
外源ATP对盐胁迫下油菜种子萌发及幼苗生长的影响
2020-07-28 03:22:20梁娟红李巧丽张小花张腾国
干旱地区农业研究 2020年3期
梁娟红,李巧丽,赖 晶,张小花,张腾国
(西北师范大学生命科学学院,甘肃 兰州 730070)
5’-三磷酸腺苷(adenosine-5’-triphosphate, ATP)作为一种高能化合物,是生物体内进行正常物质代谢和能量代谢的直接能量来源。已有研究发现,细胞外基质中也存在ATP,即胞外ATP(extracellular ATP,eATP),它可以作为一种重要的信使分子,通过特定的细胞信号转导机制参与植物对生物、非生物胁迫的响应过程[1]。Luattge等[2]研究发现施加ATP能够促进燕麦叶片维管束鞘细胞对K+的吸收。此外施加ATP还增强了核酸内切酶的活性[3]。也有研究表明:增加细胞外ATP浓度会抑制拟南芥根的向重力性,跨膜ATP浓度会破坏生长素极性运输[4],导致花粉不萌发以及抑制花粉管生长[5]。另外,外源ATP还参与次生代谢物的积累[6]、细胞凋亡[7]、胁迫抗性[8]等生理过程。但关于外源ATP对种子萌发及抗盐性的影响鲜见报道。
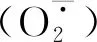
植物液泡为储存Na+提供了一个比较大的空间,液泡膜中的钠-氢交换蛋白(NHX)可以将细胞质中的Na+螯合到液泡中,降低细胞质中Na+浓度,减少过量Na+对植物的毒害[12]。第一个NHX转运体基因是在拟南芥中克隆并命名为AtNHX1[13]。研究发现,NHX也会影响植物体内K+的区室化。过表达AtNHX1基因的番茄在盐胁迫条件下促进了K+从地下向地上运输,液泡中K+浓度上升,细胞间Na+/K+降低,从而有效降低盐胁迫对过表达番茄的盐害作用[14]。Zhao等[15]在两种耐盐性不同的杨树中研究发现,外源ATP的施用显著降低了两种杨树中盐诱导的K+外流,触发盐敏感植株中的SOS(盐超敏感信号转导途径)信号级联反应,并有助于盐度下Na+稳态的控制。除此之外,在植物响应盐胁迫过程中SOS信号途径也发挥着重要的作用,其中SOS1是参与Na+从细胞质向质外体转运的关键基因[16],在细胞膜上编码Na+/H+逆向转运蛋白,在种子萌发过程中起重要作用,处于盐胁迫条件下也能帮助植物更好地生长发育[17]。Jeter等[18]研究发现,在拟南芥中,ATP的外源处理可以激活MAP激酶途径以及参与乙烯生物合成和信号转导蛋白的转录物水平。同时,MAP激酶级联途径也参与抗盐调控过程。有研究表明,MEKK1-MKK2-MPK4/MPK6级联途径可以参与盐胁迫下拟南芥的信号响应过程[19],而MKK9-MPK3/MPK6级联途径参与调节乙烯的合成并在拟南芥对盐胁迫的响应中发挥作用[20]。
随着冬油菜北移技术的成功示范,白菜型冬油菜(Brassicacampestris)已成为北方寒旱区主要的油料作物和生态作物[21]。但北方土壤广泛存在不同程度的盐渍化,严重影响了冬油菜的生长发育和高产潜力的发挥[22]。因此,本文以白菜型冬油菜‘陇油8号’为材料,研究外源ATP浸种对NaCl胁迫处理下油菜种子萌发、幼苗生理及相关基因表达(MAPK3和MAPK6、SOS1和NHX1)的影响,为提高油菜抗盐性提供理论依据。
