
2018-2019年舟山近海浮游动物群落结构春季年际变化及其与水团的关系
2020-07-21 05:34王慧君许永久俞存根章寒张洪亮蒋日进梁君郭爱朱文斌徐开达刘连为王皓学
海洋学报 2020年6期
王慧君,许永久*,俞存根,章寒,张洪亮,蒋日进,梁君,郭爱,朱文斌,徐开达,刘连为,王皓学
( 1. 浙江海洋大学 水产学院, 浙江 舟山 316022;2. 浙江省海洋水产研究所, 浙江 舟山 316021)
1 引言
舟山近海海域位于长江口以南,杭州湾以东,受冲淡水(长江、钱塘江两大江河)以及来自外海的台湾暖流的强烈影响,水域环境复杂多变。水团交汇带来大量营养盐,为该海域群落结构产生显著影响[1-8]。尤其是,浮游动物具有随波逐流的生活方式,使之对海洋中水团的变化非常敏感[9]。因此,研究浮游动物群落结构与水团的关系具有重要的意义,不仅对分析浮游动物群落结构、数量分布以及其对海洋环境变化的响应具有科学意义,也可为海洋生态系统结构和功能的修复提供重要的参考。
目前,关于舟山近海的浮游动物群落结构与时空分布有较多的研究报道[6-10]。这些研究大多集中在浮游动物种类组成、数量空间分布、多样性特征及其影响因素,而对于舟山近海水团动态变化对浮游动物群落结构的影响方面的研究较少。本文根据2018-2019年春季在舟山近海调查所获得的浮游动物和水团数据,研究了该海域浮游动物的种类组成、数量分布以及与水团的关系,探讨浮游动物群落结构年际变化的主要原因,以期为舟山近海生态环境评价以及生态系统修复提供基础资料和科学依据。
2 材料与方法
2018年4月8-22日和2019年4月11-23日(春季),搭乘“浙普渔43019”进行两个航次的浮游动物生态调查,调查区域为 29°~32°N,121°~125°E,调查站位分别为41个和37个,近岸平均水深约8~10 m,近海约15~60 m(图1)。浮游动物样品采集均采用浅水I型浮游生物网(口径50 cm、筛绢CQ14,网长145 cm、孔径0.505 mm)进行,根据网口流量计计算滤水体积,所获样品经5%福尔马林溶液固定带回实验室后,去除样品中的杂物,通过显微镜镜检进行分类鉴定和计数。将每个站位的浮游动物统一换算为单位水体中的丰度(ind./m3)。使用温盐深仪(CTD)同步观测水温、盐度和叶绿素a浓度。浮游动物样品的采集、保存、运输和分析均按照《海洋调查规范:海洋生物调查》(GB/T 12763.6-2007)中的规定执行。
2.1 数据处理
2.1.1 浮游动物类群划分
本文采取张冬融[11]对浮游动物分类的方法,将鉴定的浮游动物主要划分为桡足类、毛颚类、水母类、糠虾类、磷虾类、十足类、浮游幼体、夜光虫与其他类共9个浮游动物类群(表1)。夜光虫虽为浮游植物甲藻门,但它能够摄食浮游生物、硅藻甚至鱼卵。最近10多年,夜光虫在东海暴发频繁,对浮游生物生态系统影响巨大[12-14],本文考虑将其作为一种特殊的浮游生物,可以与浮游动物对等,在分类中将它单独作为一类进行研究。所鉴定物种如端足类、多毛类、介形类、海樽类等在所有站位中出现频率低于5%,本文将其统归为一类,即其他类进行研究。为降低浮游动物数据的极化程度,减少稀有种的影响,对浮游动物丰度采用对数转化,,N为原始丰度,Yi为转换后数值。
