
乳源外泌体的组成及其生物功能研究进展
2020-07-23 04:23仝令君郝海宁张兰威易华西
食品科学 2020年13期
仝令君,郝海宁,张兰威,易华西*
(中国海洋大学食品科学与工程学院,山东 青岛 266003)
哺乳动物母乳作为幼体的主要营养来源,含有蛋白质、乳糖、脂肪、维生素、矿物质等多种营养素以及神经元生长因子、生长调节因子、免疫相关因子等重要生理活性物质[1-2],对新生儿的生长发育和免疫系统的形成具有重要作用[3-5]。外泌体作为母乳的成分之一,能够携带和传递重要的信号分子,其信号分子的特性与宿主细胞的生理状态密切相关,目前已经成为国内外的研究热点。外泌体是一种能被大多数细胞分泌的微小膜泡,具有脂质双层膜结构,直径大约30~200 nm,存在于绝大多数真核生物的体液中,包括血液、尿液、唾液、脑脊液以及乳液中[6-7]。外泌体的概念最早在1983年被提出,但一直被认为只是细胞的废弃物,近几年研究发现在外泌体中含有细胞特异的蛋白质、脂质和核酸,能作为信号分子传递给其他细胞从而改变其他细胞的功能[8-9]。母乳是一种丰富的外泌体来源,初乳中外泌体浓度远高于成熟乳[10]。乳源外泌体无论在疾病诊断方面,还是在婴幼儿生长发育方面,都具有重要的研究价值。目前国外关于牛乳外泌体、人乳外泌体等乳源外泌体的功能研究报道较多[9,11]。研究表明,乳源外泌体不仅具有作为药物运输载体、疾病筛查标志物的潜力,还有望被研制开发为一种新型的益生元[12-13],但我国有关外泌体的研究还处于起步阶段。据此,本文综述了乳源外泌体的组成、功能和应用前景,旨在为我国乳源外泌体的研究与开发提供思路。
1 外泌体的形成途径
外泌体是细胞内多泡体与细胞膜融合后释放到胞外基质中的一种膜性囊泡。目前认为外泌体的形成途径有两种(图1)。第一种为外泌体形成的经典途径(图1a)。首先,细胞内形成包含有蛋白质和RNA等胞内生物分子的神经酰胺依赖性晚期囊泡,在神经酰胺的作用下,囊泡中的内容物避开溶酶体的消化[14];然后这些囊泡融合到胞内形成的多胞体(multivesicular body,MVBS)中,接着MVBS被整合到质膜上,并将外泌体(囊泡)释放到细胞外基质中[15]。蛋白质和RNA等内容物在外泌体形成过程中被分选并载入外泌体,整个过程中涉及了20多种囊泡分选蛋白,其中最重要的是内体运输分拣复合物(endosomal sorting complex required for transport,ESCRT)和液泡蛋白分选因子(vacuolar protein soaing 4,Vps4)。ESCRT为细胞膜塑形蛋白,可以介导内吞囊泡出芽,加速腔内囊泡的形成;Vps4可以重新塑造ESCRT再生,便于其循环再利用。ESCRT和Vps4在外泌体形成、载体蛋白分拣过程中起着不可或缺的作用[16-18]。随着研究的深入,发现外泌体形成除“经典途径”外,还存在第二种更为直接的形成途径(图1b)。T细胞和红白血病细胞系直接从其质膜释放外泌体。人类免疫缺陷病毒的Gag或Nef蛋白在T细胞、红细胞和白细胞表达后,这3 种细胞的细胞膜发生内陷,形成并释放外泌体[19-21]。这些小泡与经典途径形成的外泌体难以区分,也富含CD63和CD81等经典外泌体标记物,其直径和密度与外泌体也极为相似,目前这种形成途径的机制尚不清楚。有研究报道指出,乳源外泌体主要来自乳腺上皮细胞,其生物发生是通过细胞质膜内囊泡的向内出芽(内吞)来启动的(外泌体形成的经典途径),随后生物发生继续随着晚期内吞体限制膜的内陷形成多泡体并将“货物”分选到这些多泡体内,接着形成腔内囊泡并被整合到质膜上,最后以外泌体(囊泡)的形式出芽[13,22]。
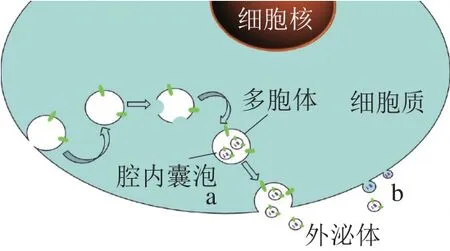
