
短尾蝮操作性条件反射训练探究
2020-07-27 09:15朴弋戈刘杨唐业忠陈勤
四川动物 2020年4期
朴弋戈,刘杨,唐业忠,陈勤*
(1.中国科学院成都生物研究所,成都610041;2.中国科学院大学,北京100049)
1938年,美国心理学家斯金纳总结并提出了“操作性条件反射”(Skinner,1938)。操作性条件反射是通过将自发行为受刺激强化影响而与刺激相联系,在正性奖励与负性惩罚的作用下,引导实验个体完成特定行为,是动物行为、认知研究中的经典训练方法,在模式动物中产生了丰富的研究成果(Huston &Borbely,1973;Wengeretal.,2004;Brembs,2011;Valenteetal.,2012)。爬行类、鸟类与哺乳类共享一些基本生理与行为特征,又因中枢神经的结构差异,具有认知能力的差别(Macphail,1982)。目前,多数认知研究仅在鸟类与哺乳类中开展,较少涉及爬行类。随着认知科学的发展,对于爬行类的认知研究亦开始起步(Vonk &Shackelford,2012;Wilkinson &Huber,2012;Matsubaraetal.,2017)。然而,爬行类中学习行为研究取得的发展仍较有限,且多集中于红腿陆龟Geochelonecarbonaria、纳尔逊伪龟Pseudemysnelsoni等少数物种或类群(Wilkinson &Huber,2012)。因此,该领域亟待探究更多的物种,在更大的分类尺度上比较多种爬行动物的学习能力(Matsubaraetal.,2017)。
蛇类作为爬行纲Reptilia中种类数量最丰富、分布最广泛的类群之一(Kerkkampetal.,2017),在生物学及诸多交叉领域都有重要的研究价值(Grachevaetal.,2010;Lillywhite,2014;Botheetal.,2019)。蛇类的学习行为研究亦取得了一定的进展(Holtzmanetal.,1999;Stoneetal.,2000;Place,2005;Burghardt,2013;Emeretal.,2015;Friesen,2017),但仍限于少数物种,且较少涉及蝮亚科Crotalinae蛇类。蝮亚科蛇类具有高度灵敏的颊窝器官,可接收环境中的红外信息(Noble &Schmidt,1937;Bullock &Cowles,1952),与经视觉感官接收的光学信息一起形成独特的“视觉”(Goris,2011)。独特的信息输入增强了蝮蛇对环境的感知能力(Kardong &Mackessy,1991;Embaretal.,2018);但额外输入的信息是否影响蝮蛇的行为决策,与其他蛇类或有鳞类相比,学习等认知能力是否存在区别尚不明确。
参考有鳞目Squamata学习行为的既有研究,本研究使用Y迷宫作为实验装置,采用既有研究中曾使用的3种惩罚刺激,以短尾蝮Gloydiusbrevicaudus进行操作性条件反射训练。基于Matsubara等(2017)的研究思路,使用现有的实验设计探究其他类群(蝮亚科)的学习行为,为在更大的分类尺度上进行比较研究提供基础数据。
1 材料与方法