2.1.2 浮游动物丰度和优势度
浮游动物丰度(N)计算公式为

式中,N为单位体积海水中浮游动物的个体数(ind./m3);V为采集浮游动物样品过滤的海水体积(m3);n为样品鉴定计数获得的浮游动物个体数;R1为浮游动物采样网出水时的流量计读数;R2为浮游动物采样网下水前流量计读数。
浮游动物优势度(Y)计算公式为


图1 2018年(a)和 2019年(b)春季调查站位Fig.1 Sampling stations of spring 2018 (a) and spring 2019 (b)

表1 舟山近海海域浮游动物种类组成Table1 Composition of zooplankton in the Zhoushan nearshore waters
式中,N为各采样站位所有种类的丰度;ni为第i种的丰度;fi为该种在各站位出现的频率。根据以往学者的观点,本文将Y大于0.02的种类定为优势种。环境因子空间分布图、浮游动物类群丰度分布图等均使用海洋数据视图软件(Ocean Date View)绘制。
2.2 统计分析
根据每个站位的表层温度(Sea Surface Temperature,SST)和表层盐度(Sea Surface Salinity,SSS),利用SPSS19.0软件中的Ward法进行空间站位聚类,据此,进行水团的划分。根据舟山近海水文条件,将该海域划分为杭州湾水团(I区)、舟山本岛上升流水团(II区)、舟山近海水团(III区);利用 Primer5.0 软件中的相似性百分比分析(SIMPER)分析不同浮游动物类群、物种在群落结构中的贡献率。采用单因子相似性分析(ANOSIM)对不同水团间的群落结构的差异显著性进行检验。应用典范对应分析(Canonical Correspondence Analysis, CCA)来研究类群组成及与环境因子关系。使用CCA三变量图,研究物种组成、站位及环境因子之间的关系。显著性检验根据Monte Carlo 检验确定。变量的重要性由变量对应轴之间的相关系数R决定,当|R|≥0.4时,该变量即认为具有重要意义[15]。
3 结果
3.1 水文条件及水团划分
2018-2019年春季 SST、SSS、表层叶绿素(Chlorophyll a,Chl a)浓度都呈现出近海高,近岸低的趋势。2018年春季SST整体较2019年低,2018年舟山近海水团SST为14~16℃,上升流水团 SST为12~13.5℃;2019年舟山近海水团SST为15.3~18.3℃,上升流水团SST为13.4~13.8℃;2018-2019年春季舟山近海水团SSS较高(26~35),杭州湾水团SSS低(<15);2018-2019年春季舟山近海Chl a浓度差异较明显,2018年 Chl a浓度较低(1.5~2.5 mg/m3),而2019 年出现 Chl a 浓度高值(2.5~5 mg/m3)(图 2)。
根据对SST、SSS的聚类分析结果,可以将调查区域分为3个水团:杭州湾内水团、舟山本岛上升流水团、舟山近海水团(图3)。杭州湾水团为低盐水,舟山本岛上升流水团为低温水,舟山近海水团为高温高盐水。
