
铁观音茶树甘油-3-磷酸酰基转移酶(CsGPAT)基因克隆及萎凋过程中的表达分析
2020-07-20 02:44毕婉君周子维武清杨郑玉成倪子鑫柳镇章
福建农业学报 2020年4期
毕婉君,周子维,武清杨,郑玉成,倪子鑫,柳镇章,孙 云
(福建农林大学园艺学院,福建 福州 350002)
0 引言
【研究意义】铁观音(Camellia sinensis var. sinensis cv.Tieguanyin)原产福建省泉州市安溪县西坪镇,是国家优良茶树品种[1-2];其抗性强,适制乌龙茶,香气高长持久,具有独特的兰花香,是中国十大名茶之一。铁观音中的脂肪族类香气物质是组成其独特香气的成分之一[3],茶叶加工过程中合成脂肪族类香气物质的前体物质是以亚油酸、亚麻酸为代表的不饱和脂肪酸。不饱和脂肪酸是磷酸的组成部分,磷脂类物质是甘油磷酸代谢途径(Glycerophospholipid metabolism)的最终产物。甘油磷脂代谢过程有许多酶的参与,其中甘油-3-磷酸酰基转移酶(Glycerol-3-phosphate acyltransferase,GPAT)是参与反应的第一个转移酶[4],它催化酰基辅酶A上的脂肪酸与sn-甘油结合,形成1-酰基-sn-甘油-3-磷酸[5-7]。此外,GPAT基因是植物中的抗性基因,低温胁迫使其表达量增加,一定程度上影响细胞膜上的不饱和脂肪酸含量[8-9]。不饱和脂肪酸的含量和膜脂流动性成正比,膜脂流动性增强使得植物抵御寒冷的能力增强[10-11]。萎凋是乌龙茶加工的关键步骤,相对低温萎凋会促使茶叶香气物质形成,从而影响茶叶品质[12]。【前人研究进展】目前,GPAT基因已在拟南芥[13]、油茶[14]、油菜[15]、油桐[16]、番茄[17]等多种植物中克隆。在模式生物拟南芥中已经发现了10个AtGPAT基因家族成员,它们各自参与脂质生物合成的途径[18-19];陈丽静等[20]以王百合为试验材料,发现GPAT基因表达量随着冷诱导时间增加而增加,表明GPAT基因和植物抗寒性相关;皮广静等[21]对花生油脂合成进行探究,发现花生含油量与GPAT基因3个时期平均表达量呈显著性相关,表明GPAT基因和油脂合成相关;在拟南芥花粉发育时期,AtGPAT1/2基因共同影响拟南芥的结实率[22],表明GPAT基因在植物育种中有重要意义。【本研究切入点】铁观音作为我国优良茶树品种,成品茶的香气馥郁芬芳,GPAT基因是合成脂肪类香气途径的上游基因,虽然GPAT基因已经在许多植物中克隆,并证实其在植物抗寒性、油脂合成、育种中有重要作用,但是在茶树中的相关研究还很少,尤其是在茶叶加工过程中表达量分析的研究鲜见报道。【拟解决的关键问题】本试验用铁观音萎凋叶为试验材料,通过克隆CsGPAT基因,分析其在不同温度下的表达量,以期能为茶树萎凋工艺中温度调控提供理论依据,为抗性基因在铁观音茶叶萎凋工艺过程中的实际应用提供重要参考。
1 材料与方法
1.1 材料
1.1.1 试验材料 2018年4月份,在福建农林大学南区教学茶场,选取3~4片小开面完整并且无病虫害的铁观音茶树叶片进行不同温度萎凋处理。鲜叶萎凋在不同温度的恒湿箱中进行处理,温度分别是20℃、30℃、40℃,记做T20、T30、T40,控制萎凋湿度为60 %,摊叶厚度10 mm,萎凋时间60 min,萎凋叶光泽消失,叶色转暗绿色,发出轻微青草香,含水率为(68.4±1.3)%。处理后用锡箔纸进行密封,用液氮速冻法进行处理,放置-80℃储藏,用于后续RNA提取。