1.1 实验动物
短尾蝮是我国广泛分布的一种蝮亚科蛇类(赵尔宓,2006),个体相对较小,在蝮蛇行为学研究中可操作性强。本实验选取10条实验室饲养的短尾蝮,包括3条雄性和7条雌性(表1)。受进食、蜕皮等情况影响,所有实验均仅选取其中部分个体参与。
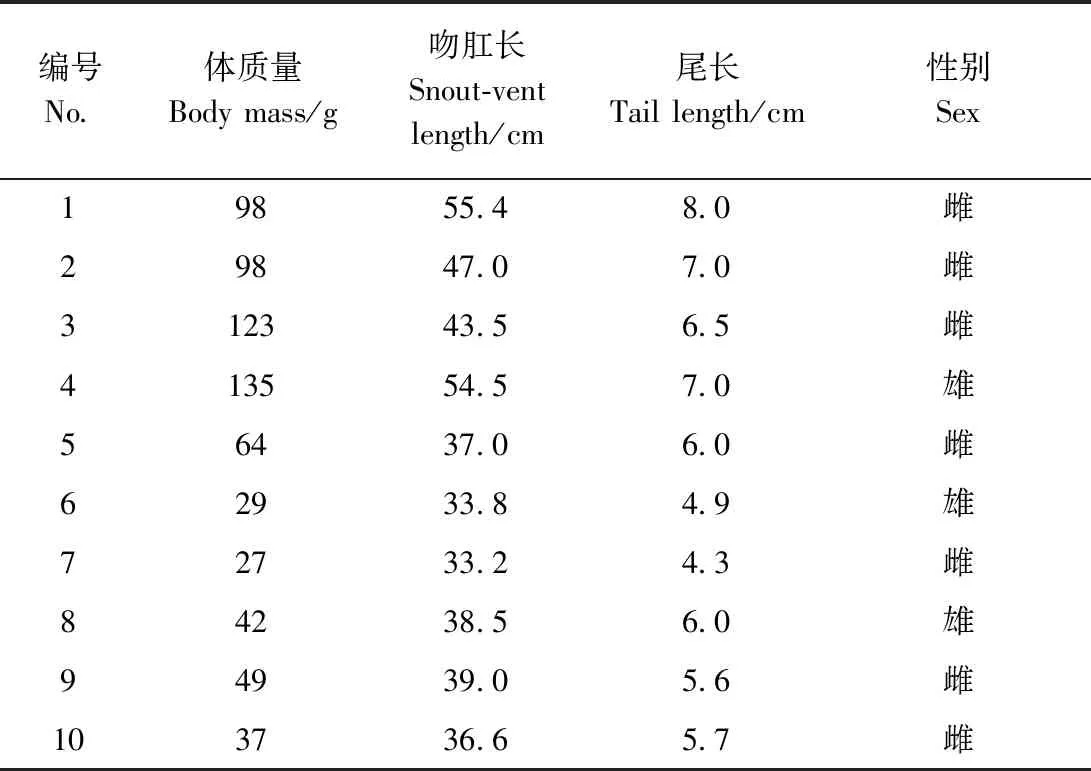
表1 样本形态测量数据Table 1 Morphological measurement of the specimens
实验动物均单独饲养于饲养箱(长32 cm×宽22 cm×高19 cm)中。饲养箱内以报纸作为垫料,放置一水盆供蛇自由饮用,同时提供爬虫躲避穴(长23 cm×宽16.5 cm×高7.5 cm)供蛇隐藏。饲养房内温度设定为25 ℃。光周期设定为12 h∶12 h,光照时间为08∶00—20∶00。使用实验小鼠以1周1次喂食,实验进行期间不提供食物。实验前遮蔽蛇的双眼,使用1层医用胶布后覆盖1层黑色胶布,适应24 h以上。若蛇已进食,则至少2 d后再进行遮蔽。若蛇临近蜕皮,则该个体不参与实验,待其蜕皮完成后进行。
1.2 实验装置
基础Y迷宫使用厚度3 mm黑色亚克力材料制成。迷宫各部分均为长方体,主体尺寸为长30 cm×宽20 cm×高15 cm,两臂尺寸均为长30 cm×宽11 cm×高15 cm,与主体部分相连,夹角约50°。以金属网(长20 cm×宽15 cm,孔径5 mm×5 mm)(Manjarrez &Vacaleón,2018)将主体部分分为适应区(20 cm×20 cm)和选择区(20 cm×10 cm)(图1)。使用规格为1 cm×1 cm的正方形白色电热陶瓷片(XH-RP1010;江苏星河电子科技,江苏宿迁)组装成2个图形(图2),作为两侧迷宫臂的指示物。图形均使用20个陶瓷片串联拼装,保证图形发热面积一致。实验过程中,2个图形串联通电30 V,摆放于迷宫臂尽头,呈现红外信息辅助蝮蛇进行定位、学习。指定“”图形所在一侧的迷宫臂为安全区(无惩罚刺激),“+”图形所在一侧的迷宫臂为惩罚区。实验中迷宫摆放朝向、图形位置与实验动物的实验顺序均随机。实验间隔期间使用肥皂水擦拭整个迷宫并吹干,以减少残留气味因素的干扰。
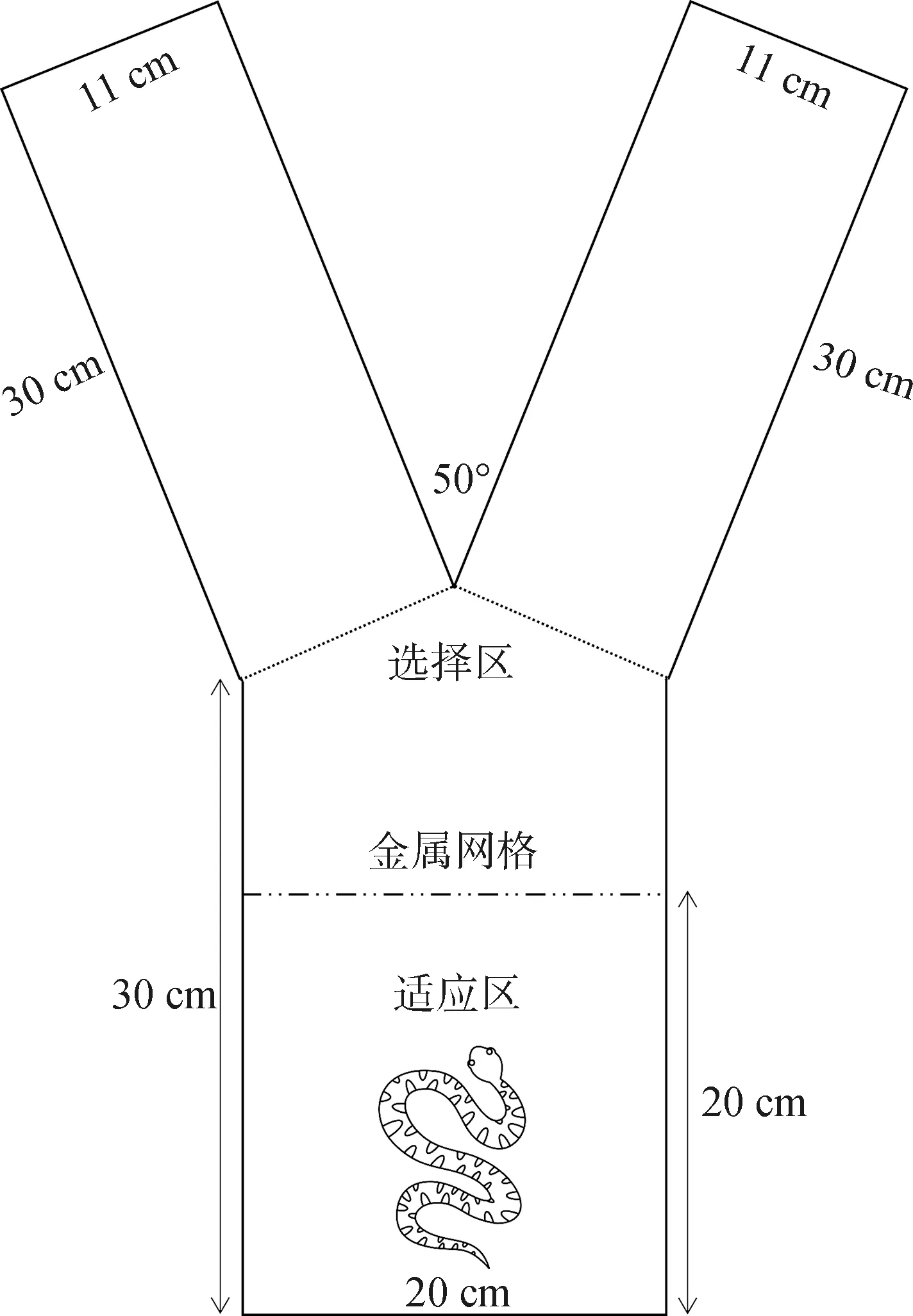
图1 Y迷宫装置示意图Fig.1 Schematic diagram of Y-maze
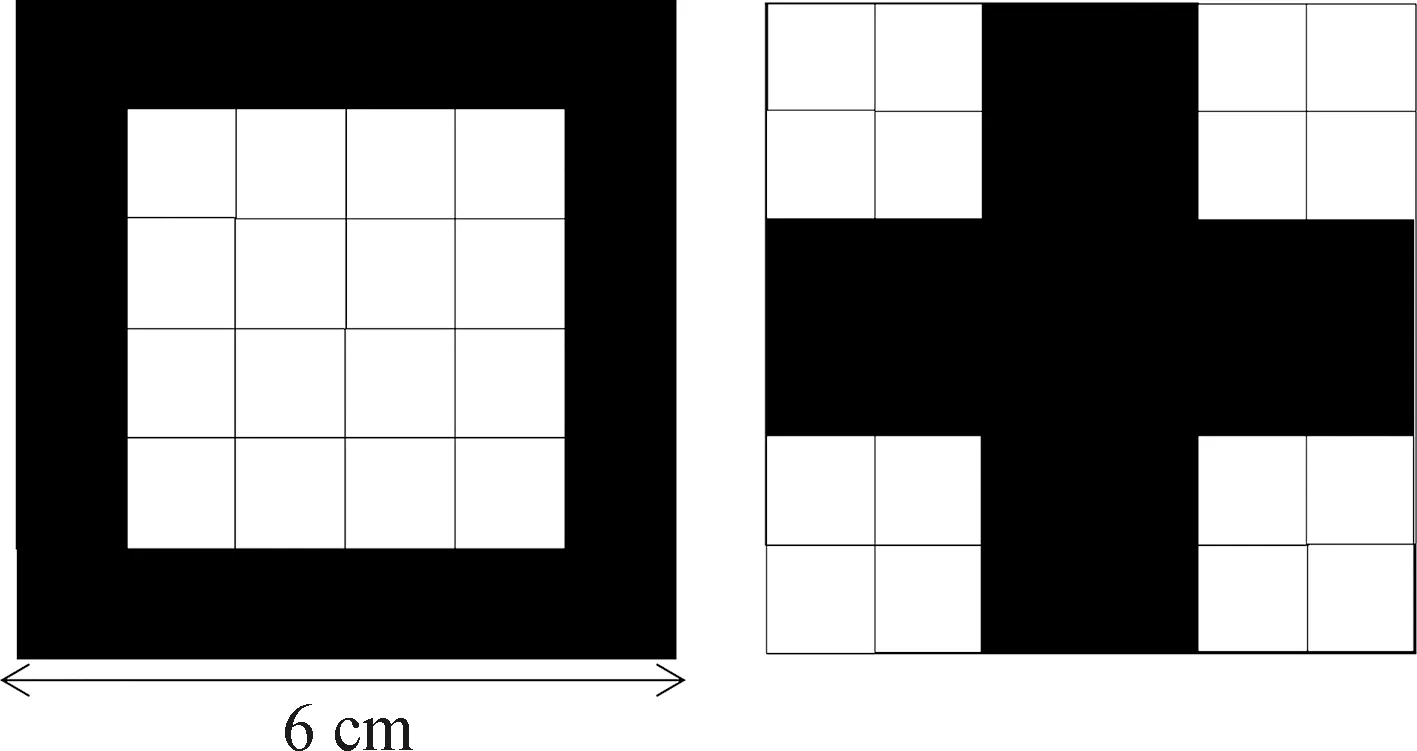
图2 指示图形Fig.2 Indicating shapes
1.3 实验刺激
蛇类进食频率低、耐饥饿能力强,以食物作为奖励难以维持实验定量、定期进行(Kellogg &Pomeroy,1936;Wilkinson &Huber,2012)。此外,蝮蛇作为典型的伏击型捕食者,常长时间蜷缩一处(Clark,2016),游走性较差,故主要使用惩罚刺激驱使其离开迷宫起始区而进入迷宫两臂。
