
少花蒺藜草对科尔沁沙地土壤氮转化效应的研究
2020-07-16 14:02王然张婷宋振张瑞海付卫东张国良
生态环境学报 2020年4期
王然,张婷,宋振,张瑞海,付卫东,张国良
中国农业科学院农业环境与可持续发展研究所,北京 100081
外来入侵植物入侵后,会对当地生态系统的完整性和生物多样性造成严重威胁,同时破坏原始生态系统的养分循环。(Crook,2002;Castro-Díez et al.,2014)氮是陆地生态系统限制生长的关键养分因子,影响着生态系统的生态过程。土壤氮矿化是土壤有机氮在微生物的作用下转化为无机态氮的过程(Abera et al.,2012),是陆地生态系统氮循环的关键过程,很大程度上决定了陆地生态系统初级生产力(陈静等,2016)。有研究表明,外来入侵植物通过引起当地土壤氮库转变(Fei et al.,2014),改变各种形态氮素的分配和周转(Laungani et al.,2009)来影响土壤氮循环(Piper et al.,2015)。如:加拿大一枝黄花(Solidago canadensis)入侵后土壤总氮库增大,而铵态氮库和硝态氮库减小(陆建忠等,2005);紫茎泽兰(Eupatorium adenophora)入侵使土壤中总氮、硝态氮、铵态氮含量均显著增加(于兴军等,2005;牛红榜等,2007);火炬树(Rhus typhina)入侵黑松幼林增加了土壤硝态氮,降低了土壤铵态氮,而对全氮没有影响(黄乔乔等,2013);黄顶菊(Flaveria bidentis)入侵对土壤氮素养分的影响在不同地区间差异较大(李博等,2006;赵晓红等,2015)。
科尔沁沙质草地位于中国北方半干旱农牧交错带,荒漠化严重(李玉霖等,2013),植被退化和生物多样性丧失严重(蒋德明等,2002),极易受外界干扰(陈静等,2016)。持续的全球环境变化影响着氮沉降率(Hejda et al.,2017),同时由大气氮沉降等方式引起的外源氮增加和氮素形式的变化,会影响输入陆生态系统的氮素质量和数量进而影响氮矿化作用(Gao et al.,2013;Hobbie,2008)。因此,研究外源氮对土壤氮矿化作用的影响非常必要。
少花蒺藜草(Censhrus Spinfex)为20世纪80年代入侵科尔沁沙质草地的一年生恶性杂草,具有极强的耐旱、耐贫瘠等特性,可在短时间内形成单一优势群落。目前关于少花蒺藜草的研究主要集中其传播与危害(杜广明等,1995;张衍雷,2015)、生物学特性(徐军等,2012;田迅等,2015;周立业等,2014)防治技术(王坤芳等,2013;付卫东等,2018)等方面,对少花蒺藜草入侵的机制研究较少。此前已有研究表明少花蒺藜草入侵后改变了本地土壤氮库的结构(张婷等,2017),但对于土壤氮矿化的影响尚未见报道。因此,本研究以少花蒺藜草为研究对象,通过室内培养试验,分析少花蒺藜草与添加外源氮对科尔沁沙质草地土壤氮矿化的影响及其交互作用,为揭示少花蒺藜草入侵生态学机制提供理论基础。
1 材料与方法
1.1 研究区概况