1.1.2 试验主要试剂 75 %乙醇(DEPC-H2O配置),交联聚乙烯吡咯烷酮(PVPP),Golden DNA聚合酶,pMD18-T载体,DH-5α感受态细胞,DreamTaqGreen酶,GelGreen荧光核酸凝胶染色试剂,D2000 DNA Ladder,TIANGEN公司的多糖多酚植物总RNA提取试剂盒,引物委托厦门闽博生物技术有限公司合成,测序委托铂尚生物技术(上海)有限公司完成。
1.1.3 试验主要仪器 移液器、微量台式离心机、高速冷冻离心机、超微量核酸检测仪、电泳仪、凝胶成像仪、BIO-RAD PCR扩增仪、OSE-PRO程控金属浴、超净工作台、Roche Light Cycler®480 PCR实时荧光定量仪、制冰机。
1.2 基因克隆方法
1.2.1 总RNA的提取及质量检测 取铁观音T20、T30、T40处理萎凋后无病虫害的完整叶片为试验材料,按照TIANGEN公司的多糖多酚植物总RNA提取试剂盒步骤,提取铁观音茶树总RNA。用1%琼脂糖凝胶对其进行电泳检测,确定其完整性,然后用超微量核酸检测仪对其OD值进行检测,选择符合试验结果的RNA用于后续试验。
1.2.2 RNA反转录 以符合要求的总RNA为试验材料,采用GeneRacer Kit试剂盒的方法合成cDNA第一链。参照林玉玲[23]逆转录的方法,第一步是RNA的去磷酸化;第二步是mRNA去帽子结构;第三步是RNA接头链接;第四步是反转录。
1.2.3 PCR引物设计 根据已知序列设计3′RACE和5′RACE引物(表1),用于PCR巢式扩增试验。基于测序获得甘油-3-磷酸酰基转移酶(GPAT)基因ORF序列,利用DNAMAN 6.0软件选择具有差异的区域设计保守区的特异性上下游引物,用于RT-qRCR试验(表2)。

表1 3′RACE和5′RACE所使用的引物Table1 Primers used in 3′ RACE and 5′ RACE

表2 实时荧光定量基因表达分析的引物序列Table2 Primer sequence for real-time fluorescence quantitative gene expression analysis
1.2.4 目的片段的扩增、回收、克隆与测序 用获得的cDNA第一链为模板,GPAT-F、GPAT-R为引物,通过常规的PCR扩增技术获得铁观音茶树的GPAT基因目的片段。用购自TaKaRa公司的SYBR Premix ExTaqTM(Code NO.RR716)试剂盒,并且配合使用Roche LightCycler®480实时荧光定量PCR仪,严格按照其步骤,进行qRT-PCR。qRT-PCR反应体系为20 μL,配置用量:10 μL 2×SYBR Premix ExTaq、0.8 μL 200 nmol·L-1的上下游引物、2 μL cDNA 模板、6.4 μL ddH2O。qRT-PCR反应程序(两步法)为:94℃预变性5 min;94℃变性1 min;53℃退火40 s;72℃延伸45 s,共45个循环,72℃延伸10 min。得到目的产物进行电泳鉴定。采用 TaKaRa公司提供的试剂盒上的实验步骤回收目的条带,将得到的目的条带连接到pMD-18 T载体上,对其做PCR扩增检验,交由铂尚生物技术(上海)有限公司进行测序。