1.3.1 水刺激尽管许多蛇类可入水游泳(赵尔宓,2006),但多数蛇类仍积极离开水体。先前研究中使用半水栖型平原束带蛇Thamnophisradix与水栖型南部水蛇Nerodiafasciata的水迷宫实验均取得了较好的实验结果(Kellogg &Pomeroy,1936;Fuenzalida &Ulrich,1975)。
第一组实验中,迷宫倾斜5°摆放,主体位于斜面下方,两臂位于斜面上方。迷宫底部垫以砂纸,增大摩擦力以辅助蛇爬行。于适应区顶部开一小孔(直径4 mm),以塑料软管连接至水槽。实验时,将短尾蝮引入适应区后关闭金属网,待其适应10 min。适应结束后,打开金属网,计时10 min,允许蛇自由爬行。若计时3 min蛇仍未离开适应区,则将水流自适应区顶部小孔注入,待水淹没至迷宫两臂分叉处后停止注水。该水位迫使蛇离开淹没的适应区,向干燥的斜面上方(即迷宫两臂)爬行。以蛇身体前段三分之一进入某一侧迷宫臂判断为其进入该侧。若蛇进入惩罚区,则用金属蛇钩以3 s/次频率轻压其头部(Morietal.,2016)迫使其离开。若蛇进入安全区,则计时停止,记录其逃出时间,实验结束。若实验计时满10 min蛇仍未进入安全区,则使用蛇钩小心将蛇引入安全区,该时间记录为10 min。
第二组实验中,迷宫摆放于平面,内部注水,水深12 cm。同时撤去金属网,使用尺寸略小于适应区的一金属网笼(长20 cm×宽19 cm×高11 cm,孔径5 mm×5 mm)悬挂于适应区水面上方。实验时,将短尾蝮引入金属网笼,待其适应5 min后,将金属网笼沉入水中,打开网笼计时,允许蛇自由游动,寻找离开水体的出口。惩罚区出口封闭,无法逃出;安全区出口开放,可以逃出。待蛇经出口爬出时计时停止,记录其逃出时间,实验结束。若实验计时满5 min蛇仍未逃出,则使用木棒将蛇引入出口,该时间记录为5 min。
每条蛇每天进行5次实验,实验间隔20 min(Fuenzalida &Ulrich,1975)。
1.3.2 高温刺激已有研究显示颊窝对蝮蛇的体温调节具有意义,保留颊窝的蝮亚科蛇类均可准确进入低温躲避处,遮蔽颊窝的蝮亚科蛇类或无颊窝的其他蝰科Viperidae蛇类则无法完成这一行为(Krochmal &Bakken,2003;Krochmaletal.,2004)。本实验借鉴Krochmal和Bakken(2003)的实验设计:以高温作为惩罚刺激,Y迷宫主体部分加热、维持温度高于40 ℃(该温度为蝮亚科蛇类的胁迫温度),迫使蛇寻找出口逃离。
本实验使用加热垫置于基础迷宫主体下方,持续通电提供高温刺激(图3),迷宫两臂不作加热处理。Y迷宫上方覆以一透明打孔亚克力板防止蛇逃逸。该板厚1 cm,尺寸60 cm×60 cm,打孔排列为28×28(行×列),孔内径9 mm,相邻两孔边缘间距为1 cm。实验时,将蛇引入适应区,适应1 min后,打开金属网并开始计时,任其自然爬行。以蛇身体前段三分之一进入迷宫两臂之一判断为进入该侧。若蛇进入安全区,停止计时,实验结束。若蛇进入惩罚区,则以木棒经上方亚克力板的小孔穿入,轻敲其头部,直至迫使其离开该侧。实验以10 min内蛇进入安全区为止,记录该时间。若超过10 min,蛇仍未进入安全区,则用木棒轻触蛇,引导其进入,该时间记录为10 min。实验结束后,蛇停留于安全区10 min以降低体温。