3.2 舟山近海浮游动物群落结构
2018年和2019年分别鉴定浮游动物50种和31种(表1),2018年鉴定物种如中华哲水蚤、五角水母,拿卡箭虫、太平洋磷虾以及桡足类无节幼体,它们分别属于桡足类、水母类、毛颚类、磷虾类、浮游幼体。水母类为2018年春季的绝对优势类群,共鉴定出16种,占浮游动物总种数的32%,桡足类为第2大优势类群,共鉴定出13种,种数比例为26%。另外其他类 10种(20%),浮游幼体 6种(12%),磷虾类2种(4%),毛颚类、糠虾类、十足类各1种(2%)。
与2018年相比,2019年所鉴定出的种数明显减少,桡足类成为绝对优势类群,共鉴定出10种,种数比例为32.26%。水母类仅有4种,种数比例为12.90%;另外6大类群鉴定种数由多到少依次为:其他类6种(19.35%),浮游幼体和十足类各 3种(9.68%),糠虾类2种(6.45%),毛颚类和磷虾类各1种(3.23%)。夜光虫在2019年数量巨大,在近海区域分布广泛(3.23%)。

图2 2018年春季与2019年春季舟山近海海域表层温度(SST)、盐度(SSS)、叶绿素a(Chl a)浓度分布与年际变化Fig.2 Surface temperature, salinity, Chl a concentration distribution in the Zhoushan nearshore waters in spring 2018 and spring 2019

图3 2018 年春季(a)与 2019年春季(b)水团聚类Fig.3 Water mass group clustering of sampling stations in spring 2018 (a) and spring 2019 (b)
水团对春季浮游动物群落结构空间分布格局具有显著影响(Anosim 检验,p<0.05),2018 年春季,Ⅰ区-Ⅱ区、Ⅱ区-Ⅲ区平均相异性分别为48.67%、63.83%,2018年春季所有物种的SIMPER分析表明,引起3个水团区差异的主要贡献种(贡献率>10%)为中华哲水蚤。2019年春季,Ⅰ区-Ⅱ区、Ⅱ区-Ⅲ区平均相异性分别为44.76%、64.91%,引起Ⅰ区与Ⅱ区、Ⅱ区与Ⅲ区差异的主要贡献种为中华哲水蚤(贡献率分别为52.98%、63.10%)。Ⅰ区与Ⅲ区差异的主要贡献种为捷氏歪水蚤(Tortanus derjugini)(贡献率 38.85%)(表2)。水团对浮游动物群落结构年际差异亦具有显著影响(Anosim 检验,p<0.05),Ⅰ区、Ⅱ区、Ⅲ区年际平均相异性分别为:52.21%、50.72%、75.58%。引起2018年和2019年Ⅰ区、Ⅱ区、Ⅲ区以及整个调查区域年际差异的主要贡献种为中华哲水蚤、夜光虫和捷氏歪水蚤(贡献率分别为33.63%、19.15%、12.75%)(表 3)。
3.3 舟山近海优势种及其空间分布格局
