
大蒜有机硫化物对解淀粉芽孢杆菌的抑菌作用及机理
2020-07-16 15:13代博仁申光辉刘海娜张丽丹何春桥葛润邱
华南农业大学学报 2020年4期
代博仁,申光辉,刘海娜,张丽丹,何春桥,葛润邱
(四川农业大学 食品学院,四川 雅安 625014)
豆杆,又称豆筋、豆棒,是川渝地区的一种非发酵腐竹类豆制品[1]。预浸泡豆杆食用方便,营养丰富,口感独特,深受市场欢迎。但预浸泡豆杆水分活度高,易腐败变质。郑丽君等[2]研究表明,解淀粉芽孢杆菌Bacillus amyloliquefaciens是真空包装预浸泡豆杆的优势腐败菌,具有很强的耐高温能力和较强的胞外蛋白酶产生能力,在腐败过程中,通过降解蛋白质产生大量挥发性盐基氮和生物胺等有害物质,严重影响产品质量安全[3]。夏硕等[4]采用传统热杀菌方法对预浸泡豆杆进行处理,结果表明110 ℃条件下能够有效杀灭芽孢细菌,但对产品硬度、弹性等质构品质造成了较为严重的破坏。唐桂宇等[5]复配了Nisin、ε–聚赖氨酸和山梨酸钾,结合100 ℃、20 min热杀菌处理,豆杆货架期仅延长了4 d,与实际应用尚存在一定差距。因此,寻求既安全有效,又能保持良好质构特性的预浸泡豆杆腐败芽孢杆菌的控制措施,对于保证产品品质与安全具有重要意义。
植物源天然防腐剂具有种类丰富、广谱抑菌等特点,是筛选安全有效的防腐剂的重要来源之一,受到国内外研究者的广泛关注[6-7]。大蒜Allium sativum是百合科Liliaceae葱属Allium植物的地下鳞茎,富含有机硫化物、黄酮类化合物、皂苷类化合物和多糖等活性成分[8],具有良好的抑菌、杀菌活性[9]。目前已证明,大蒜中的蒜氨酸在受到蒜氨酸酶作用后会生成二烯丙基二硫醚、阿霍烯等具有抑菌作用的硫化物[8-10]。其中二烯丙基二硫醚与阿霍烯能够干扰细菌正常生理代谢[11-12],具有广谱抑菌的特点,能够有效抑制德式乳杆菌Lactobacillus delbruckii[13]、阪崎克罗诺杆菌Cronobacterium sakazakii[12]和蜡样芽孢杆菌Bacillus cereus[14]等细菌。而目前关于大蒜活性成分对解淀粉芽孢杆菌的抑菌作用及其机理研究鲜见报道。彭光华等[15]采用乙醇萃取大蒜有机硫化物(Garlic organic sulfide,GOS),通过气相色谱–质谱法分析发现大蒜有机硫化物中含量最高的组分为大蒜素热分解衍生得到的噻烯类化合物(相对含量为55.8%)及丙烯基硫醚类化合物(相对含量为6.04%)。在前期对多种香辛料抑菌活性筛选试验中发现,用φ为75%乙醇溶液提取获得的大蒜有机硫化物对2株致腐解淀粉芽孢杆菌具有较强的抑菌活性。