图1 外泌体形成的两种途径Fig. 1 Two pathways of exosome formation
2 乳源外泌体的结构与组成
外泌体是具有脂质双层膜结构的膜外小泡,其具有与细胞相同的拓扑结构,直径约为30~200 nm。不同方法测得的外泌体大小存在一定差异,即使是单个细胞分泌的外泌体也是如此。每种外泌体测定技术都会引入相应的误差范围进行校正。通过透射电子显微镜、原子力显微镜或者冷冻电子显微镜对外泌体的结构进行表征,几乎所有表征都显示出简单的球状形态。大约一半数量的外泌体直径都在预期的30~200 nm直径范围内,其余显示为直径较大的微泡。此外,极少数外泌体呈现不寻常的膜状结构,甚至具有其他不规则的形态[23]。这些不规则的结构可能与外泌体形成过程中某个定向生物过程有关,但是外泌体提取过程中的物理机械压力也可能是导致这些结构形变的主要原因[24]。
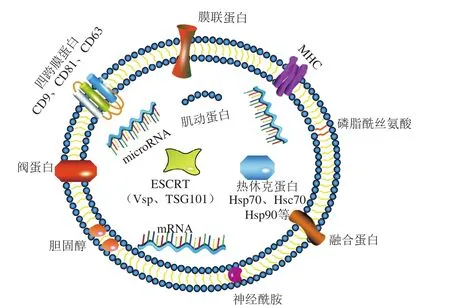
图2 外泌体结构Fig. 2 Structure of exosome
外泌体主要的组成部分是蛋白质、核酸、脂质。如图2所示,外泌体由双层磷脂膜包裹,含有较高水平的胆固醇、鞘磷脂、神经酰胺以及短/长链饱和脂肪酸[25-26]。不同细胞来源的外泌体,虽然组成存在不同程度的差异,但其表面都具有相似的保守蛋白,如CD63、CD81、CD9等。这些标志蛋白在外泌体分离鉴定过程中具有重要靶标作用。此外,不同来源的外泌体蛋白组成存在一定差异,肠上皮细胞来源的外泌体含有特异性A33抗原,而免疫B细胞外泌体中富集有CD86和组织相容性复合体(major histocompatibility complex,MHC)。不同动物乳外泌体中含有多种可调节细胞免疫功能的蛋白质,如载脂蛋白、白细胞介素(interleukin,IL)-19等[27]。Théry等[28]从树突状细胞来源的外泌体中共检测到21 种蛋白质;Merchant等[29]从人体尿液外泌体中检测到43 种蛋白质;Reinhardt等[30]从牛乳外泌体中共检测到2 107 种蛋白质;Yang Mei等[27]从母乳和牛乳不同泌乳期样品中一共鉴定出920 种外泌体蛋白。上述研究表明,相较于其他体液,牛乳和母乳中外泌体蛋白的含量更为丰富。母乳外泌体中含有丰富的免疫相关蛋白、转运相关蛋白、参与核糖体途径的蛋白以及肌动蛋白细胞骨架调节蛋白。这些蛋白在婴幼儿免疫调节[31]、细胞间传导[32]、蛋白质合成、mRNA翻译[33]以及肌动蛋白相关细胞调节[34]中起着至关重要的作用。
核酸是外泌体中的另外一种主要组分,对外泌体发挥生理功能具有重要意义。乳源外泌体中核酸主要包括非编码单链RNA 分子(microRNA)、长链非编码RNA(long non-coding RNA,lncRNA)、环状RNA(circular RNA,circRNA)、mRNA、tRNA等[35-36]。序列同源性分析表明,microRNA约占乳源外泌体平均RNA含量的13%[37]。microRNA是由约70 nt的长茎环前体衍生而来的约22 个核苷酸非编码小RNA,已成为基因表达中关键的转录后调控因子,在宿主生理和病理过程中发挥重要作用[38]。乳源外泌体microRNA也可以进入人体循环并调节外周血细胞中的基因表达[39]。与成熟乳相比,初乳的外泌体中含有丰度更高的免疫相关microRNA[40]。lncRNA是一类转录本长度超过200 nt、非编码蛋白的RNA。lncRNA能在表观遗传、转录及转录后水平上调控基因表达。近期研究发现lncRNA存在于生物体液中,一部分lncRNA游离于体液中,一部分被包裹在外泌体中进行远距离的信息传递[41]。CircRNA是一类特殊的非编码RNA分子,也是RNA领域最新的研究热点。