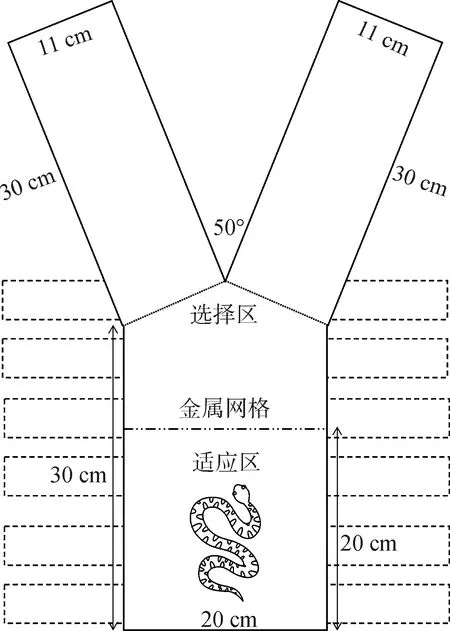
图3 加热Y迷宫装置示意图Fig.3 Schematic diagram of the heating Y-maze
每条蛇每天进行4次实验,实验间隔1 h(Holtzmanetal.,1999;Stoneetal.,2000;Friesen,2017)。
1.3.3 轻敲刺激使用轻触或轻敲的方法在一些蜥蜴的学习行为探究中是一种较为成熟的方法(Amiel &Shine,2012;Sunetal.,2014;Chungetal.,2017;Dayananda &Webb,2017;Kangetal.,2018)。因此,本实验尝试给予轻敲刺激迫使短尾蝮逃离适应区,Y迷宫中仅安全区可免于该惩罚刺激,以训练实验动物持续逃入安全区。每6次实验为一组,设定学习标准为:在一组连续6次实验中出现5次首次选择正确(Chungetal.,2017;Kangetal.,2018),达到该标准的动物被认为基本学会持续逃入安全区域。
参考Lemaster和Mason(2001)将Y迷宫修改为无墙迷宫。撤除迷宫全部垂直面,仅保留原迷宫底面。迷宫放置于金属支架上,抬高距离地面60 cm,以防止实验过程中蛇爬下迷宫。同时,使用双层黑色塑料袋和金属框架制作一个矩形围帐(长72 cm×宽70 cm×高70 cm),实验时将抬高的迷宫置于其中以减少外界环境的干扰。使用热熔胶固定32支塑料短管垂直于迷宫底面,为蛇提供爬行的支点。塑料管直径1 cm、高2 cm,排成1列,塑料管间隔2.5 cm(图4)。实验时,将短尾蝮置于迷宫上,使用一金属网笼(长15 cm×宽10 cm×高11 cm,孔径5 mm×5 mm)将蛇罩住,待其适应5 min。适应结束后,提起金属网笼,允许蛇自由爬行,计时10 min。蛇在爬动过程中不给予刺激;若蛇连续15 s未发生运动,则给予轻敲刺激。使用木棒以1 s/次的频率轻敲蛇的尾基部(泄殖腔背侧),直至其开始运动。以蛇身体前段三分之一进入迷宫臂某一侧判断为其进入该侧。若蛇进入惩罚区,则以木棒轻敲其头部迫使其离开。若蛇进入安全区,计时停止,记录其逃出时间,实验结束。若10 min后蛇仍未进入安全区,则使用蛇钩将蛇轻轻引入安全区域,该时间记录为10 min。
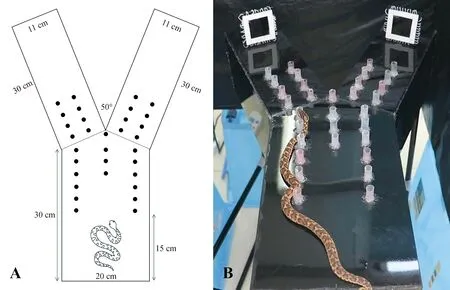
图4 无墙Y迷宫装置示意图(A)与实物图(B)Fig.4 Schematic diagram (A) and photo (B) of the wall-less Y-maze
每条蛇每天进行4次实验,实验间隔1 h。
1.4 数据分析
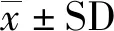
2 结果
水刺激与高温刺激实验以选择正确率与随机选择(50%)存在显著差异为标准,达到该标准的实验动物判断为基本习得训练行为。
2.1 水刺激