2018年春季优势种有中华哲水蚤、拿卡箭虫、小拟哲水蚤、拟长腹剑水蚤(表4)。其中,中华哲水蚤在3个水团区出现频率均较高(图4a),在近海海域出现高值,平均丰度达124.93 ind./m3;拿卡箭虫多分布在舟山近海海域(图4b),平均丰度13.25 ind./m3;小拟哲水蚤和拟长腹剑水蚤分布区域相似,更多的分布在近海海域(图4c,图4d),平均丰度分别为6.58 ind./m3、6.12 ind./m3。与2018年相比,2019年春季水温的升高,长江径流量增大,调查海域优势种发生改变,分别为中华哲水蚤、捷氏歪水蚤、五角水母、夜光虫。中华哲水蚤仅在上升流水团和近海水团出现频率较高(图4e),平均丰度12.28 ind./m3;其次捷氏歪水蚤在杭州湾内较集中(图 4f),平均丰度 8.17 ind./m3;五角水母在上升流水团数量较大(图4g),平均丰度2.46 ind./m3。
2018年总丰度时空分布特征为近海(Ⅲ区)高,近岸(Ⅰ区和Ⅱ区)低(图4i),而2019年分布特征为近海(Ⅲ区)低,近岸(Ⅰ区和Ⅱ区)高(图4j)。春季浮游动物总丰度年间差异显著(Anosim 检验,p<0.05),两年浮游动物总丰度分别为:7 152.78 ind./m3,1 007.43 ind./m3。
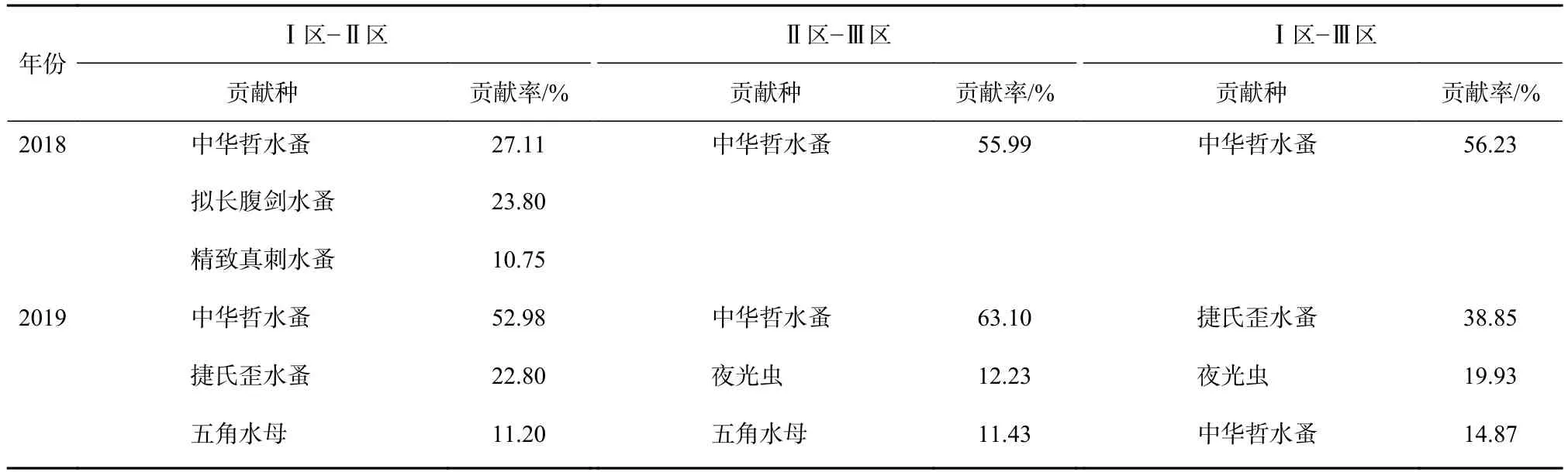
表2 2018年和2019年春季各水团间差异的主要贡献种和贡献率Table2 Main contribution species and contribution rate of differences among water masses in spring 2018 and spring 2019, respectively
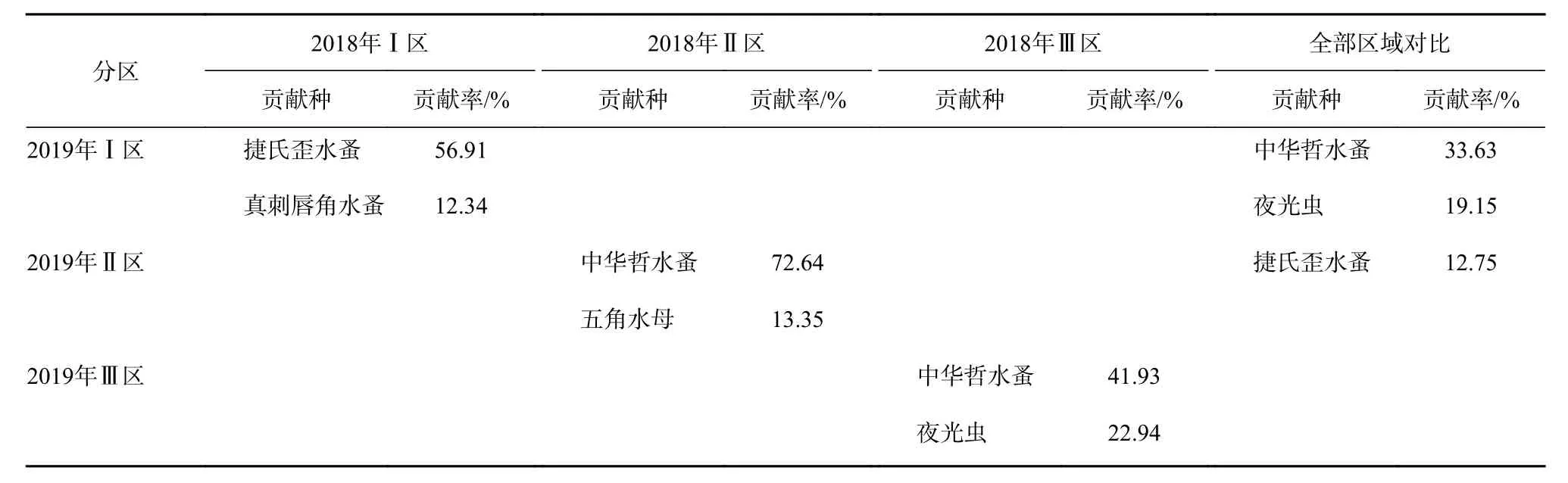
表3 2018年和2019年春季不同水团对浮游动物群落结构组成年际变化和区系格局的影响Table3 The influence of different water masses on the interannual variation and floristic pattern of zooplankton community in spring 2018 and spring 2019

表4 2018年和2019年春季优势种和优势度Table4 Dominant species and their dominance in spring 2018 and spring 2019
3.4 舟山近海浮游动物春季优势种数量年际变化与水团及环境因子的关系
2018年春季与2019年春季各优势种丰度和环境变量的CCA分析结果显示(图5),前4轴与环境变量之间的相关性显著(F=10.7,p<0.01),第1CCA轴(CCA1,特征值:0.44)和第2CCA 轴(CCA2,特征值:0.37)表现出很高的物种-环境相关性,CCA1和CCA2解释了物种累计方差百分比的46.14%和物种-环境累计方差总百分比的97.82%,Chl a浓度、海底层温度(SBT)、SST、海底层盐度(SBS)和 SSS聚集在CCA1的负轴,与CCA1呈负相关。CCA1主要反映了近海水团和湾内水团的差异。CCA2正负轴两边呈现较好的2018年站位和2019年站位的年际差异。