与传统的线性RNA(含5’和3’末端)不同,CircRNA分子呈封闭环状结构,不受RNA外切酶影响,表达更稳定,不易降解。研究发现外泌体CircRNA能够通过远距离的信号传递来调节受体细胞内的生物功能[42]。核小RNA(small nuclear RNA,snRNA)在前体RNA的剪接中发挥作用,而核仁小RNA(small nucleolar RNA,snoRNA)主要指导rRNA、tRNA和snRNA的修饰[13]。由于脂质双层膜的保护作用,乳源外泌体中核酸在低pH值、高温和酶处理等条件下非常稳定[43],为其进入人体循环并发挥功能提供了基础。
3 乳源外泌体的生物学功能
3.1 乳源外泌体与肠道发育
肠道不仅是营养物质消化吸收的主要部位,也是重要的免疫器官和内分泌器官[44]。在新生儿肠道中,营养运输沿着整个隐窝-绒毛轴线发生[45]。胃肠道作为负责将营养物质运送到身体细胞的第一个生理器官,在调节婴幼儿发育和健康方面发挥着重要作用[46]。乳源外泌体在婴幼儿肠道发育方面具有重要作用[47-49]。乳源外泌体耐受核糖核酸酶消化,并不受冻融循环和酸度的影响,允许外泌体内容物完整地到达新生儿的肠腔并被吸收[48]。Wolf等[50]研究表明,肠道对牛乳外泌体的摄取是通过内吞作用及膜融合作用来介导的,并依赖于肠道中的细胞和外泌体表面糖蛋白,而肠道对牛乳外泌体中microRNA的吸收途径主要是在顶端到基底外侧方向的单向过程。Chen Ting等[49]研究表明,猪乳来源的外泌体可以促进猪小肠细胞增殖和肠道发育,并提高猪小肠细胞中尾侧型同源盒基因-2(caudal-related homeobox gene 2,CDX2)、增殖细胞核抗原(proliferating cell nuclear antigen,PCNA)、胰岛素样生长因子1受体(insulin-like growth factor 1 receptor,IGF-1R)基因的表达以及抑制p53抑癌基因(tumor suppressor gene p53,p53)的表达。CDX2是肠道特异性转录因子,在整个肠道上皮细胞核中表达,直接参与肠发育、肠表型的保持和维持[51]。PCNA与细胞周期相关,并在DNA复制和修复中起作用[52]。已有报道PCNA免疫定位可用作正常组织和淋巴样肿瘤中细胞增殖的指标[53]。IGF-1R下游Akt/Wnt信号通路在细胞增殖中起关键作用。此外,猪乳来源的外泌体可以显著增加小鼠十二指肠和空肠的绒毛高度和隐窝深度,从而提高绒毛高度(villus height,V)与隐窝深度(crypt depth,C)的比值。目前认为V/C越高,表明肠道发育以及免疫功能越好,营养物质更易被吸收[54]。Hock等[55]指出,小鼠(母)乳来源的外泌体可以促使PCNA基因相对表达显著增加,从而提高小鼠肠上皮细胞活力,促进细胞增殖,刺激肠干细胞活性。乳源外泌体可以在肠系统内“行进”并被Lgr5+干细胞摄取,上调Lgr5基因表达,提高肠道上皮细胞的活力[55],并通过上调肠道CDX2、PCNA、IGF-1R基因的表达,促进肠道细胞增殖,从而促进婴幼儿肠道发育[49]。乳源性外泌体还参与母乳介导的抑制Toll样受体(Toll-like receptor,TLR)4表达,从而保护肠上皮免受细胞凋亡和增强肠上皮细胞增殖,并在肠杯状细胞分化中起作用,防止过度炎症,并维持肠上皮完整[48]。此外,饮用牛奶后,会在人体血浆和肝脏中检测出牛乳外泌体中特异性microRNA,表明来自动物源食物中的microRNA可以通过肠道上皮组织进入人体循环,对于人类的营养和健康具有非常重要的意义[56]。Wang Yanhong等[57]研究发现,由于CircRNA比线性RNA更稳定,因此在膳食乳外泌体中携带的circRNA有更多机会进入体内组织并发挥特定的生物学功能。例如,衍生自染色质重塑基因ARID1A反义RNA的CircRNA59(在乳源外泌体中含量高)也在小鼠和人组织中表达。ARID1A促进心脏祖细胞分化,抑制子宫上皮细胞增殖[58]。ARID1A损伤可重塑染色质,并在多种肝损伤后改善再生[59]。