研究区位于科尔沁沙地南缘的辽宁省彰武县阿尔乡白音花农场,海拔230—250 m。属温带季风性气候,年均温5.7 ℃,有效积温2800—3200 ℃,平均无霜期154 d。年降水量450—550 mm,多集中在 7—8月,年均蒸发量 1570 mm,年均湿度60%—65%(张衍雷等,2015)。土壤类型为风沙土,表土有机质含量1%—1.5%,保水保肥能力差。该区域少花蒺藜草危害严重,伴生植物主要为鹅观草(Roegneria kamoji)、绿珠藜(Chenopodium acuminatum)、马唐(Digitaria sanguinalis)等。
1.2 试验设计与样品采集
在研究区内选择3种类型样地:(1)裸地:样地内无植被生长;(2)鹅观草样地:样地内仅生长本地植物鹅观草,无人为干扰,长期生长,覆盖度为40%—70%,无少花蒺藜草;(3)少花蒺藜草入侵样地:样地内生长着少花蒺藜草,入侵时间5年以上,覆盖度为70%—100%。各样地相距200—300 m,除植被外,土壤其他条件基本一致。
分别于6月18日、7月23日、8月30日、9月30日及11月2日在各样地内采集供试土壤,在每个样地内按照五点取样法选取5个1 m×1 m的样方,样方间距5 m以上。在每个样方内,采用四分法取0—20 cm土层风干土样,混匀过2 mm筛备用。收割地上部分植株,并收集凋落物作为植物样品,带回实验室备用。
1.3 土壤氮矿化作用测定方法
土壤氮矿化作用测定采用室内培养法(Zhang et al.,2016)。称取过筛后的风干土样30 g,置于250 mL塑料广口瓶中,去离子水调节土壤含水量至20%(质量含水量),用聚乙烯膜包扎瓶口,并在膜上扎 2个小孔,置于 25 ℃培养箱中预培养7 d。预培养后,分成3组:分别添加N 50 mg·kg-1(以烘干土壤计)硫酸铵、硝酸钠溶液,以及等重去离子水(CK)。3组土壤湿度均调整至土壤持水能力的40%。所有培养瓶再次密封,25 ℃黑暗培养14 d。通过称重确定土壤水分变化情况,以去离子水补充土壤水分。在培养的0 d和14 d,加2 mol·L-1KCL溶液75 mL,于温度为25 ℃的恒温振荡机内震荡 1 h,滤液用连续流动分析仪(Auto Analyzer 3 System,SEAL Analytical GmbH公司,英国)测定土壤中硝态氮和铵态氮浓度。
1.4 数据处理
采用Excel 2010和SPSS 19.0软件对数据进行统计分析。验证正态分布后,以添加氮素形式和植被类型作为处理因子进行双因素方差分析,并对所有指标进行单因素方差分析(One-way ANOVA),采用LSD和Tukey法进行多重比较(α=0.05)。用Origin 9.0软件作图。图表中数据为平均值±标准误。
土壤氮素转化速率计算式:

这几种速率所用的单位为 mg·kg-1·d-1。公式中,N0和Nt是培养初始和培养t天后的硝态氮浓度,M0和Mt是培养初始和培养后t天的铵态氮浓度。
2 结果与分析
2.1 少花蒺藜草入侵及外源氮对土壤净氮硝化速率的影响