1 材料与方法
1.1 试验材料处理及培养
挑选籽粒饱满、大小均一的‘陇油8号’种子(由甘肃农业大学农学院提供)用自来水反复冲洗,室温下浸泡2 h,再进行以下处理。(1)不同浓度的外源ATP溶液浸种处理:将种子分别置于浓度为0、1、10、25、50、100 μmol·L-1的外源ATP溶液中,于(25±1)℃培养箱黑暗浸种12 h;浸种后的种子用0.5%的次氯酸钠消毒5 min,无菌水冲洗5~6次后接种到1/2 MS固体培养基中,每个培养皿中放置30粒种子,置于(25±1)℃、光照强度为250 μmol·(m2·s)-1、光照周期16 h/8 h(光照/黑暗)的培养箱(RDN型人工气候箱)中培养。每个处理5个重复。(2)CK:将油菜种子置于1/2 MS固体培养基中,培养条件与(1)处理相同。(3) NaCl单独处理:将油菜种子置于含200 mmol·L-1NaCl的1/2 MS固体培养基中,培养条件与(1)处理相同。(4)ATP+NaCl复合处理:用25 μmol·L-1外源ATP溶液,按照(1)处理的方法浸种。用上述方法接种于含200 mmol·L-1NaCl的1/2 MS固体培养基中,培养条件与(1)处理相同。
1.2 种子萌发相关指标的测定
培养3 d后观察种子萌发情况,以胚根凸出种皮长度大于等于种子长度作为发芽标志,计算发芽势。处理7 d后观察种子发芽情况,再以上述方法观察种子的萌发情况,计算发芽率。
1.3 生理指标的测定
取培养7 d的油菜幼苗地上部分进行后续试验。幼苗根长与株高测定参照田梦雨[23]的方法;过氧化氢(H2O2)含量测定参照Yin等[24]的方法;羟自由基(·OH)含量测定参照Halliwell[25]的方法;丙二醛(MDA)含量测定参照硫代巴比妥酸法[26];相对电导率测定参照电导率仪法[27];游离脯氨酸含量测定参照磺基水杨酸法[28];可溶性糖含量测定参照蒽酮显色法[29]。
1.4 活性胶电泳及染色
活性胶制备参照Lammli[30]的方法。采用5%的浓缩胶和12%的分离胶。CAT上样量为6 μL,POD上样量为9 μL。浓缩胶电压为80 V,分离胶电压为120 V,冰浴中稳压电泳约5~6 h。
CAT活性染色参照 Woodbury等[31]的方法;POD同工酶活性胶染色参照黄永芬等[32]的醋酸联苯胺方法。
1.5 实时荧光定量PCR
油菜幼苗总RNA的提取采用TaKaRa MiniBEST Plant RNA Extraction Kit试剂盒,参照试剂盒实验步骤操作。以提取的总RNA为模板,参照反转录试剂盒合成cDNA。以Actin-F、Actin-R为管家基因引物,MAPK3-F和MAPK3-R为MAPK3引物,MAPK6-F和MAPK6-R为MAPK6引物,SOS1-F和SOS1-R为SOS1引物,NHX1-F和NHX1-R为NHX1引物(表1),按照荧光定量PCR试剂盒SYBR Premix Ex TaqTM(购自TaKaRa公司)的操作步骤用25 μL体系进行扩增检测。采用2-ΔΔCt方法分析数据,计算基因的相对表达量。