1.2.5 目的基因表达量定量测定 反应结束后进行扩增曲线和熔解曲线分析,标准曲线每个梯度均进行3次重复,其中 cDNA模板为各个样本cDNA混合样的5倍梯度稀释,以灭菌蒸馏水作为阴性对照。
采用qRT-PCR仪采集的数据做熔解曲线。选用单内参方法,选用CsEF-1α为内参基因,设计引物如表2所示,用于实行荧光定量PCR[24]。对比CK样品的数据,使用 Excel办公工具,通过2-ΔΔCt公式计算铁观音茶树叶中影响低温胁迫的目的基因(GPAT)在不同萎凋温度时的相对表达量。接着使用DNAman软件设计目的基因的特异性引物见表1。
1.2.6 多序列比对和系统发育分析 利用NCBI(http://www.ncbi.nlm.nih.gov)网站的BlastP工具对克隆得到的CsGPAT基因进行氨基酸序列的相似性和同源性查找,并利用这些序列进行氨基酸序列同源性比较。然后将这些序列下载下来保存为fasta格式,采用MEGA-X软件进行系统发生和进化分析,系统发育树采用Neighbour-Joining(距离邻接法)构建。
1.2.7 序列分析及生物信息学分析 编码蛋白质的基本理化性质预测利用ExPASy Protparam(http://www.expasy.ch/tools/protparam.htmL)分析完成;蛋白质结构功能域预测通过SMART(http://smart.emblheidelberg.de/)分析完成;亚细胞定位情况预测分别通过SignalP 4.1Server(http://www.cbs.dtu.dk/ services/SignalP/)和 PSORT(http://psort.nibb.ac.jp/)以 及prediction protein(https://www.predictprotein.org/home)分析完成;蛋白质二级结构预测通过prediction protein(https://www.predictprotein.org/home)完成;蛋白质三维结构预测通过SWISS-MODEL在线工具(http://www.swissmodel.expasy.org/)完成。
2 结果与分析
2.1 铁观音茶树总RNA质量的检测和cDNA扩增序列的获取
用1%琼脂糖凝胶对得到的铁观音茶树总RNA做电泳测定,测得结果显示,总RNA条带无缺失、无拖尾现象,表明RNA基本无降解。用超微量核酸检测仪检测其OD值判断其纯度,OD260/280在1.8-2.2范围内,表明提取的总RNA纯度较高,达到试验标准。测序表明,GPAT基因的5′-RACE PCR产物长度约为100 bp,3′-RACE PCR长度约为250 bp(图1)。用获得的 cDNA第一链为模板,进行常规PCR技术扩增铁观音茶树GPAT基因中间保守区,获得在1 554 bp的基因片段(图1)。
2.2 甘油-3-磷酸酰基转移酶基因编码的蛋白质生物信息学分析