捷氏歪水蚤聚集在杭州湾内水团(CCA1的正轴),与SSS、SBS、SST及SBT呈现明显的负相关性,说明SSS、SST是影响其分布水域的主要因素。同时,捷氏歪水蚤在2019年呈现高值,说明冲淡水势力较强,湾内SSS较低,导致该河口种类在2019年春季聚集。同时,优势种夜光虫聚集在CCA2的正轴,说明2019年春季的高SST、高Chl a浓度是影响夜光虫分布的主要因素。而中华哲水蚤、拟长腹剑水蚤、小拟哲水蚤、拿卡箭虫和五角水母则聚集在近海海域,说明暖温环境是影响其分布的主要因素。这些分隔模式可能反映了浮游动物对栖息地类型的偏好或者其生态习性的适应性。

图4 2018年和2019年春季浮游动物优势种分布Fig.4 Distribution of dominant zooplankton species in spring 2018 and spring 2019
4 讨论
4.1 春季浮游动物优势种的年际变化与水团的关系

图5 浮游动物优势种与环境因子典范对应分析Fig.5 Correspondence analysis between dominant zooplankton species and environmental factors
本研究结果显示,2018年各水团区域浮游动物丰度由高到低依次为舟山近海水团(平均332.37 ind./m3)、上升流水团(平均11.11 ind./m3)、杭州湾水团(平均10.17 ind./m3)。无论是物种组成还是类群组成,3个划分区域所有浮游动物组成均具有显著差异(表2),此外,在近海海域,近海种如中华哲水蚤、拿卡箭虫丰度最高,显示出近海水团具有适合近海种栖息的环境。这与以往研究[15-18]相一致。春季水温的回升,有利于浮游植物的生长繁殖,不仅为浮游动物提供了丰富的饵料,而且为浮游动物摄食浮游植物提供了较好的环境条件[19-21]。
然而,与2018年春季的分布趋势相反,2019年春季各水团区域浮游动物丰度由高到低依次为:上升流水团(平均 57.52 ind./m3)、杭州湾水团(平均 44.75 ind./m3)、舟山近海水团(平均22.07 ind./m3),相关分析也表明,2019年春季研究海域浮游动物丰度与水温和盐度呈显著负相关(p<0.05)。造成这一差异的主要原因可能为2019年春季近海夜光虫暴发,大量聚集在近海高Chl a浓度区域,侵占浮游动物原本的生存空间[12-13],导致其他浮游动物如中华哲水蚤、捷氏歪水蚤在其他海域(杭州湾水团、上升流水团)聚集,最终影响浮游动物的分布格局或生物量,这一结果与胡翠林等[14]的研究类似。
2018年春季长江冲淡水径流量(19 823.64 m3/s),显著小于2019年(26 387.1 m3/s),本文猜测冲淡水的差异可能会对浮游动物的优势种的数量变化与空间分布格局造成显著的影响[22-23]。2018年春季浮游动物大都分布于舟山近海水团,而较少在杭州湾内水团出现,这可能与冲淡水量较小导致的杭州湾内营养盐的总体水平较低有关。2019年较大的冲淡水量,引起了较多的浮游动物在杭州湾内聚集,同时,舟山近海发生了较强的浮游植物藻华,也引起了部分浮游动物(比如夜光虫)在近海聚集。因此,2018年和2019年的浮游动物空间分布格局主要体现了饵料对生物聚集的调控。周孔霖[24]也发现了饵料浓度影响浮游动物的生长和聚集,本文的研究结论与其结果一致。