表1 PCR引物序列
1.6 数据处理
试验数据使用SPSS 20.0软件进行统计分析,用Excel 2010软件作图。
2 结果与分析
2.1 不同浓度外源ATP浸种对油菜种子发芽率和发芽势的影响
由表2可知,与CK相比较,发芽势和发芽率均随外源ATP浓度增大呈先升后降的趋势。当外源ATP浓度为25 μmol·L-1时,发芽势和发芽率达到最大值,分别比CK提高26.9%和17.2%。结果表明,外源ATP浓度在一定范围内对种子萌发具有促进作用,而25 μmol·L-1的外源ATP处理效果最明显。

表2 不同浓度外源ATP对油菜种子发芽率和发芽势的影响
2.2 外源ATP浸种对NaCl胁迫处理下油菜种子发芽率和发芽势的影响
如表3所示,与CK相比,200 mmol·L-1NaCl胁迫处理显著抑制了油菜种子萌发,发芽率、发芽势分别下降了13.5%、4.7%;ATP+NaCl复合处理下的种子萌发情况显著优于NaCl单独处理,其发芽率、发芽势分别提高了13.7%和15.0%。结果表明,NaCl胁迫处理显著抑制了油菜种子萌发,而ATP浸种能显著缓解NaCl胁迫对油菜种子萌发的抑制作用。

表3 外源ATP 浸种对NaCl胁迫处理下 油菜种子萌发的影响
2.3 外源ATP浸种对NaCl胁迫处理下油菜幼苗生物量的影响
如表4所示,200 mmol·L-1NaCl胁迫处理下的油菜幼苗根长、株高及鲜重均低于CK且鲜重显著下降;ATP+NaCl复合处理下的油菜幼苗根长、株高及鲜重相较于NaCl单独处理分别升高了23.6%、27.3%、28.6%。结果表明,NaCl胁迫处理能抑制油菜幼苗生长和生物量积累,外源ATP浸种能够缓解NaCl胁迫处理对油菜幼苗的抑制作用。
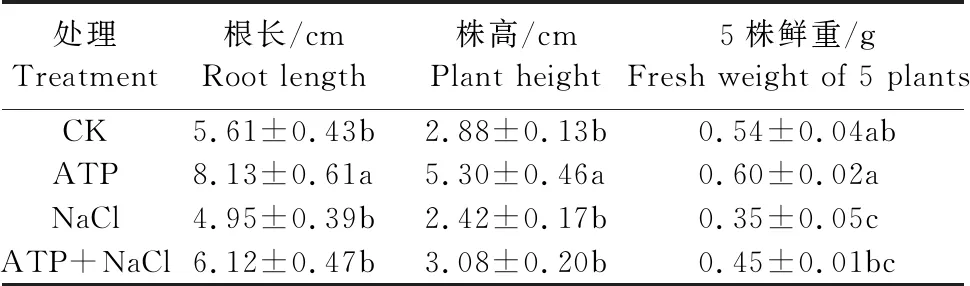
表4 外源ATP浸种对NaCl胁迫处理下 油菜幼苗生物量的影响
2.4 外源ATP浸种对NaCl胁迫处理下油菜幼苗生理特性的影响
2.4.1 外源ATP浸种对NaCl胁迫处理下油菜幼苗H2O2和·OH含量的影响 如图1显示,与CK相比,外源ATP浸种、NaCl胁迫处理均显著诱导油菜幼苗中H2O2(图1A)和·OH(图1B)的产生;ATP+NaCl复合处理与NaCl单独胁迫处理相比,H2O2和·OH的含量分别降低了13.6%和6.3%。结果表明,NaCl胁迫处理显著促进了油菜幼苗中ROS的积累,而外源ATP浸种能够降低NaCl胁迫处理下油菜中活性氧(ROS)含量,缓解盐胁迫对油菜幼苗的损伤。
2.4.2 外源ATP浸种对NaCl胁迫处理下油菜幼苗CAT和POD活性的影响 由图2A可知,不同处理下CAT同工酶条带数相同,有且仅有一条带,在外源ATP浸种、NaCl单独胁迫处理下,油菜幼苗CAT酶带相对灰度比CK分别高3.0%和16.0%;ATP+NaCl处理下的CAT酶带相对灰度比NaCl单独胁迫处理高10.3%。如图2B所示,4个处理下的POD有5条同工酶带,但同一酶带在不同处理下有差异。NaCl胁迫处理下油菜幼苗POD酶带相对灰度比CK高89.0%;ATP+NaCl处理下POD同工酶条带相对灰度比NaCl单独处理高27.0%。结果表明,外源ATP浸种、NaCl胁迫处理都会提高CAT、POD酶活性水平;外源ATP浸种进一步提高了NaCl胁迫下油菜幼苗中的CAT、POD酶活性,增强了油菜幼苗对体内过量ROS的清除能力。