由图1可以看出,在不添加氮素土样中,净氮硝化速率差异显著,均值大小顺序为裸地 (0.28 mg·kg-1·d-1)>入侵地 (-0.30 mg·kg-1·d-1)>鹅观草样地 (-0.44 mg·kg-1·d-1)。在 6—10 月,土样净硝化速率范围为入侵地 0.78—0.31 mg·kg-1·d-1,鹅观草样地-0.60— -0.34 mg·kg-1·d-1,裸地-0.08—0.84 mg·kg-1·d-1。与鹅观草样地相比,入侵地净硝化速率在6—7月显著增加,10月显著降低。与裸地相比,入侵地净硝化速率在6月显著增加,在7—10月均显著降低。
添加铵态氮显著增加了 3组处理的净硝化速率,增加了21—38倍。均值大小顺序为裸地(11.05 mg·kg-1·d-1)>鹅观草样地 (9.10 mg·kg-1·d-1)>入侵地 (8.66 mg·kg-1·d-1)。在 6—10 月,土样净硝化速率范围为入侵地 5.25—11.71 mg·kg-1·d-1,鹅观草样地 8.05—10.07 mg·kg-1·d-1,裸地 10.29—12.24 mg·kg-1·d-1。与鹅观草样地相比,入侵地净硝化速率在6—7月显著增加,8—10月显著降低。与裸地相比,入侵地净硝化速率在7月显著增加,在8—10月均显著降低。
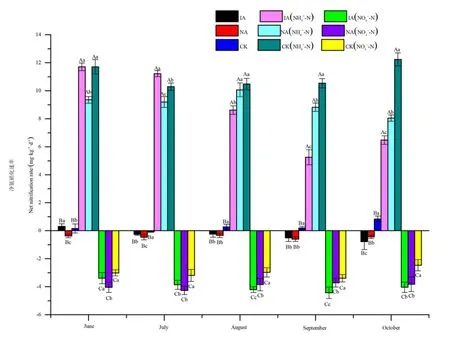
图1 不同植被类型与氮素处理的净硝化速率的月变化Fig.1 Net nitrification rate in different vegetation types and nitrogen treatments at different periods
添加硝态氮显著降低了这3组处理的净硝化速率,降低了7—12倍。均值大小顺序为裸地 (-3.00 mg·kg-1·d-1)>鹅观草样地 (-3.93 mg·kg-1·d-1)>入侵地 (-3.98 mg·kg-1·d-1)。在 6—10 月,土样净硝化速率范围为入侵地-4.43— -3.38 mg·kg-1·d-1,鹅观草样地-4.26— -3.71 mg·kg-1·d-1,裸地-3.39—-2.47 mg·kg-1·d-1。与鹅观草样地相比,入侵地净硝化速率在6月显著增加,8—9月显著降低。与裸地相比,入侵地净硝化速率7—10月均显著降低。
由表1可以看出,少花蒺藜草入侵与外源氮添加均对土壤净氮硝化速率有显著影响,且这两个因素的交互效应显著。其中,与净硝化速率的关联性(R2)的大小次序为,添加外源氮处理>外来植物入侵>交互效应。
2.2 少花蒺藜草入侵与外源氮对土壤净氮氨化速率的影响
由图2可得,在不添加氮素土样中,净氮氨化速率差异显著,均值大小顺序为入侵地 (0.73 mg·kg-1·d-1)>鹅观草样地 (0.57 mg·kg-1·d-1)>裸地(-0.59 mg·kg-1·d-1)。在 6—10 月,土样净硝化速率范围为入侵地-0.12—1.65 mg·kg-1·d-1,鹅观草样地0.01—0.86 mg·kg-1·d-1,裸地-2.56—0.20 mg·kg-1·d-1。与鹅观草样地相比,入侵地净氨化速率在 6月显著降低,8—10月显著增加。与裸地相比,入侵地净氨化速率在7—10月显著增加)。

表1 净氮硝化速率的主体效应检验Table 1 Inter-subjective test of net nitrification rate