第一组实验使用2号、3号与5号共3条个体,进行8 d共计115次有效训练。其中,2号个体在第8天实验开始前受外伤,仅有7 d实验数据。3条个体在训练过程中表现出差异化的行为,但均未达到学习标准;其选择正确率与逃出时间见图5。

图5 第一组水刺激实验选择正确率(A)与逃出时间(B)Fig.5 Choice accuracy (A) and escape time (B) in the water stimulation experiment 1
2号个体正确率始终未达到随机水平(50%),其逃出时间也最长。实际在7 d共35次实验内,该个体在24次实验中停留于适应区水体,未表现出明显移动。3号个体选择正确率在8 d 实验内5次高于50%,但与随机水平之间的差异无统计学意义(P>0.05),其逃出时间在8 d实验内未见明显变化(P>0.05)。5号个体选择正确率在第4天、第5天与第6天逐步提高,但随后降低,正确率与随机水平之间的差异无统计学意义(P>0.05),其逃出时间在8 d实验内无显著变化(P>0.05)。
第二组实验使用3号与4号共2条个体,进行9 d共88次有效训练,但2条个体均未达到学习标准。个体选择正确率与逃出时间见图6。
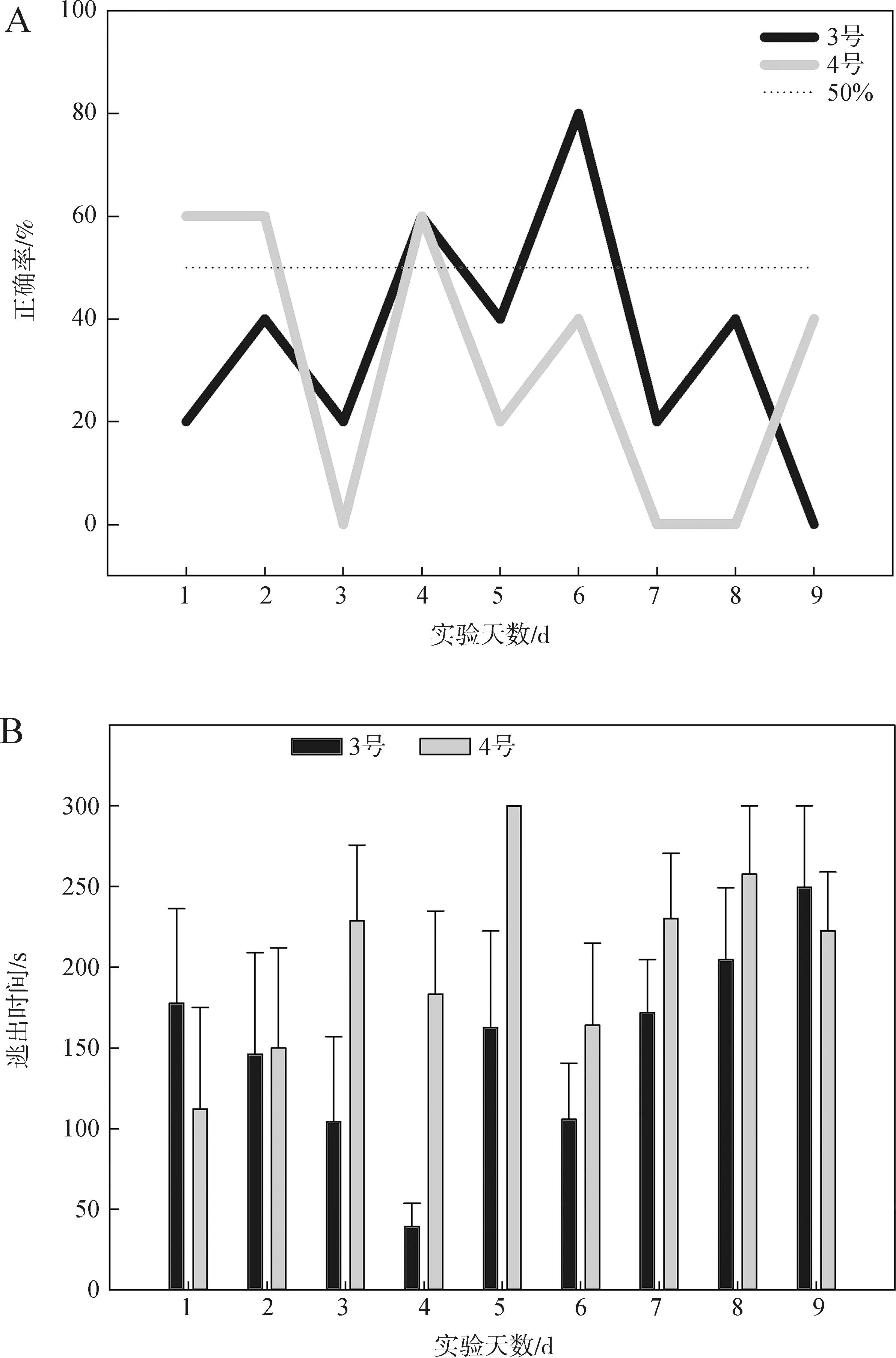
图6 第二组水刺激实验选择正确率(A)与逃出时间(B)Fig.6 Choice accuracy (A) and escape time (B) in the water stimulation experiment 2
3号个体选择正确率在前6 d实验中逐步提高,随后降低至低于随机水平,其正确率与随机选择差异无统计学意义(P>0.05);该个体逃出时间在前4 d实验中逐渐缩短,但随后恢复至较长时间,在8 d实验内未见明显变化(P>0.05)。4号个体选择正确率总体仍低于50%,与随机水平间的差异无统计学意义(P>0.05),其逃出时间在 8 d 实验内未表现出显著变化(P>0.05)。
2.2 高温刺激
实验一共分为3期进行。第一期实验使用3号、4号个体,进行5 d共计34次有效实验,实验个体未表现出习得训练行为,其选择正确率与逃出时间见图7。
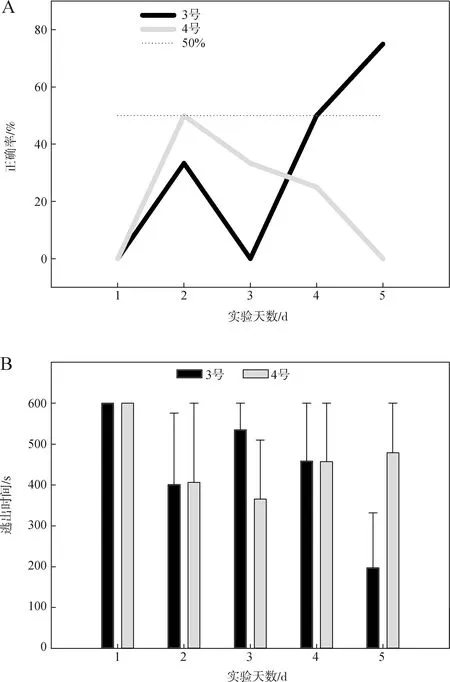
图7 高温刺激第一期实验选择正确率(A)与逃出时间(B)Fig.7 Choice accuracy (A) and escape time (B) in the high temperature experiment 1
3号个体选择正确率在前3 d实验中维持在较低水平,后稍有增长,但总体与随机水平(50%)之间的差异无统计学意义(P>0.05),其逃出时间在5 d实验内的差异无统计学意义(P>0.05)。4号个体选择正确率始终低于50%,其逃出时间在实验过程中未表现出显著差异(P>0.05)。
第二期实验使用1号、5号个体,进行9 d共计75次有效实验,实验个体未表现出习得训练行为,其选择正确率与逃出时间见图8。
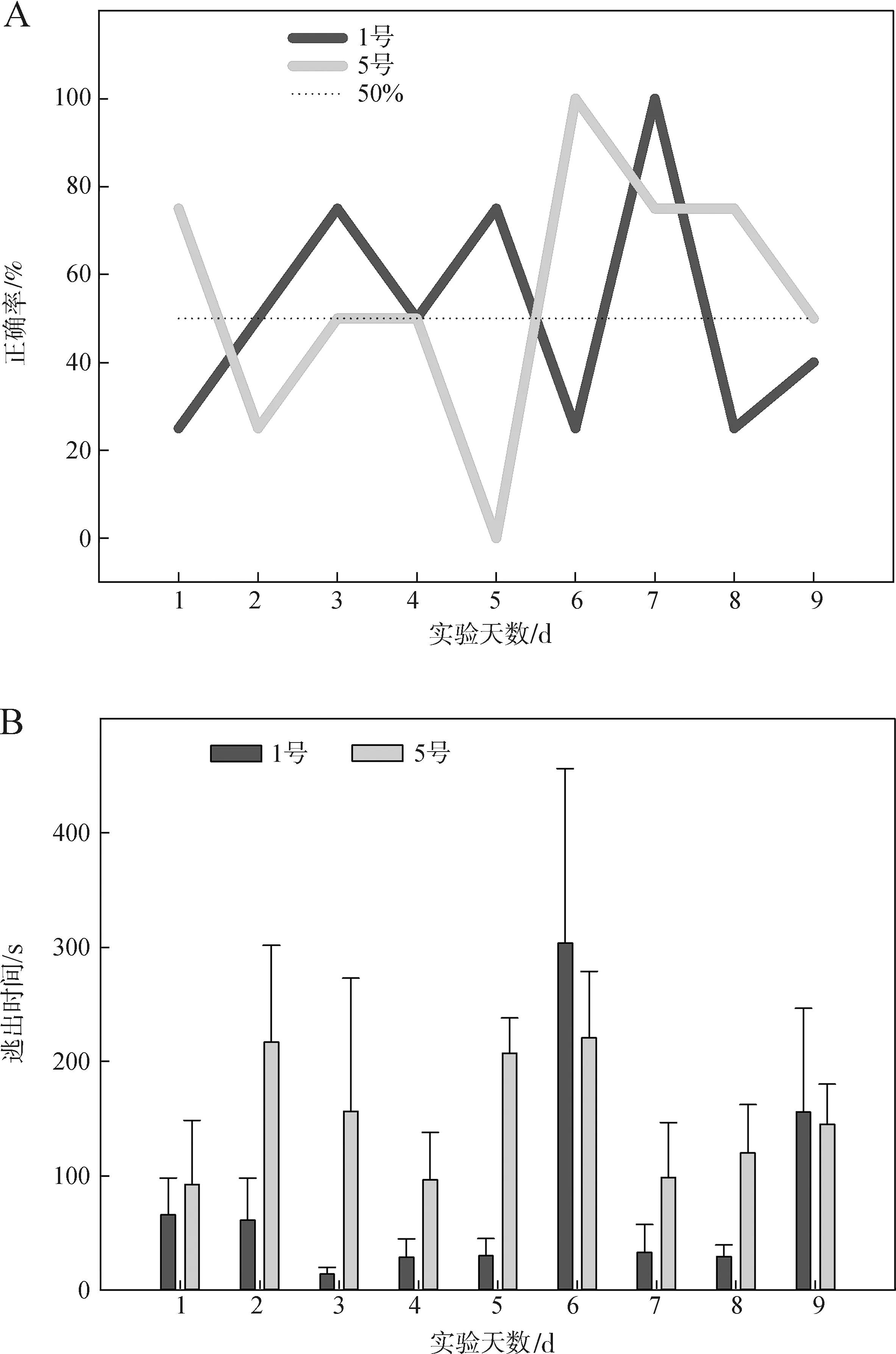