因此,本文主要通过测定大蒜有机硫化物对2株优势腐败菌的抑制作用、杀灭性能及对胞外蛋白酶抑制活性,并通过考察大蒜有机硫化物处理对细菌的代谢活性、细胞壁和细胞膜通透性的影响,扫描电镜观察处理后的细胞形态变化,明确其对解淀粉芽孢杆菌的抑菌作用机理,为利用大蒜有机硫化物控制预浸泡豆杆腐败芽孢杆菌提供依据。
1 材料与方法
1.1 材料
1.1.1 菌株 解淀粉芽孢杆菌菌株 DY1a、DY1b,分离自腐败豆杆样品,保存于四川农业大学食品学院。
1.1.2 材料与试剂 新鲜瓣蒜 (产地:四川成都)购于四川雅安吉选超市。
邻硝基苯–β–D–半乳吡喃糖苷(ONPG)购于碧云天生物技术有限公司;2, 3, 5–三苯基氯化四氮唑(TTC)购于北京索莱宝科技有限公司;碱性磷酸酶试剂盒购于南京建成生物工程研究所;其余试剂(分析纯)购于成都市科龙化工试剂厂。
1.1.3 培养基和试剂的配制 LB 液体培养基:胰蛋白胨 10.0 g/L,酵母浸出粉 5.0 g/L,NaCl 10.0 g/L,pH自然。
营养琼脂培养基:蛋白胨 10.0 g/L,牛肉膏 3.0 g/L,NaCl 5.0 g/L,琼脂 20.0 g/L,pH 自然。
PCA 培养基:胰蛋白胨 5.0 g/L,酵母浸粉 2.5 g/L,葡萄糖 1.0 g/L,琼脂 15.0 g/L,pH 自然。
牛奶琼脂培养基:A液,脱脂奶粉50.0 g/L,溶解于1/2最终体积的培养基中;B液,琼脂20.0 g/L,溶解于1/2最终体积的培养基中。灭菌后将A液与B液混合。
M9 乳糖诱导培养基:Na2HPO412.8 g/L,KH2PO43.0 g/L,NaCl 0.5 g/L,NH4Cl 1.0 g/L,MgSO40.5 g/L,CaCl20.01 g/L,乳糖 5.0 g/L,pH 自然。
灭菌条件:121 ℃ 15 min(牛奶琼脂培养基 A 液的灭菌条件为 115 ℃ 20 min)。
β–半乳糖苷酶反应缓冲液:NaCl 8.0 g/L,KCl 0.2 g/L,Na2HPO4·12H2O 2.9 g/L,KH2PO40.24 g/L,MgSO4·7H2O 0.25 g/L,β–巯基乙醇 3.9 g/L。
1.2 主要仪器
CPA225D型电子天平,德国赛多利斯公司;YXQ-LS-50SⅡ立式压力蒸汽灭菌器,上海博讯实业有限公司;SW-CJ-2FD洁净工作台,苏净集团苏州安泰空气技术有限公司;ZWY-211B恒温培养振荡器,上海智城分析仪器制造有限公司;KH-300DE型数控超声波清洗器,昆山禾创超声仪器有限公司;RE-2000B型旋转蒸发器,上海亚荣生化仪器厂;UV-3100PC紫外分光光度计,上海美谱达仪器有限公司;Thermo BR4i型冷冻离心机,美国赛默飞世尔科技;Varioskan Flash型荧光酶标仪,美国赛默飞世尔科技;ZEISS Sigma 500 场发射扫描电镜,德国卡尔蔡司公司。