2.2.1 甘油-3-磷酸酰基转移酶蛋白质基本理化性质分析 通过ExPASy Protparam预测网站,对该蛋白质序列进行基本理化性质分析,并得出结论。该蛋白质相对分子质量为55.15 kD,理论等电点(pI)的值为9.3,发现原子构成为 C2519H4025N655O691S19,原子总个数为7 909,共有108个带电荷氨基酸,其中有46个氨基酸(Asp+Glu)带有负电荷,剩下62个氨基酸(Arg+Lys)带正电荷,蛋白质序列不稳定系数为36.49,表明该蛋白质为稳定蛋白。对其蛋白质在不同部位的半衰期分析后发现,在哺乳动物红细胞、酵母细胞、大肠杆菌细胞中的半衰期分别为30 h、20 h和10 h。总平均亲水性(GRAVY)0.123,为疏水性蛋白。
2.2.2 甘油-3-磷酸酰基转移酶蛋白质结构功能域预测与分析 通过预测分析后得出:GPAT蛋白质序列中一共含有3个功能域,分别是pfam域、跨膜螺旋区域和SMART PlsC域。其中pfam域处于23~203位氨基酸,跨膜螺旋区域处于242~264位氨基酸,SMART PlsC域处于301~402位氨基酸。SMART PlsC域又名为磷酸酰基转移酶,具有磷脂生物合成功能,具有磷酸甘油、1-酰基甘油磷酸酯或2-酰基甘油磷酸乙醇氨酰基转移酶活性。所以SMART PlsC域是甘油-3-磷酸酰基转移酶基因的主要功能区域,也是该基因重要组成部分,合成磷脂生物是PlsC域的本质特征,因此该蛋白具有磷脂生物合成功能。
2.2.3 甘油-3-磷酸酰基转移酶蛋白质信号肽与亚细胞定位分析 对GPAT蛋白的结构运用SignalP 4.1 Server进行预测,结果如图2所示。信号肽是一段特殊的氨基酸序列,其功能是在新合成的多肽链中,对蛋白质的跨膜转移进行有效指导。蛋白质信号肽及其断裂位点预测结果显示该蛋白质不含信号肽(图2)。亚细胞定位对生物学功能有一定影响,通过PSORT(http://psort.nibb.ac.jp/)软件进行预测,结果显示,对GPAT蛋白质在细胞中定位分析后,得出其在内质网上的定位分值为44.4 %、质膜定位分值为22.2 %、液泡、线粒体、高尔基体的定位分值为11.1 %,表明其很有可能位于内质网中。
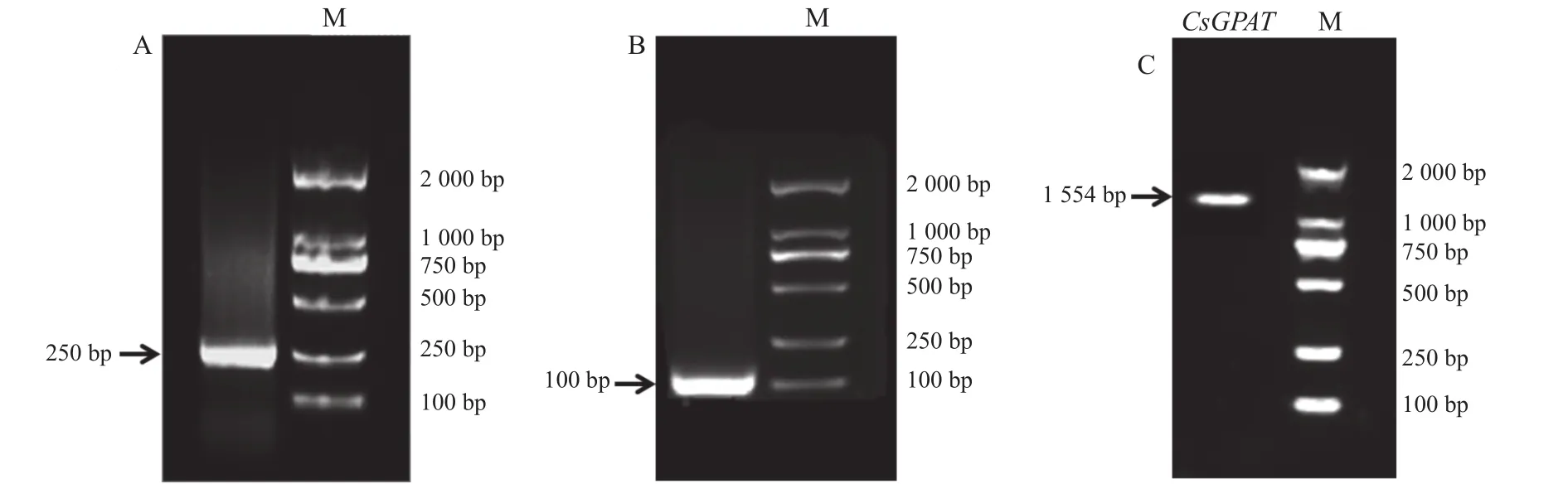
图1 PCR产物凝胶电泳分析Fig.1 Gel electrophoresis analysis on PCR products

图2 甘油-3-磷酸酰基转移酶蛋白质的信号肽预测结果Fig.2 Prediction of signal peptide in GPAT protein
2.2.4 甘油-3-磷酸酰基转移酶蛋白质三维结构预测与分析 对GPAT蛋白质的三维结构运用prediction protein软件进行预测,从而使得可以更加直接清晰地理解GPAT基因编码的蛋白质结构在铁观音茶叶中的存在情况。结果表明多肽链中有规则构象主要有两种,分别是α-螺旋和β-折叠。α-螺旋、β-折叠的比例分别为39.03 %和16.50 %。其还包含无规则卷曲状态,所占比例为44.47 %。通过SWISS-MODEL在线工具对其进行蛋白质三维结构预测进一步说明蛋白质的功能(图3)。