图8 高温刺激第二期实验选择正确率(A)与逃出时间(B)Fig.8 Choice accuracy (A) and escape time (B) in the high temperature experiment 2
1号与5号个体的选择正确率于随机水平(50%)上持续波动,差异无统计学意义(P>0.05)。3号与5号个体的逃出时间在9 d的实验过程中未表现出显著差异(P>0.05)。
综合前2期实验的结果,第三期实验每次实验时间延长至20 min。使用6号、7号与10号个体,进行5 d共计56次有效训练,除6号个体外,其他实验个体未达到学习标准,其选择正确率与逃出时间见图9。
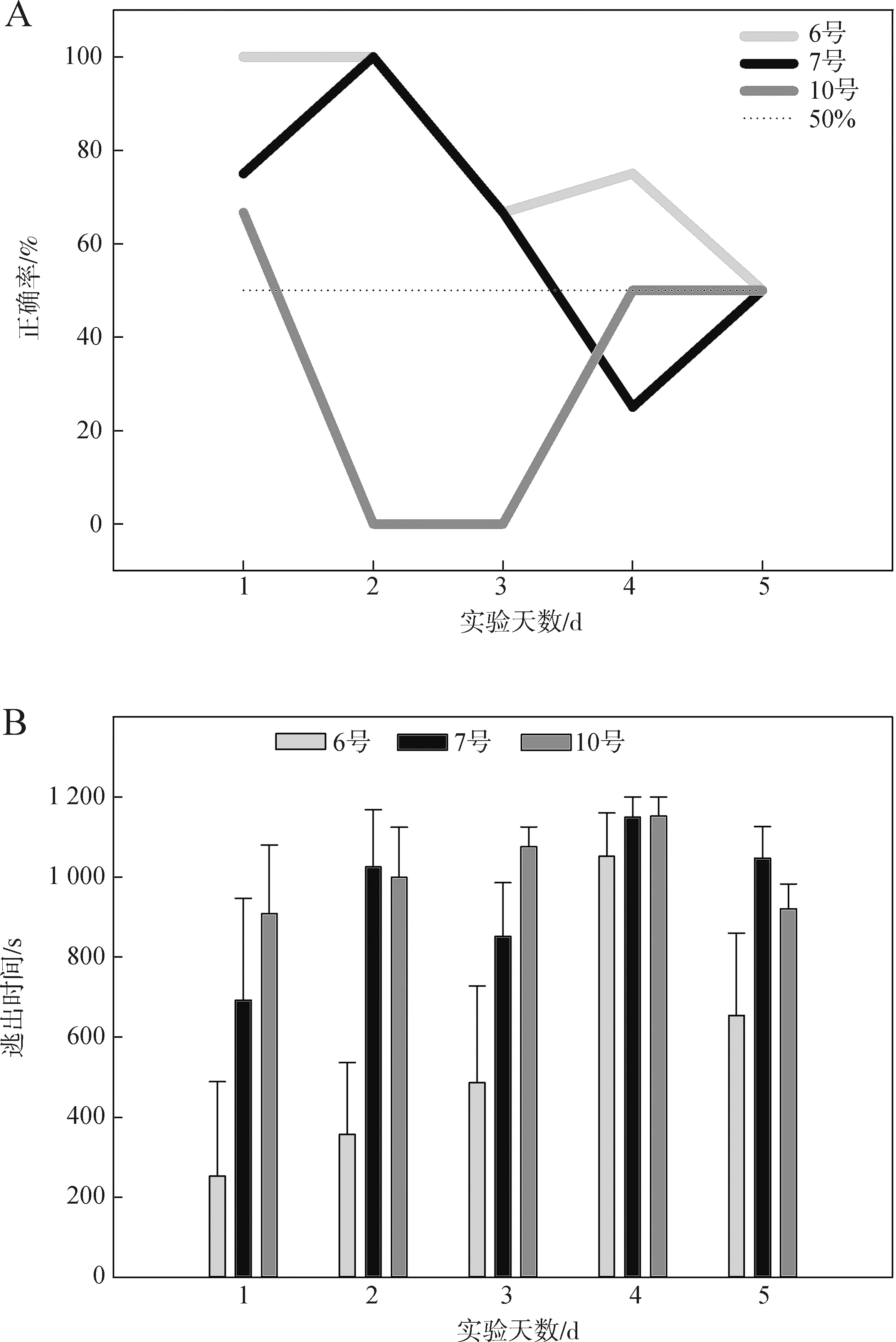
图9 高温刺激第三期实验选择正确率(A)与逃出时间(B)Fig.9 Choice accuracy (A) and escape time (B) in the high temperature experiment 3
6号个体选择正确图形的次数显著高于随机水平(50%)(P=0.012),但成功率随实验天数增加呈下降趋势,其逃出时间总体呈增加趋势,但随实验天数增加未表现出显著差异(P>0.05)。7号个体的选择成功率略呈下降趋势,与随机水平之间的差异无统计学意义(P>0.05),逃出时间在5 d实验内变化差异无统计学意义(P>0.05)。10号个体选择正确率与随机水平之间的差异无统计学意义(P>0.05),逃出时间在实验过程中无显著变化(P>0.05)。
2.3 轻敲刺激
无墙迷宫实验使用6~10号共5条个体,进行5 d共计91次有效实验,逃出时间见图10。
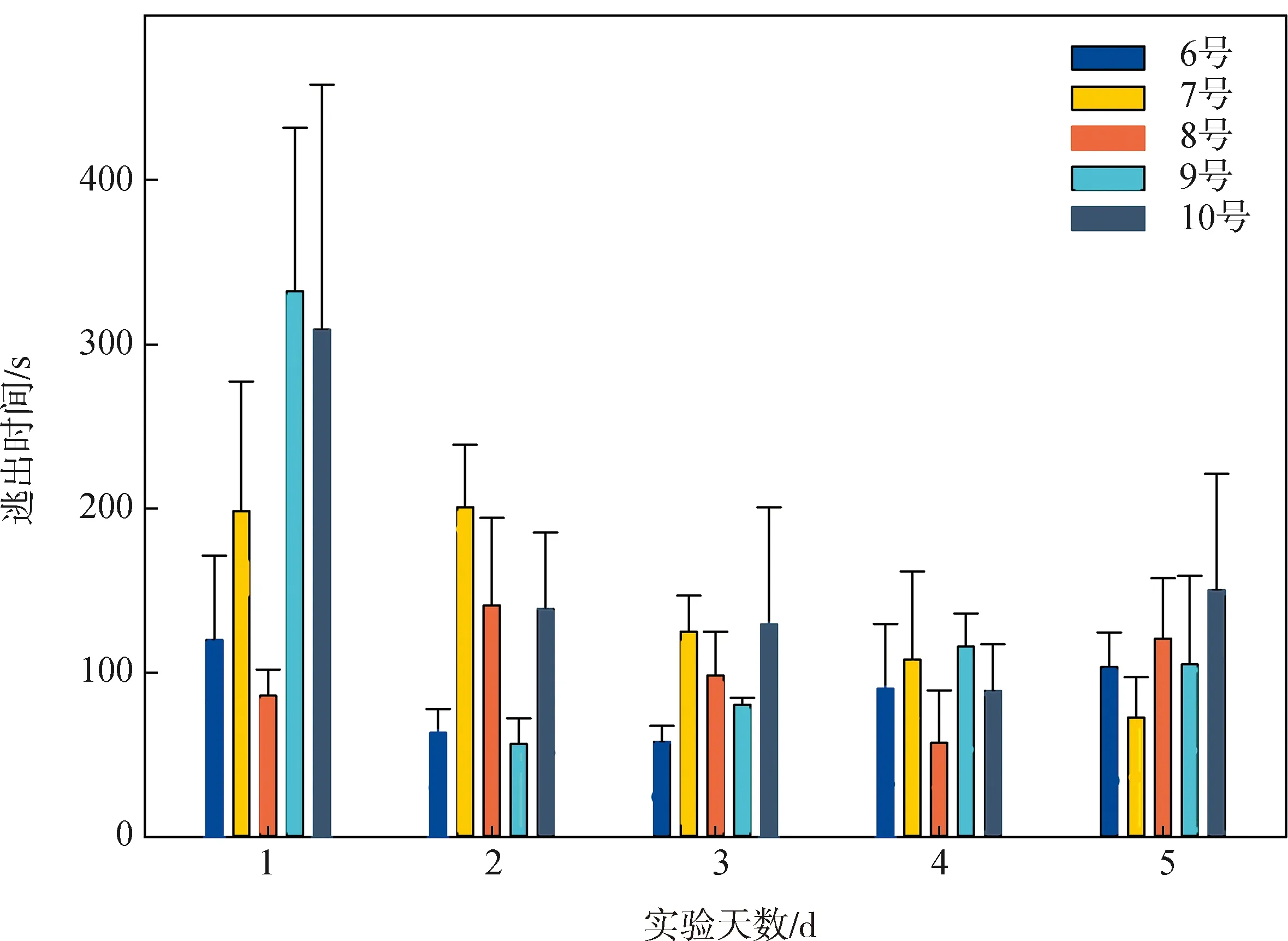
图10 无墙迷宫实验逃出时间Fig.10 Escape time in the wall-less Y-maze experiments
在5 d实验中,全部实验个体均未达到学习标准;多数个体在每天4次实验中正确、错误选择各2次,即为随机选择(50%)。在连续实验中,尽管9号个体逃出时间明显缩短,7号、10号个体的逃出时间也表现出下降趋势,但全部个体逃出时间在5 d实验内均未表现出显著差异(P>0.05)。
3 讨论
本实验使用水、高温和轻敲作为惩罚刺激,驱使短尾蝮离开适应区、进入迷宫安全区一侧逃离惩罚刺激,以此训练短尾蝮学习识别安全区信号的行为。3个不同刺激的实验中,尽管部分个体在短期内曾表现出选择正确率上升和逃出时间缩短的现象,但统计学上未表现出习得训练行为。
3.1 水刺激
习惯化是动物在重复刺激下对该刺激行为反应降低的现象,是一种基础的学习形式(Harris,1943),在动物界极为常见,蛇类的行为学研究中亦有发现(Glaudas,2004;Place,2005;Place &Abramson,2008)。水刺激惩罚实验说明,这一强度较低的刺激难以长时间维持,实验动物很快表现出对水环境的适应性,不再积极逃离,无法持续进行条件反射训练。水刺激惩罚实验中,可见短尾蝮停留、浸泡于水体中的现象。在第一组115次实验中,共计32次观察到该现象,其中24次存在于2号个体,该个体明显较适应水环境。