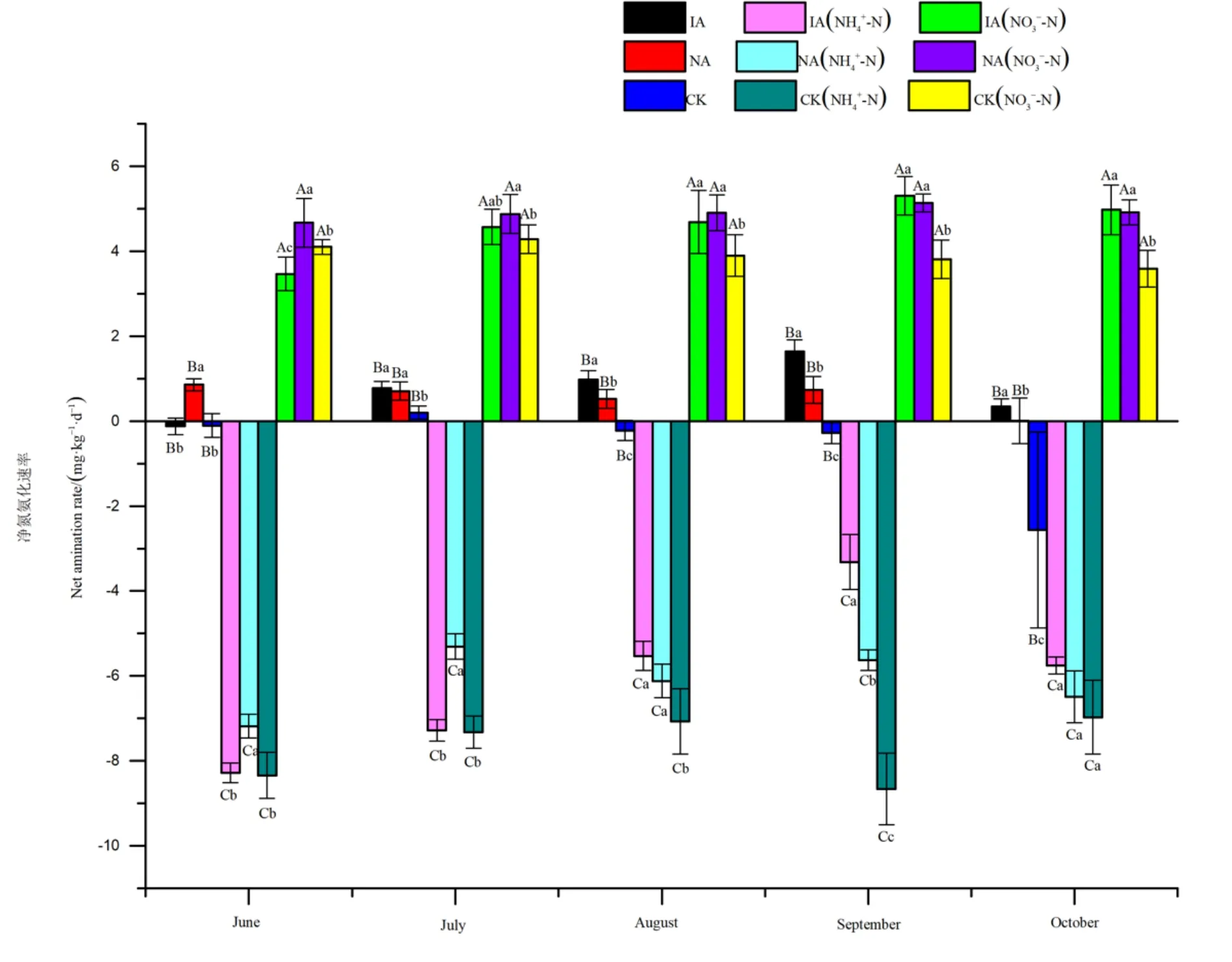
图2 不同植被类型与氮素处理见间的净氮氨化速率比较Fig.2 Net amination rate in different vegetation types and nitrogen treatments at different periods
添加铵态氮显著降低了这 3组处理的净氨化速率,降低了9—12倍。均值大小顺序为入侵地 (-6.03 mg·kg-1·d-1)>鹅观草样地 (-6.14 mg·kg-1·d-1)>裸地(-7.67 mg·kg-1·d-1)。在 6—10 月,土样净氨化速率范围为入侵地-8.28— -3.31 mg·kg-1·d-1,鹅观草样地-7.18 — -5.31 mg·kg-1·d-1, 裸 地-8.66 — -6.97 mg·kg-1·d-1。与鹅观草样地相比,入侵地净氨化速率在6—7月显著降低,8月显著增加。与裸地相比,入侵地净氨化速率在8—9月显著增加)。
添加硝态氮显著增加了这3组处理的净氨化速率,增加了 4—5倍。均值大小顺序为鹅观草样地(4.90 mg·kg-1·d-1)>入侵地 (4.60 mg·kg-1·d-1)>裸地(3.94 mg·kg-1·d-1)。在 6—10 月,土样净氨化速率范围为入侵地 3.47—5.31 mg·kg-1·d-1,鹅观草样地4.67—5.14 mg·kg-1·d-1,裸地 3.59—4.29 mg·kg-1·d-1。与鹅观草样地相比,入侵地净氨化速率在6月显著降低。与裸地相比,入侵地净氨化速率在6月显著降低),在8—10月显著增加。
由表2可以看出,少花蒺藜草入侵与外源氮添加均对土壤净氮氨化速率有显著影响,且这两个因素的交互效应显著。其中,与净氨化速率的关联性(R2)的大小次序为,添加外源氮处理>外来植物入侵>交互效应。
2.3 少花蒺藜草入侵与外源氮对土壤净氮矿化速率的影响
由图3可得,在不添加氮素土样中,净矿化速率差异显著,均值大小顺序为入侵地 (0.43 mg·kg-1·d-1)>鹅观草样地 (0.13 mg·kg-1·d-1)>裸地(-0.31 mg·kg-1·d-1)。在 6—10 月,土样净矿化速率范围为入侵地-0.44—1.14 mg·kg-1·d-1,鹅观草样地-0.43 — 0.49 mg·kg-1·d-1, 裸 地 -1.72 — 0.12 mg·kg-1·d-1。与鹅观草样地相比,入侵地净矿化速率在6月显著降低,在7—9月显著增加。与裸地相比,入侵地净矿化速率在6—10月均显著增加。

表2 净氮氨化速率的主体效应检验Table 2 Inter-subjective test of net amination rate
添加铵态氮显著增加了这3组处理的净矿化速率,增加了5—22倍。均值大小顺序为裸地 (3.38 mg·kg-1·d-1)>鹅观草样地 (2.95 mg·kg-1·d-1)>入侵地 (2.63 mg·kg-1·d-1)。在 6—10 月,土样净矿化速率范围为入侵地 0.74—3.94 mg·kg-1·d-1,鹅观草样地 1.56—3.94 mg·kg-1·d-1,裸地 1.88—5.27 mg·kg-1·d-1。与鹅观草样地相比,入侵地净矿化速率在6月显著增加,在8—10月显著降低。与裸地相比,入侵地净矿化速率在10月显著降低。