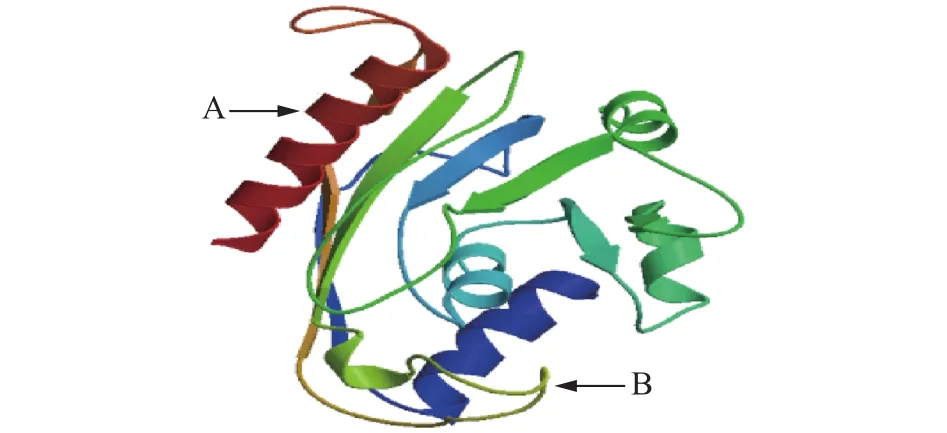
图3 甘油-3-磷酸酰基转移酶蛋白质三维结构预测Fig.3 Predicted 3D structure of GPAT protein
2.2.5 甘油-3-磷酸酰基转移酶蛋白质同源性分析 对铁观音茶叶中检测得到的GPAT基因编码得到的氨基酸序列做同源性分析,在 GenBank库中寻找筛选出其他植物的GPAT基因编码的氨基酸序列与铁观音物种的GPAT氨基酸序列进行比较。结果表明,CsGPAT氨基酸序列与胡杨(Populus euphratica)、毛果杨(Populus trichocarpa)、中华猕猴桃(Actinidia chinenesis var. chinensis) 、棉花(Gossypium hirsutum)的GPAT氨基酸序列相似度较高,分别是87.53 %、86.72 %、87.55 %、86.52 %。进化树通过GPAT蛋白序列组成,系统进化树显示铁观音茶树GPAT与油茶(Camellia oleifera)的亲缘关系最近(图4)。
2.3 铁观音不同温度萎凋过程中CsGPAT基因的表达分析

图4 NJ法构建的GPAT氨基酸序列的系统进化树Fig.4 Phylogenetic tree of GPAT amino acid sequence constructed by NJ method
通过实时荧光定量PCR技术,对铁观音萎凋过程中茶叶甘油-3-磷酸酰基转移酶基因的相对表达量变化进行检测。分析结果显示在铁观音萎凋过程中,CsGPAT基因的相对表达量在萎凋温度20℃时达到最高,当萎凋温度达到30℃和40℃时,该基因表达量被显著抑制表达(图5)。对CsGPAT基因的相对表达量用SPSS进行差异性分析,结果表明室温表达量和20℃、30℃、40℃萎凋表达量存在极显著差异,20℃萎凋表达量和30℃、40℃萎凋表达量也存在极显著差异,30℃和40℃萎凋表达量之间不存在差异性。表明CsGPAT基因的相对表达量与温度密切相关,相对低温环境会促进CsGPAT基因表达。

图5 甘油-3-磷酸酰基转移酶基因在不同萎凋温度下的相对表达量Fig.5 Relative expressions of CsGPAT at different withering temperatures
3 讨论与结论
本研究通过巢式PCR克隆得到1个长度为1 554 bp的铁观音茶树甘油-3-磷酸酰基转移酶基因(GPAT),编码497个氨基酸。GPAT的结构特征是具有磷脂生物合成功能的SMART PlsC结构域,大部分植物GPAT具有这种结构域,说明GPAT基因与磷脂生物合成密切相关。GPAT是甘油磷脂代谢途径的第一个关键转移酶,最终合成磷脂类物质是不饱和脂肪酸香气的前体物质。在植物受到逆境胁迫时,GPAT基因的表达量会随着温度下降而上调,说明CsGPAT与温度调控存在重要关系,同时可能对于茶叶加工过程中香气形成有一定作用。甘油-3-磷酸酰基转移酶基因家族具有Phospholipid/glycerol acyltransferase蛋白质保守结构域,在茶树(阿萨姆种,CSA)基因组中共有20个拷贝数(IPR002123),在茶树(中国种,CSS)中则存在9个拷贝数(EC:2.3.1.15)。本研究基于茶叶萎凋过程中的转录组数据,筛选获得了一条GPAT相关基因(IDcsa:CSA000941.1/IDcss:TEA019813.1)。目前,茶树基因组数据库已经公布(http://tpia.teaplant.org/),显示低温胁迫会使GPAT基因表达量上调。