外泌体如何介导新生儿肠道发育是目前的研究热点,也是研究难点。母乳外泌体可以将蛋白质、核酸等转运到新生儿的肠道系统,保护其免受酸度降解和消化,并使其保持结构完整及被吸收[48],这是乳源外泌体可以刺激肠道发育最为关键的一步。Good等[60]研究报道,TLR4的激活参与坏死性小肠结肠炎(necrotizing enterocolitis,NEC)的发生与进程,但母乳中的表皮生长因子可以抑制TLR4信号的传导,抑制NEC的发生,从而保护肠上皮细胞免于凋亡并促进肠细胞增殖。母乳来源的外泌体是否通过运输表皮生长因子到达肠道,参与抑制TLR4信号的传导,促进肠细胞增殖,这有待进一步深入研究。Pieters等[61]研究表明,乳源外泌体中富含转化生长因子β(transforming growth factor beta,TGF-β),其在婴儿时期对肠屏障功能的发育、免疫球蛋白(immunoglobulin,Ig)A的产生以及黏膜免疫起着重要的作用。此外,几种母乳外泌体microRNA已被证明在肠道杯状细胞分化中具有预防过度炎症、保持肠上皮完整的功能[48,62-64]。Ma Jideng等[65]指出大熊猫初乳和成熟乳外泌体中富含与免疫和肠道发育相关的内源性microRNA,如let-7家族、ame-miR-148a-3p、ame-miR-30a-5p等。乳源外泌体中miR-135b-5p和miR-664b-3p也可以分别通过靶向肌肉萎缩相关蛋白MEF2C和PLP2促进肠道细胞增殖[66-67]。Kosaka等[62]发现乳源外泌体中的miR-223可以作为肠道嗜中性粒细胞增殖和活化的调节剂,而let-7i作为TLR4的调节剂,在NEC中可以抑制TLR4信号的传导,保护肠上皮细胞免于凋亡并促进肠细胞增殖。近年来,肠道微生物已经被证明对宿主具有重要生理调节功能,关于母乳外泌体对肠道微生物组成是否具有调控作用、是否可以通过调节肠道菌群介导新生儿肠道发育的研究报道较少,是值得深入研究的方向之一。
3.2 乳源外泌体的免疫功能
母乳是一种具有免疫细胞的复杂液体,可为婴幼儿提供免疫因子,影响婴幼儿免疫系统的发育与成熟。母乳外泌体携带抗原结构,表明其能够参与指导免疫反应。研究发现,母乳外泌体可以抑制抗分化簇3诱导的来自异源和同源外周血单个核细胞(peripheral blood mononuclear cell,PBMC)中的IL-2和干扰素-γ的产生。另外,在乳源外泌体培养的PBMC中,外泌体中MHC II与CD86的表达对CD4+T细胞具有刺激作用,从而促使调节性T细胞(regulatory cell,Treg)Foxp3+、CD4+、CD25+数量增加,表明母乳外泌体具有免疫调节能力[32]。母乳是婴幼儿重要的免疫支持系统,乳源外泌体在婴儿免疫系统中具有双重功能,外泌体能够参与婴幼儿免疫刺激[68]和耐受[69],发挥类似益生元的功能,为研究与开发乳源外泌体这一新型益生元奠定了理论基础。
Zhou Qi等[8]对母乳外泌体中的microRNA组成和转录组学进行了研究,发现高特异性microRNA占所有microRNA数量的62.3%。通过Pathway Central分析,有4 种前体microRNA与免疫相关,分别为miR-148a-3p、miR-30b-5p、miR-182-5p、miR-200a-3p。miR-30b-5p促进细胞侵袭和免疫抑制[70],miR-182-5p由IL-2诱导,能促进T细胞介导的免疫应答[71],miR-200a-3p与霍奇金淋巴瘤和口腔癌症相关[72-73],miR-148a-3p调节诱导多种转运蛋白和药物代谢酶表达[74-75]。