添加硝态氮也显著增加了这3组处理的氮矿化速率,增加了0.4—6倍。均值大小顺序为鹅观草样地(0.97 mg·kg-1·d-1)>裸地 (0.93 mg·kg-1·d-1)>入侵地 (0.62 mg·kg-1·d-1)。在 6—10 月,土样净氮矿化速率范围为入侵地 0.08—0.94 mg·kg-1·d-1,鹅观草样地 0.62—1.43 mg·kg-1·d-1,裸地 0.42—1.13 mg·kg-1·d-1。与鹅观草样地相比,入侵地净矿化速率在6月和8—10月显著降低。与裸地相比,入侵地净矿化速率在6—8月和10月显著降低,在9月显著增加。
由表3可以看出,少花蒺藜草入侵与外源氮添加均对土壤净氮矿化速率有显著影响,且这两个因素的交互效应显著。其中,与净矿化速率的关联性(R2)的大小次序为,添加外源氮处理>交互效应>外来植物入侵。
3 讨论

图3 不同植被类型与氮素处理见间的净氮矿化速率比较Fig.3 Net mineralization rate in different vegetation types and nitrogen treatments at different periods

表3 净氮矿化速率的主体效应检验Table 3 Inter-subjective test of net mineralization rate
入侵生态学机制的重要内容之一就是入侵植物对氮循环的影响。大量研究表明,外来入侵植物影响入侵地生态系统氮循环的方式之一就是改变土壤氮素环境(Castro-Díez et al.,2014)。本研究选取裸地及本土植物鹅观草样地作为对照,研究少花蒺藜草入侵与外源氮对土壤氮转化速率的影响,以期从土壤氮转化速率的角度探讨少花蒺藜草的入侵机制。
植物与土壤间的相互关系是影响植物扩张强度的重要因素(刘小文等,2016),有研究表明,外来入侵植物通过改变土壤氮素环境、物理化学性质直接或间接改变土壤氮矿化速率、硝化速率和氨化速率(Piper et al.,2015;Zhang et al.,2016),如互花米草入侵中国盐沼湿地显著增加了土壤氮矿化速率(Zhang et al.,2016)等。类似地,在本研究中少花蒺藜草入侵增加了沙质草地土壤氮素矿化速率,并在这一生长季中均差异显著。分蘖期前后,入侵地铵态氮含量分别呈现出较少和较多两种表现,这与分蘖期前后入侵地硝化速率分别呈现出较高和较低两种表现恰好对应。同时,分蘖期前后入侵地氨化速率分别呈现出较低和较高两种表现,这可能是由于植物的氮素利用形式不用导致的,即旱地植物在幼苗期偏好铵态氮,而在主要生育期则主要吸收硝态氮(Christensen,1985),但少花蒺藜草始终偏好于吸收铵态氮。净硝化速率出现负值,可能是由于原料NH4+-N或产物NO3--N被植物或微生物吸收利用;氨化速率出现负值,可能是由于氨化作用产生的NH4+-N被植物或微生物吸收利用,或被硝化细菌硝化为 NO3--N(李铭等,2012)。土壤养分状况关系到植物生长和微生物活动等,土壤氮素转化率取决于土壤中生物活性的高低,最终归因于土壤中有机质和全氮含量等的多少(Sánchez et al.,1997)。后续试验可以通过对土壤养分状态的研究,结合氮转化效率和速率,解释少花蒺藜草对入侵地氮转化的机制。