在茶叶加工中,不饱和脂肪酸经脂肪氧合酶途径合成大量脂肪族类香气物质,比如,正己醛、青叶醛、青叶醇、乙酸乙酯、茉莉酸甲酯等[25],最终影响茶叶品质的形成。铁观音具有的独特花香主要由脂类、醇类、醛类挥发性物质组成[1],GPAT基因促进此类香气的前体物质产生,表明GPAT基因可能作用于铁观音香气的形成。GPAT基因在铁观音不同萎凋温度过程中的表达量分析表明,铁观音GPAT基因表达量随着萎凋温度的降低而上调,在萎凋温度20℃时表达量最高,推测在茶叶萎凋过程中,叶片受到相对低温胁迫诱导基因表达,有利于脂肪族类香气物质的形成。目前,拟南芥中已鉴定出有10个GPAT家族成员,以拟南芥10个GPAT家族成员蛋白(http://tpia.teaplant.org/#)为种子序列,与本试验克隆的CsGPAT构建系统进化树,分析显示CsGPAT与AtGPAT4和AtGPAT8的同源性高。前人研究表明AtGPAT4与AtGPAT8与植物角质的形成相关,它们存在酰基转移酶结构域,还同时存在磷酸化酶结构域[26]。利用在酵母中GPAT-缺失突变体(gatl delta)和/或麦胚无细胞转移系统进行表达验证GPAT4催化反应,表明这个酰基转移酶酯化偏爱酰化基团在G3P的sn-2位,并认为酰基转移和磷酸酶的双功能性可能参与角质组成单体的组合。植物角质层是植物细胞与外界环境隔离的一道天然屏障,因此过表达GPAT,可能会提高植物的抗逆表现。由此推测茶树中CsGPAT基因极有可能具有此功能,在茶叶的抗逆性中发挥重要作用。后续研究将通过基因共表达分析,进一步探究GPAT基因在冷胁迫条件下的表达模式,构建GPAT基因的调控网络。
抗逆性不仅对茶树的生长发育十分重要,同时在茶叶加工过程中,茶叶在萎凋中受到相对低温时的防御反应会影响茶叶香气的形成,促使茶叶香气物质的增加[27-29],提高茶叶品质,从而增加茶叶经济效益。本研究通过克隆和分析GPAT基因在萎凋过程中的表达分析,初步确认GPAT基因与茶叶加工过程中脂肪族类香气物质合成密切相关,铁观音GPAT基因在不同萎凋温度中响应表达,为后续研究甘油-3-磷酸酰基转移酶基因在低温响应中对茶叶香气形成的调节机制方面提供了理论参考。GPAT基因表达可能受温度调控,对茶叶萎凋过程中脂肪族类香气物质的形成发挥重要作用。
猜你喜欢
保健与生活(2022年11期)2022-06-09
少年文艺·我爱写作文(2021年5期)2021-07-20
今日农业(2020年18期)2020-12-14
生物学通报(2020年10期)2020-08-13
海峡姐妹(2019年11期)2019-12-23
天然产物研究与开发(2018年8期)2018-09-10
能源(2017年7期)2018-01-19
中学科技(2017年11期)2017-12-26
广州化工(2016年8期)2016-09-02
国际妇产科学杂志(2016年2期)2016-06-16