1.3 试验方法
1.3.1 大蒜有机硫化物的制备 参照彭光华等[15]的方法提取大蒜有机硫化物,稍作修改。取45 g新鲜瓣蒜,剥皮去蒂,加入φ为75%的乙醇溶液180 mL,打浆机破壁,300 W 超声辅助提取 90 min,提取液 4 ℃条件下 4 000 r/min 离心 10 min,收集上清液,50 ℃条件下减压浓缩脱除乙醇,蒸馏水定容至15 mL,获得3 g/mL的大蒜有机硫化物,冷冻保存备用。
1.3.2 大蒜有机硫化物抑菌活性测定 将解淀粉芽孢杆菌涂布于营养琼脂培养基中,均匀放置3个牛津杯 (直径 7 mm)。用 0.22 μm 水系滤膜过滤大蒜有机硫化物,再用无菌蒸馏水进行10倍梯度稀释。分别取150 μL大蒜有机硫化物原液及10倍、100倍稀释液于牛津杯中,将平板正置,37 ℃条件下培养24 h,测定抑菌圈直径。
1.3.3 大蒜有机硫化物对解淀粉芽孢杆菌的最低抑菌浓度 (Minimum inhibitory concentration,MIC)与最低杀菌浓度 (Minimum bactericidal concentration,MBC) 采用试管二倍稀释法制备添加不同浓度大蒜有机硫化物的LB培养基1.5 mL,接种菌液至终浓度 105CFU/mL,吸取 200 μL 菌液于 96 孔板中,测定D600nm,记为初始值;37 ℃ 条件下 160 r/min振荡培养 24 h,吸取各管菌液 200 μL于96孔板中,测定样品D600nm,记为终止值。不同浓度样品重复3次,比较各样品终止值与初始值的差值 ΔD600nm,ΔD600nm<0.01 的最低浓度即为MIC。从MIC及以上浓度的试管中取10 μL培养液涂布于PCA平板中,37 ℃条件下培养24 h,无细菌生长的最低浓度即为MBC。
1.3.4 细菌生长曲线测定 向LB液体培养基中接种培养24 h的细菌菌悬液,调节菌液浓度至105CFU/mL并分装,加入大蒜有机硫化物,使其质量浓度为 15 和 30 mg/mL,37 ℃ 条件下 160 r/min振荡培养,定时取样,测定D600nm,并绘制生长曲线。以未添加大蒜有机硫化物的LB液体培养基为空白对照。
1.3.5 细菌胞外蛋白酶活性测定 向LB培养基中分别加入15、30和45 mg/mL大蒜有机硫化物,接种菌悬液,调节浓度至 106CFU/mL,37 ℃ 条件下160 r/min振荡培养24 h,以未添加大蒜有机硫化物为对照。各培养液 10 000 r/min 离心 10 min,上清液使用0.22 μm滤膜过滤,获得无菌胞外溶液。在牛奶琼脂平板上放置4个牛津杯,分别向牛津杯内加入150 μL的无菌胞外溶液,置于37 ℃恒温培养箱,24 h后观察并测定水解圈直径,并计算大蒜有机硫化物对胞外蛋白酶活性的抑制率:
蛋白酶活性抑制率=(对照的水解圈面积−处理的水解圈面积)/对照的水解圈面积×100%。
1.3.6 细胞代谢活性测定 将培养 24 h 的腐败菌菌悬液在 4 ℃ 条件下 4 000 r/min 离心 10 min,用无菌生理盐水洗涤菌体沉淀3次,加入生理盐水重悬菌体,调节细胞浓度至106CFU/mL并分装,再按浓度分别加入0、30和120 mg/mL大蒜有机硫化物。取上述添加不同浓度大蒜有机硫化物的腐败菌菌悬液 1 mL,加入 0.05 mol/L 的 Tris-HCl缓冲液(pH=8.6)2 mL、2 mol/L 的葡萄糖溶液 2 mL、1 mg/mL 的 TTC 溶液 2 mL,37 ℃ 条件下恒温静置孵育5 h后,加入2滴浓硫酸终止反应,再加入5 mL甲苯震荡萃取,以甲苯为空白对照参比测定有机相的D490 nm。
1.3.7 胞外碱性磷酸酶活性测定 定时取 “1.3.6”中添加不同浓度大蒜有机硫化物的腐败菌菌悬液,10 000 r/min 离心 10 min,取上清液,使用碱性磷酸酶 (Alkaline phosphatase,AKP)试剂盒测定碱性磷酸酶活性,AKP定义:以1 mL培养液在37 ℃条件下与基质作用15 min产生1 mg酚为1个酶活性单位(U),测定并计算胞外溶液的碱性磷酸酶活性(U/mL)随时间的变化情况。
1.3.8 胞外β–半乳糖苷酶活性测定 向 “1.3.6”中的菌体沉淀中加入M9乳糖诱导培养基重悬菌体,调节菌浓度至106CFU/mL,分装,分别加入0、15和30 mg/mL大蒜有机硫化物,置于37 ℃条件下 160 r/min 振荡孵育 5 h。定时取样,10 000 r/min离心 10 min,取上清液 50 μL 于 96 孔板中,加入β–半乳糖苷酶反应缓冲液100 μL,邻硝基苯β–D–半乳吡喃糖苷 (ONPG, 10 mg/mL)50 μL,摇匀后置于 37 ℃ 条件下孵育 2 h,加入 1 mol/L 的 Na2CO3溶液50 μL终止反应,用酶标仪测定D420nm随时间的变化情况。
1.3.9 胞外溶液电导率测定 参照陶文卿[16]的方法测定胞外溶液电导率。用0.278 mol/L的葡萄糖溶液洗涤菌体沉淀,直至菌悬液电导率与等渗溶液电导率相等。调节菌体浓度至106CFU/mL。随后,加入30 mg/mL大蒜有机硫化物,置于37 ℃条件下160 r/min振荡孵育24 h。定时取样,测定菌悬液的电导率随时间的变化情况。以不加提取物的菌悬液为对照。