添加外源氮对土壤氮转化速率的影响比较复杂。于占源等(2007)发现施用尿素显著增加了科尔沁沙质草地土壤净氮矿化速率与硝化速率。外源氮输入增强了土壤氮转化作用,且促进了中国东北部地区沼泽湿地土壤N2O排放(Zhang et al.,2007)。但Entry(2000)曾报道添加铵态氮或硝态氮会显著降低森林湿地土壤中有机质降解和氮矿化速率。本研究发现,少花蒺藜草入侵对氮转化速率的影响表现为:添加NH4+-N或NO3--N对整体的氮转化速率水平有影响,但不改变三样地间硝化和氨化速率的大小关系。与净硝化速率及净氨化速率的关联性大小顺序为:添加外源氮处理>少花蒺藜草入侵>交互效应。与净矿化速率的关联性大小顺序为:添加外源氮处理>交互效应>少华蒺藜草入侵。添加外源氮更有利于裸地及本地植物鹅观草样地的氮矿化作用。Laungani et al.(2009)指出,外来植物在氮限制环境中成功入侵的生态策略,是通过提高土壤氮库周转或降低植物氮库周转,进而实现对生态系统氮库周转产生影响。本实验仅证明了少花蒺藜草加速了入侵地氮库的周转,但其自身氮库如何变化尚待深入研究。科尔沁沙质草地土壤氮的有效性较低,施氮肥可明显提高土壤供氮能力(于占源等,2007)。与少花蒺藜草入侵地相比,添加外源氮更有利于裸地及本地植物鹅观草样地的氮矿化作用。这表明,少花蒺藜草在氮素含量较低的环境中更能表现出入侵者的优越性,但其究竟是通过何种途径来影响入侵地的氮素变化,有待进一步研究。
4 结论
本实验通过对少花蒺藜草入侵地、鹅观草样地和裸地添加外源氮素来研究3种土壤氮转化率的变化,结果发现:
(1)少花蒺藜草入侵地相对于鹅观草样地和裸地土壤氮净矿化速率更高,以少花蒺藜草分蘖期为节点,土壤氮硝化速率和氮氨化速率呈相反趋势变化:分蘖期前,土壤氮硝化速率增加,分蘖期后,土壤氮氨化速率增加。
(2)添加NH4+-N或NO3--N显著影响了3个样地间土壤氮硝化速率和氮氨化速率,但并未改变三者的大小关系;添加外源氮素后鹅观草样地和裸地的氮矿化水平比少花蒺藜草入侵地有了显著的提高。
综上表明少花蒺藜草在氮素含量低的沙质环境中有较好的适应性,这在一定程度上为少花蒺藜草的入侵生态学机制奠定了理论基础。
猜你喜欢
舰船科学技术(2022年11期)2022-07-15
亚太传统医药(2021年9期)2021-10-26
农业资源与环境学报(2021年5期)2021-10-06
文艺生活·中旬刊(2020年8期)2020-11-23
河南畜牧兽医(2020年11期)2020-01-11
读者·校园版(2018年11期)2018-05-15
现代园艺(2018年3期)2018-02-10
上海农业学报(2017年3期)2017-04-10
农村农业农民·B版(2015年9期)2015-10-16
养殖与饲料(2015年3期)2015-01-03