图2 盐胁迫处理下外源ATP浸种的油菜幼苗CAT(A)和POD(B)电泳图及相对灰度分析Fig. 2 Electrophoresis and relative grayscale analysis of antioxidant enzymes in Brassica campestris seedling under NaCl stress by exogenous ATP seed soaking

注:不同小写字母表示处理间差异显著(P<0.05)。下同。Note: Different lowercase letters in theFigure indicate significant differences among treatments (P<0.05). The same below.图1 NaCl胁迫处理下外源ATP浸种的油菜幼苗中H2O2 (A)和·OH (B)含量Fig.1 H2O2 (A) and ·OH (B) accumulation of Brassica campestris seedling exposed under NaCl stress by exogenous ATP seed soaking
2.4.3 外源ATP浸种对NaCl胁迫处理下油菜幼苗膜损伤程度的影响 逆境条件下,MDA和相对电导率能够作为细胞损伤程度的指标。从图3可以看出,与CK相比,NaCl胁迫显著增加了油菜中的MDA含量(图3A)和相对电导率(图3B),表明NaCl胁迫可以加剧油菜幼苗的膜脂过氧化程度。外源ATP浸种显著降低了NaCl胁迫处理下的MDA含量和相对电导率,MDA含量降低了27.6%,相对电导率降低了20.5%。结果表明,外源ATP浸种可以降低NaCl胁迫处理下油菜中的MDA含量和相对电导率,减少膜脂过氧化程度和电解质的渗漏,保护细胞膜的完整性,减轻盐胁迫对油菜的损伤。

图3 外源ATP浸种对NaCl胁迫处理下油菜幼苗中丙二醛(A)和相对电导率(B)的影响Fig.3 Effects of seed soaking with exogenous ATP on malondialdehyde (A) and relative conductivity (B) of Brassica campestris seedling under NaCl stress
2.4.4 外源ATP浸种对NaCl胁迫处理下油菜渗透调节物质的影响 可溶性糖和脯氨酸是细胞内重要的渗透调节物质,可保护细胞内蛋白分子,稳定生物大分子结构。如图4可见,NaCl胁迫处理、ATP+NaCl复合处理都显著增加了油菜幼苗中可溶性糖(图4A)和脯氨酸(图4B)含量,NaCl胁迫处理下可溶性糖、脯氨酸含量分别达到CK的187.8%和459.5%;ATP+NaCl复合处理下油菜幼苗中可溶性糖和脯氨酸含量分别达到NaCl胁迫处理的15.9%和41.0%。结果表明,NaCl胁迫处理下外源ATP浸种能显著增加油菜幼苗中渗透调节物质积累,增强细胞的持水能力,维持细胞的正常生理代谢,从而增强油菜幼苗的耐盐性。