1.3.10 细胞形态结构扫描电镜观察 向108CFU/mL的菌悬液中加入大蒜有机硫化物,使其质量浓度分别为30和480 mg/mL,以不加提取物的菌悬液为对照。37 ℃ 条件下振荡孵育 12 h后,4 ℃条件下 4 000 r/min 离心 10 min,弃去上清液得到菌体沉淀。参照Guo等[17]的方法进行扫描电镜样品制备,向菌体沉淀加入φ为2.5%的戊二醛溶液10 mL,4 ℃条件下固定12 h,先后使用φ为30%、50%、70%和90%的乙醇溶液脱水15 min,最后使用无水乙醇脱水2次,每次15 min,挑取适量脱水菌体于无菌盖玻片上,冷冻干燥后喷金,场发射扫描电子显微镜观察、拍照。
1.3.11 数据统计分析 采用 SPSS 22.0 软件对试验结果进行差异显著性分析,Duncan’s法进行多重比较;采用OriginPro 9.0软件作图。
2 结果与分析
2.1 大蒜有机硫化物的抑菌效果
由图1可知,3 g/mL的大蒜有机硫化物产生的抑菌圈直径最大,对菌株DY1a的抑菌圈直径为(31.7±3.2) mm,对菌株 DY1b的抑菌圈直径为(25.7±1.5) mm;300 mg/mL 的大蒜有机硫化物产生的抑菌圈直径次之,对DY1a的抑菌圈直径为(21.0±4.6) mm,对 DY1b 抑菌圈直径为 (15.0±1.5)mm;30 mg/mL的大蒜有机硫化物没有产生抑菌圈。由图2可知,大蒜有机硫化物对2株腐败细菌的最小抑菌浓度(MIC)均为30 mg/mL(即稀释倍数为1/32)。低于MIC的大蒜有机硫化物对2株腐败菌具有一定的生长促进作用。由表1可知,大蒜有机硫化物对于2株细菌的最小杀菌浓度(MBC)不同,对菌株DY1a的MBC为240 mg/mL,对菌株DY1b 的 MBC为 480 mg/mL。
2.2 解淀粉芽孢杆菌的生长曲线
由图3A可知,空白组的菌株DY1a在0~4 h内处于延滞期,在4 h后进入对数生长期,在12 h后进入稳定期。15 mg/mL大蒜有机硫化物处理组的菌株 DY1a在 0~28 h 内D600nm没有变化,然而在 28 h后,D600nm急速升高。30 mg/mL 大蒜有机硫化物处理组的菌株DY1a在48 h内D600nm均无变化。由图3B可知,空白组的菌株DY1b在0~8 h内处于延滞期,在8 h后进入对数生长期,在33 h后进入稳定期。15 mg/mL大蒜有机硫化物处理组的菌株DY1b在0~22 h 内D600nm没有变化,之后D600nm迅速升高。30 mg/mL大蒜有机硫化物处理组的菌株DY1b 在 48 h 内D600nm无变化。

图2 不同浓度大蒜有机硫化物(GOS)对2株解淀粉芽孢杆菌生长的影响Fig. 2 Effects of different concentrations of garlic organic sulfide (GOS) on the growth of two strains of Bacillus amyloliquefaciens