4.2 浮游动物总生物量年际差异的原因
本文结果显示,2018年春季SSS高、SST低、Chl a浓度低,而浮游动物总丰度大,与此相反,2019年SSS较低、SST较高、Chl a浓度较高,而浮游动物总丰度低(图2,图4)。根据食物匹配和温度有利于生理生长的理论[25-28],2018年的浮游动物总丰度应该较2019年低。本文的研究结果与常规的理论推测相反,造成这一现象可能与两个因素有关。
一是2019年春季长江冲淡水量异常偏高,为有害赤潮藻暴发提供了丰富的营养盐,导致适宜生长的夜光虫暴发。夜光虫虽然本身不含毒素,但其暴发后,导致其他浮游生物生存环境更加恶劣(比如缺氧、挤占生存空间、浮游动物适宜的饵料数量减少),从而导致2019年春季高营养物质流入有害赤潮藻类,导致大量有害浮游植物不能被浮游动物有效利用,造成浮游动物数量偏低。很多研究[14,29-30]也表明,发生赤潮藻华时,浮游动物生物量不升反降,与本文的研究结果一致。
二是气候条件所致,2018年春季呈现明显的拉尼娜现象[31],温度总体偏低,可能导致偏冷水性浮游动物数量较多,而偏暖温性浮游动物数量较少,亦有研究(http://www.bom.gov.au/climate/enso/)表明,2019年春季呈现厄尔尼诺现象[31],导致原来栖息于此的偏冷水性浮游动物大量死亡,从而导致总的浮游动物数量偏低。如果这两年推论成立,本研究结果表明,人类活动和气候变化会对浮游动物种群数量变化产生显著的影响,对优势种的交替具有重要的影响。对于本研究推论的广泛适用性,还需要结合其他年份的调查数据,进一步的调查验证。
5 结论
本研究根据2018-2019年春季的浮游动物丰度数据和环境因子数据,发现舟山近海海域存在3个水团:杭州湾内水团、舟山本岛上升流水团和舟山近海水团。分析发现,不同水团对春季浮游动物群落结构地理分布格局影响显著,且对浮游动物群落结构的年际变化具有显著影响,盐度是影响春季浮游动物群落结构空间差异的主要因素,温度、叶绿素a浓度是春季浮游动物群落结构年际差异的主要因素。其次,夜光虫等浮游生物的暴发对浮游动物分布会造成重要影响。
致谢:感谢“浙普渔43 019”号全体船员在出海采样过程中的帮助,A17海洋资源与环境班级董军、官道伊、江丹妮、张晗、龙丽梅等同学参与部分实验,谨致谢忱。
猜你喜欢
大气科学学报(2022年2期)2022-05-12
华北水利水电大学学报(自然科学版)(2021年4期)2021-10-08
空间科学学报(2021年2期)2021-07-21
辽宁化工(2021年5期)2021-06-03
——以“不同咖啡因饮料对水蚤心率的影响”为例
中学生物学(2021年2期)2021-04-30
科技视界(2020年2期)2020-03-23
铁道通信信号(2020年8期)2020-02-06
海洋与湖沼(2020年1期)2020-02-06
作文小学中年级(2019年10期)2019-11-04
中国海洋大学学报(自然科学版)(2019年9期)2019-01-04