图4 外源ATP浸种对NaCl胁迫处理下油菜幼苗中可溶性糖(A)和脯氨酸(B)的影响Fig.4 Effects of seed soaking with exogenous ATP on soluble sugar (A) and proline (B) content of Brassica campestris seedling under NaCl stress
2.5 外源ATP浸种对NaCl胁迫处理下油菜幼苗相关基因表达的影响
2.5.1 外源ATP浸种对NaCl胁迫处理下油菜幼苗MAPK3/MAPK6基因表达的影响 如图5所示,外源ATP处理下的油菜幼苗中MAPK3(图5A)和MAPK6(图5B)基因表达量分别较CK增加16.0%、7.0%。在NaCl单独胁迫处理下的MAPK3和MAPK6基因表达量分别较CK增加86.0%和69.0%。ATP+NaCl处理下的MAPK3和MAPK6基因表达量分别较NaCl单独胁迫处理增加75.3%、146.2%。结果表明,油菜幼苗中MAPK3和MAPK6基因的表达水平受到NaCl胁迫处理的诱导,外源ATP浸种增强了NaCl胁迫处理下油菜幼苗中MAPK3和MAPK6基因的表达。
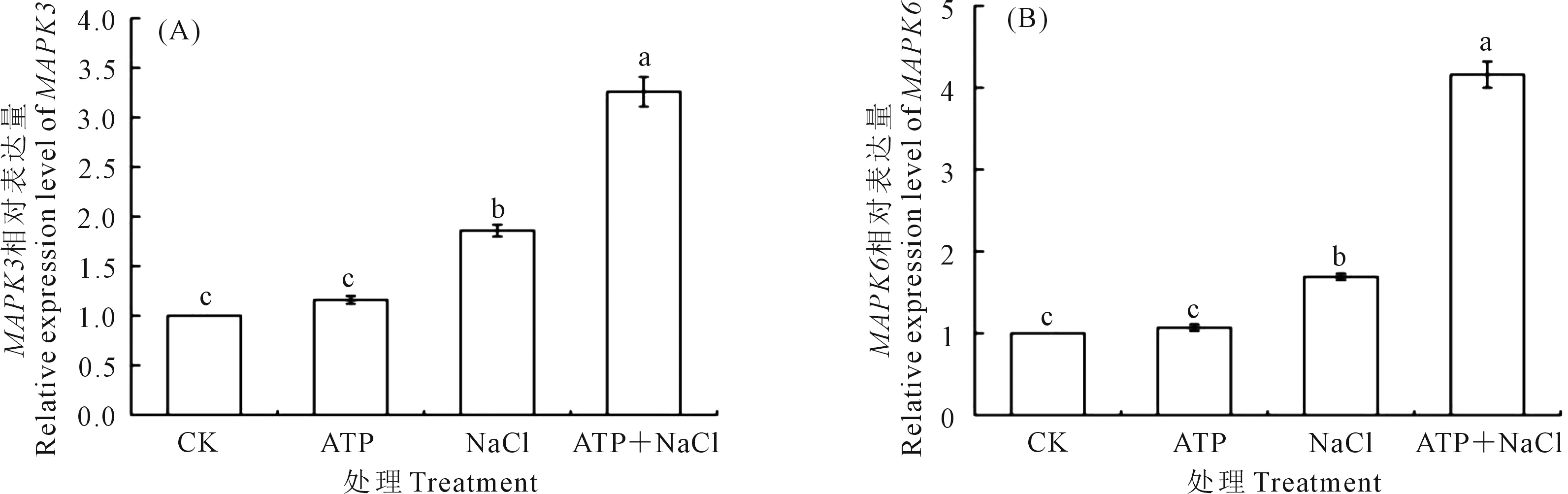
图5 外源ATP浸种对NaCl胁迫处理下油菜幼苗中MAPK3(A)和MAPK6(B)基因表达的影响Fig.5 Effects of seed soaking with exogenous ATP on the expression of MAPK3 (A) and MAPK6 (B) genes in Brassica campestris seedling under NaCl stress
2.5.2 外源ATP浸种对NaCl胁迫处理下油菜幼苗SOS1/NHX1基因表达的影响 如图6所示,NaCl胁迫处理下的油菜幼苗中NHX1(图6A)和SOS1(图6B)基因表达量上调,分别是CK的1.41倍和1.34倍。与NaCl单独胁迫处理相比,ATP+NaCl处理下的油菜幼苗中NHX1和SOS1基因表达量分别上升了17.0%和66.4%。结果表明:油菜幼苗中NHX1和SOS1基因的表达受到NaCl胁迫处理的诱导,外源ATP浸种进一步提高了NaCl胁迫处理下油菜幼苗中NHX1和SOS1基因的表达。

图6 外源ATP浸种对NaCl胁迫处理下油菜幼苗NHX1(A)和SOS1(B)基因表达的影响Fig.6 Effects of seed soaking with exogenous ATP on the expression of NHX1 (A) and SOS1 (B) genes in Brassica campestris seedling under NaCl stress
3 讨论与结论