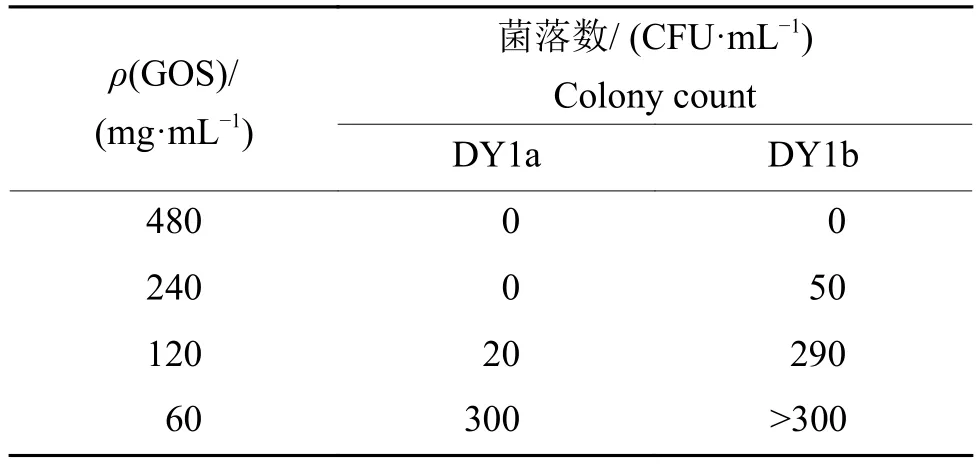
表1 不同浓度大蒜有机硫化物(GOS)对2株解淀粉芽孢杆菌的杀菌作用Table 1 Bactericidal effect of different concentrations of garlic organic sulfide (GOS) against two strains of Bacillus amyloliquefaciens
2.3 解淀粉芽孢杆菌胞外蛋白酶活性

图3 不同浓度大蒜有机硫化物(GOS)对2株解淀粉芽孢杆菌生长曲线的影响Fig. 3 Effects of different concentrations of garlic organic sulfide (GOS) on the growth curves of two strains of Bacillus amyloliquefaciens
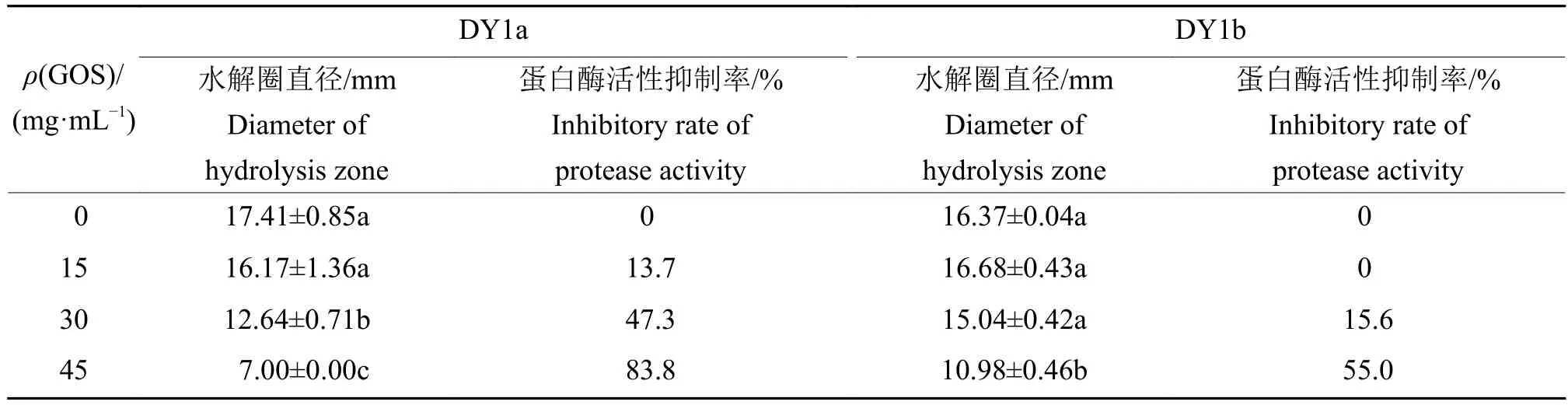
表2 不同浓度大蒜有机硫化物(GOS)对2株解淀粉芽孢杆菌胞外蛋白酶活性的影响1)Table 2 Effects of different concentrations of garlic organic sulfide (GOS) on extracellular protease activities of two strains of Bacillus amyloliquefaciens
腐败菌胞外蛋白酶是高蛋白食品重要的致腐因子。由表2可知,不同浓度大蒜有机硫化物对解淀粉芽孢杆菌DY1a与DY1b胞外蛋白酶活性具有不同程度的抑制作用,随着大蒜有机硫化物添加浓度的提高,蛋白水解圈直径逐渐减小。当大蒜有机硫化物添加质量浓度为15 mg/mL时,对2株腐败菌胞外蛋白酶的活性无显著影响(P>0.05);添加30和45 mg/mL大蒜有机硫化物对解淀粉芽孢杆菌DY1a的蛋白酶活性具有显著抑制效果,抑制率分别为47.3%和83.8%。添加30和45 mg/mL大蒜有机硫化物对菌株DY1b胞外蛋白酶活性也具有抑制作用,抑制率分别为15.6%和55.0%,且后者差异显著。
2.4 解淀粉芽孢杆菌细胞代谢活性