3号和5号个体的不逃离行为共发生8次,其中3次均发生于第2天实验,另外5次集中于最后3 d实验。这种行为导致3号和5号个体的选择正确率下降与逃出时间延长,表明蛇已逐步适应水刺激,不再倾向于移动、躲避。第二组实验显著增加了注水量,主要参考平原束带蛇的实验设计与步骤,但仍未能观察到短尾蝮的学习迹象。第二组共计88次实验,33次蛇停留于水体中的时间超过实验时间,其中19次发生于最后4 d实验中。这一现象同第一组实验一致,表示蛇对水刺激已逐渐适应,不再积极逃离水体。
3.2 高温刺激
实验过程中部分个体存在明显的趋触性行为,即蛇的身体紧靠垂直面、沿迷宫壁爬行;这一行为在多数蛇类中均可观察到(Ford &O’Bleness,1986;Costanzo,1989;Greeneetal.,2001)。趋触性行为显著混淆了实验结果:短尾蝮并非主动选择进入某一侧,其选择取决于首先接触的迷宫壁,随后则沿壁爬行被动进入对应一侧迷宫臂。因此,蛇类Y迷宫实验中应注意考虑消除趋触性的影响。高温刺激惩罚实验中,第三期实验中的6号个体选择正确的次数显著高于随机水平,然而该个体并非学会了持续进入安全区。在未经训练的第1天实验中,6号个体表现出100%的正确率,并在第2天实验中维持了这一正确率。尽管图形位置为随机决定,实际在该个体前2 d(共8次)的实验中,7次实验内安全区位于右侧。在预实验的12次空白实验中,6号个体并未表现出显著的方向偏好性。随着实验进行,该个体的正确率逐步下降;同时,其逃出时间反而逐步上升。以上数据说明,6号个体的较高正确率可能仅为小概率事件。
实验过程中,高温刺激对短尾蝮的驱避作用并不明显。第一期和第二期实验共109次实验中,存在23次蛇停留不动、直至实验结束的情况。在第三期实验中,实验时间延长至20 min;但在共计56次实验中,仍存在15次相同的情况。部分个体明显更耐热,3号、4号与7号个体可忍受加热时间达25 min而无明显行动。这一现象在以往研究中亦存在(Krochmal &Bakken,2003;Krochmaletal.,2004);为解决这一问题,Krochmal等(2004)将迷宫温度进一步升高至45 ℃甚至50 ℃以迫使实验动物逃离迷宫。本实验也尝试了这种方法。当短尾蝮置于迷宫中体温升高至较高温度时,可观察到其呼吸频率显著增高(Seebacher &Franklin,2005)。随后蛇产生“惊恐式”窜逃行为,表现为身体颤动、急速而无序地爬行。此时,若蛇进入惩罚区域,即使给予其高频率(1 s/次)轻敲头部的惩罚亦不能迫使其返回温度较高的迷宫主体部分;该状态下蛇对致死温度的躲避行为已不具备条件反射训练的可行性。
3.3 轻敲刺激
相较于水刺激与高温刺激实验,轻敲刺激实验有效避免了趋触性。逃出时间显著缩短,绝大多数实验在5 min内完成;表明轻敲刺激强度较高,短尾蝮对该刺激存在明显逃避行为。在实验过程中可观察到蛇面对木棒敲击时表现出较激烈的反应,其典型行为为蛇的头部朝向木棒尝试攻击,同时身体蜷缩、倒退进入某一侧迷宫臂。91次实验中,记录到6次因试图躲避刺激时窜逃而摔下迷宫的现象,同时存在约10次扑咬木棒的行为。
3.4 惩罚刺激缺陷