盐胁迫对植物的危害包含离子胁迫和渗透胁迫。斯琴巴特尔等[33]研究玉米(Zeamays)种子萌发时发现盐胁迫抑制了玉米种子的发芽。孙君艳等[34]研究盐胁迫对小麦种子萌发及幼苗生长的影响,发现高浓度盐处理对小麦种子萌发和幼苗生长起抑制作用。Chivasa等[35]研究结果表明:细胞外ATP可能通过直接或间接的方式与细胞膜外的某种蛋白质结合,从而在胁迫条件下提高细胞生存率。本试验结果发现,200 mmol·L-1NaCl处理对油菜种子萌发有抑制作用,而25 μmol·L-1的外源ATP对油菜种子萌发有显著的促进作用,并能够提高油菜幼苗的生物量积累,外源ATP浸种能缓解NaCl处理对油菜种子萌发的抑制作用。

植物在遭受盐胁迫时会触发一系列抗盐相关基因的表达,如NHX和SOS基因的表达。液泡型Na+/H+逆向转运蛋白(NHX1)可以将细胞质中的Na+逆Na+浓度梯度转运到液泡中进行区室化,质膜型Na+/H+逆向转运蛋白(SOS1)也可以将Na+排到胞外,防止细胞质中的细胞器受到Na+的危害。过表达OsNHX1转基因水稻细胞在盐条件下存活较好,生长速度和总Na+含量显著高于野生型[38]。本研究发现,油菜幼苗中NHX1、SOS1基因的表达受到盐胁迫的诱导,而外源ATP浸种提高了盐胁迫处理下油菜幼苗中NHX1、SOS1基因的表达,NHX1和SOS1转录水平的提高,预示着外源ATP处理触发了NHX基因家族和SOS途径介导的Na+外排。MAPK信号级联途径在植物应对盐、干旱、寒冷、炎热和受伤等信号过程中发挥着十分重要的作用,主要是MAPK3、MAPK4和MAPK6被快速激活[39]。Teige等[20]研究发现,冷和盐胁迫激活拟南芥的MKK2,控制冷应答基因(COR)的表达,调控植物耐冷和耐盐能力。而且,MKK2上游的MEKK1和下游的MPK4和MPK6都能被各种胁迫所激活。在本研究结果中,油菜幼苗中MAPK3、MAPK6基因的表达受到盐胁迫的诱导,而外源ATP浸种提高了盐胁迫处理下油菜幼苗中MAPK3、MAPK6基因的表达。以上这些研究结果表明,外源ATP在调节植物响应盐胁迫的过程中,盐超敏感 (SOS)通路以及MAPK激酶途径等多种信号传导途径之间可能存在交叉对话。
综上所述,外源ATP浸种通过激活ROS清除系统来有效地清除过量的ROS,降低NaCl胁迫处理下油菜幼苗膜脂过氧化程度,促进渗透调节物质的积累及抗盐胁迫相关基因的表达,有效保证油菜幼苗代谢活动的正常进行,从而提高油菜幼苗的抗盐性。
猜你喜欢
舰船科学技术(2022年11期)2022-07-15 07:51:56
现代园艺(2020年19期)2020-10-02 03:15:48
西藏农业科技(2019年3期)2019-11-04 00:35:10
种子(2019年6期)2019-07-13 07:45:16
种子(2019年4期)2019-05-28 02:04:14
现代农村科技(2019年4期)2019-01-06 14:13:19
现代园艺(2018年3期)2018-02-10 05:18:12
上海农业学报(2017年3期)2017-04-10 12:39:12
中学生数理化·八年级数学华师大版(2008年3期)2008-08-26 11:26:16
现代农业科技(2006年23期)2007-01-05 03:23:54