miR-148a-3p在牛奶中高度表达,被认为是可作为识别牛奶和其他乳的潜在生物标志物[76]。Ma Jideng等[65]对大熊猫不同哺乳期中母乳外泌体及外泌体中的microRNA进行了表达特征分析,发现大熊猫初乳和成熟母乳中富集有与免疫和发育相关的内源性microRNA。研究结果表明,大熊猫母乳中外泌体microRNA可能是新生大熊猫幼崽发育的关键母体“调节者”。Zeng Bin等[77]通过RNA测序对牛乳外泌体中的lncRNA进行了全面分析,研究表明,从牛乳外泌体中共鉴定出3 475 个新的lncRNA和6 个可注释的lncRNA。lncRNA表达量比mRNA更高。基因本体(gene ontology,GO)注释和京都基因与基因组百科全书(Kyoto encyclopedia of genes and genomes,KEGG)途径分析表明,lncRNA具有调节免疫功能,促进成骨细胞生成、神经发育、繁殖、细胞增殖和细胞间通讯等方面的功能。此外,Lönnerdal等[78]发现母乳外泌体中与免疫功能相关的高丰度microRNA为miR-146、miR-181、miR-21、miR-16和let-7,而Zhou Qi等[8]研究发现丰度前10的microRNA中,其中与免疫相关的4 个高丰度microRNA为miR-148a-3p、miR-30b-5p、miR-182-5p、miR-200a-3p。上述研究结果存在一定差异,表明乳源外泌体microRNA对免疫功能的影响还需进一步深入系统研究。
流行病学研究发现,婴儿出生的第一年进行牛乳饮食可以防止特应性疾病的发展,增加Treg的数量[79]。牛乳外泌体来源的microRNA可能通过促进T细胞下调IL-4/辅助性T细胞(IL-4/T helper 2 cell,IL-4/Th2),也可能通过诱导胸腺Treg谱系所需的关键免疫调节因子和表观遗传修饰因子来介导特应性致敏作用和免疫应答[40]。牛乳外泌体中miR-155与免疫系统的发育密切相关,控制参与调节FoxP3(Treg的标志性分子)表达、IL-4信号转导和免疫球蛋白类转换为IgE和IgE的高亲和力受体——关键靶基因FcεRI的表达[80]。这些研究都表明乳源外泌体在免疫调节中可能具有不可或缺的作用。然而,加热及均质处理对牛乳外泌体microRNA介导的生物活性具有负面影响[46]。加热及均质处理是乳制品加工中不可缺少的操作单元,婴幼儿配方奶粉由于缺失了具有生物活性的外泌体microRNA,从而影响FoxP3表达[76]。在奶粉喷雾干燥后期通过干混加入牛乳外泌体能够避免上述影响,最大程度地保护牛乳外泌体microRNA的生理活性。
3.3 乳源外泌体在炎症预防方面的作用
炎症反应是临床常见的病理反应,可以在机体各部分的组织和器官中发生。发掘新型对机体炎症适度调节的天然功能成分对预防及治疗由宿主异常炎症反应造成的健康隐患具有重要意义。研究表明,免疫细胞可能通过分泌外泌体介导炎症[81-82]。来源于红细胞、间充质干细胞和支气管肺泡灌洗液的外泌体已被证明具有促进促炎细胞因子产生的作用[83-85]。体液来源的外泌体可以通过TLR-2和TLR-4激活核因子κB信号通路,导致促炎细胞因子mRNA和蛋白水平升高[86]。然而,这些来源的外泌体由于获取比较困难,其作为调节机体炎症的天然功能成分具有一定局限性。牛乳是大规模生产外泌体的可靠来源[87]。最近研究表明,乳源外泌体在改善关节炎方面具有一定作用[9]。类风湿关节炎是炎症反应的一种,其发展与遗传和环境因素有关,一些研究表明与生活方式和饮食因素(主要是牛乳的摄入量)有关,其发病机制可能是这些因素会通过影响调节T细胞应答来促使类风湿关节炎的发展[88]。