N协调程度对‘李光杏’花芽分化具有极其重要的调节作用[32-33]。但内源激素的作用方式和C/N的变化与‘李光杏’花芽分化的关系十分复杂,本试验只研究了‘李光杏’分化过程中的生理变化(内源激素和碳氮含量等物质),但尚未探讨内含物质变化的内因,这一系列的变化中基因的调节十分复杂,有关影响李光杏花芽分化的调节机制还有待进一步研究。
4 结 论
‘李光杏’花芽形态分化期因受各种因素的影响,各个分化期出现的时间早晚及持续长短都略有差异。在6月份(未分化期),ABA、CTK这两大促花激素的含量显著升高(分别为ABA 491 ng·g-1,CTK 448 ng·g-1),有利于花原基的形成。ABA/IAA、ZR/IAA及ABA/GA的比值升高(分别为3.75,0.41,1.94)有利于‘李光杏’花芽形态分化的进行。碳氮比升高(37.31~54.79)时,也有利于‘李光杏’花器官的发育。
猜你喜欢
农业工程学报(2022年8期)2022-08-08
作物学报(2022年9期)2022-07-18
扬子江诗刊(2022年1期)2022-01-10
金山(2021年10期)2021-11-02
中国食用菌(2021年3期)2021-05-08
植物研究(2021年1期)2021-02-26
青年文学家(2020年22期)2020-08-31
植物研究(2020年6期)2020-03-05
热带生物学报(2019年1期)2019-04-18
安徽理工大学学报·社会科学版(2014年6期)2015-02-06