综合全部实验,实验现象体现出单一惩罚刺激的缺陷。在Y迷宫实验中,需要有效刺激驱使蛇离开起始区域,并作为惩罚区域对应的惩罚。在一系列先前研究曾使用或本研究尝试使用的惩罚刺激中,构成了由弱到强渐变的多种刺激。然而,对于强度较低的弱刺激,如强光(预实验中尝试使用)、水(水刺激惩罚实验)、振动(Crawford &Holmes,1966)等,蛇表现出明显的适应性(即习惯化),实验进行数天后蛇对于该刺激的行为反应弱化甚至不再逃避。对于强度较高的刺激,如电击(Wolfle &Brown,1940;Mills,1970)、轻敲刺激等,蛇行为反应过激,突然窜逃、挣扎、扑咬的现象持续存在,不利于条件反射的训练。加热刺激虽是较为合适的中强度刺激,但具有时间累计效应的特点,实验中短尾蝮表现出了较强的耐热能力,延长了加热时间,失去了训练的可行性。
短尾蝮在相似的实验设计下的学习表现弱于先前研究中的平原束带蛇、西部菱背响尾蛇Crotalusatrox、彩虹石龙子Lampropholisdelicata等动物(Fuenzalida &Ulrich,1975;Krochmal &Bakken,2003;Chungetal.,2017)。爬行动物类群中,蛇类与蜥蜴具有一定的学习能力,但认知能力弱于龟鳖类与鳄类(Northcutt,2013)。因此,可能原因之一为安全区位置的随机变换增加了学习难度。实验中刺激亦可能影响了动物的本能反应。短尾蝮虽是地栖型蝮蛇,但自然环境中存在降雨等水刺激,因此持续使用水刺激时,蛇可逐步对刺激形成习惯化。蝮蛇为变温动物,体温随环境温度波动。使用加热刺激时,刺激初期蛇体温较低,其本能行为可能更倾向于停留原处、升高体温,由此延长了加热时间,直至体温过热引起窜逃行为。在无墙迷宫实验中,因刺激强度较高,持续刺激时蛇的本能可能由逃离转为防御性攻击行为。在攻击行为中,短尾蝮头部朝向木棒,已无法观察身后迷宫臂是否为安全区域,不利于学习训练。
综上,由实验现象可见短尾蝮表现出“低敏感度、高耐受性”的特点。惩罚实验效果提示,在动物条件反射训练中,实验设计应符合动物在实验即时环境中的自然行为(即时本能)(Place,2005)。建议在未来的行为学实验中,多使用其自然行为作为动物认知能力的判断依据。
致谢:感谢日本京都大学福田将矢博士与中国科学院成都生物研究所孙晓倩同学在实验操作过程中给予的帮助,感谢中国科学院成都生物研究所两栖爬行动物研究室各位老师在实验设计、修改过程中提供的指导。
猜你喜欢
科普童话·百科探秘(2022年2期)2022-05-24
科普童话·百科探秘(2022年1期)2022-05-24
医学食疗与健康(2022年3期)2022-04-23
作文评点报·作文素材初中版(2020年15期)2020-04-24
故事家·花开不败(2019年2期)2019-09-10
生物学教学(2018年3期)2018-11-29
数学小灵通·3-4年级(2018年7期)2018-07-12
农村百事通(2018年6期)2018-03-29
第二课堂(小学版)(2018年2期)2018-03-26
软件(2018年1期)2018-02-05