Arntz等[9]研究了牛乳外泌体对小鼠关节炎的影响,对两种T细胞依赖的慢性和破坏性关节炎模型小鼠灌胃乳源外泌体,结果发现两种老鼠的关节炎明显减轻,同时由脾细胞产生的单核细胞趋化蛋白-1(monocyte chemotactic protein 1,MCP-1)和IL-6的血清水平降低,表明口服外泌体可改善实验性关节炎。牛乳外泌体中还包含许多具有免疫调节作用的microRNA和lncRNA[40,76-77],例如直接靶向MCP-1 mRNA的miR-124a[89-90]、靶向TLR/IL-1炎症途径的miR-21、miR-let7、miR-146a、miR-146b、miR-126、miR-155、miR-199a[91-92],以及靶向miR-146a-5p的lncRNA核内小RNA宿主基因16(lncRNA SNHG16)[93],这些microRNA和lncRNA都可能在炎症预防方面发挥重要作用。除了核酸之外,牛乳囊泡中存在的一些蛋白质也发挥抗炎作用,如乳铁蛋白、载脂蛋白E和TGF-β[93-96]。上述研究为开发乳源外泌体作为口服益生元提供了理论依据。
3.4 乳源外泌体的其他生物学功能
Oliveira等[97]指出向小鼠口服递送乳源外泌体能促进其骨细胞数量以及编织骨形成,从而促进成骨细胞分化和骨基质形成。同时研究发现口服递送外泌体后,能够促进成骨细胞分化和小破骨细胞的形成,但会减少骨吸收和骨连接蛋白的表达,并抑制结节的形成,导致骨基质质量受到损害[98]。因此,乳源外泌体可能作为调节成骨细胞和破骨细胞分化的骨调节剂,在婴幼儿骨发育方面发挥一定的作用。此外,有研究发现乳源外泌体对小鼠骨骼肌基因表达、生殖能力以及空间学习与记忆能力都有一定的影响[99-101]。Leiferman等[99]指出,乳源外泌体可以显著提高骨骼肌发育相关基因Rhobtb1和Socs2的表达,对年轻成年小鼠骨骼肌中的基因表达和代谢略有影响。Mutai等[100]研究发现,与饲喂外泌体组相比,未饲喂外泌体的小鼠在Barnes迷宫中定位逃生洞所需的时间延长了130%,定位和到达Morris水迷宫中的水下逃生平台所需的时间明显延长。Sadri等[101]研究表明,雄/雌两者均饲喂乳源外泌体的小鼠平均产仔数是均不饲喂外泌体组的两倍。如果至少有一只小鼠不饲喂外泌体,62%~75%的交配不会导致怀孕。上述研究成果为乳源外泌体的生物学功能及其作为新型益生元的研究与开发提供了新思路。
4 结 语
外泌体是在细胞间和细胞内通讯过程中具有多种功能的纳米级囊泡,携带有蛋白质、核酸等含有丰富生物学信息的生物分子。目前对外泌体的研究主要集中在两个方面:一方面是以外泌体作为癌症等疾病筛查或诊断的标识物,期望开发出基于外泌体的疾病快速活检技术;另外一方面是利用外泌体的稳定性和传输性,以牛乳外泌体作为药物载体,携带药物用于疾病的靶向治疗。但是对人体母乳、牛乳等乳源外泌体生理功能方面的深入研究较少。已有的基础研究表明,乳源外泌体具有调节免疫、预防炎症、促进生长发育等多方面的生理功能,具有益生元的开发潜能。随着对乳源外泌体生理功能及其机制的深入研究,乳源外泌体有望作为一种新型的益生元,在婴幼儿配方奶粉、功能型饮料等功能食品的研究与开发中发挥重要作用。
猜你喜欢
安徽医科大学学报(2022年8期)2022-09-21
色谱(2021年9期)2021-09-06
食品安全导刊(2021年21期)2021-08-30
宝藏(2020年2期)2020-10-15
临床检验杂志(电子版)(2020年3期)2020-04-11
自我保健(2020年8期)2020-01-01
科学导报(2019年40期)2019-09-03
宝藏(2019年5期)2019-06-21
宝藏(2019年2期)2019-03-20
中国乳品工业(2018年11期)2018-12-29