2,3,5−三苯基氯化四氮唑(TTC)能够与活细胞线粒体内的呼吸链脱氢酶发生反应,生成红色的甲臜,可用来反映细胞的代谢活性[18]。通过对大蒜有机硫化物处理后解淀粉芽孢杆菌培养液TTC染色,测定细菌代谢活性,可间接反映菌株细胞的活性情况。由图4可知,空白对照组的细胞代谢活性显著高于大蒜有机硫化物组(P<0.05),大蒜有机硫化物质量浓度为30 mg/mL时,菌株DY1a与DY1b的代谢活性显著降低,D490nm与对照组相比分别下降了2.771和4.091,且随着大蒜有机硫化物浓度的进一步升高而继续减小。此外,在相同大蒜有机硫化物浓度下,菌株DY1b的细胞代谢活性高于菌株DY1a。
2.5 解淀粉芽孢杆菌细胞壁的完整性
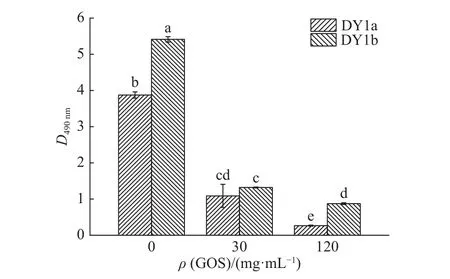
图4 不同浓度大蒜有机硫化物(GOS)对2株解淀粉芽孢杆菌细胞代谢活性的影响Fig. 4 Effects of different concentrations of garlic organic sulfide (GOS) on metabolic activities of two strains of Bacillus amyloliquefaciens
碱性磷酸酶(AKP)是存在于细菌细胞壁和细胞膜之间的酶,在正常情况下,AKP不会泄露到胞外,但当细胞壁结构受到破坏后,会泄露至胞外溶液中[19]。因此通过检测胞外溶液中AKP活性的变化,可以间接反映大蒜有机硫化物对解淀粉芽孢杆菌细胞壁完整性的影响。由图5可知,解淀粉芽孢杆菌DY1a与DY1b的空白对照的胞外溶液AKP活性在0~5.0 h内无明显变化,而添加15和30 mg/mL大蒜有机硫化物的菌株胞外溶液AKP活性在1.0 h内迅速升高,且大蒜有机硫化物添加浓度越高,胞外溶液AKP活性越高;添加30 mg/mL大蒜有机硫化物组的胞外溶液在5.0 h时,菌株DY1a和DY1b的胞外AKP活性分别提高了0.029和 0.036 U/mL,均显著高于 15 mg/mL组与空白对照。由此可以发现,大蒜有机硫化物对腐败解淀粉芽孢杆菌细胞壁的完整性具有较强的破坏作用,且浓度越高,破坏作用越强。
2.6 解淀粉芽孢杆菌细胞膜的通透性与完整性
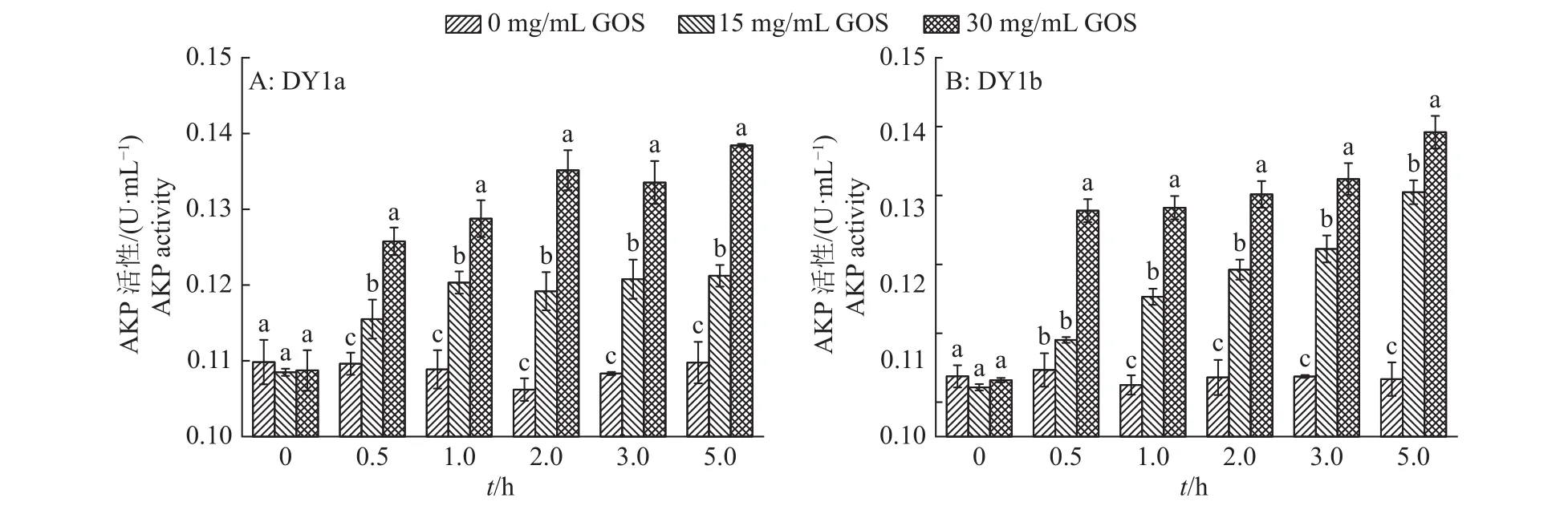
图5 不同浓度大蒜有机硫化物(GOS)处理2株解淀粉芽孢杆菌的胞外碱性磷酸酶(AKP)活性的变化Fig. 5 Changes of extracellular AKP activity in two strains of Bacillus amyloliquefaciens treated with different concentrations of garlic organic sulfide (GOS)
正常生长条件下,细菌细胞膜内的物质不会泄露到胞外,但当细胞膜通透性和完整性遭受破坏后,细胞膜内的电解质和胞内酶将出现不同程度的泄露,导致胞外溶液电导率的增加。β–半乳糖苷酶位于细菌细胞膜内,正常生理状态下在胞外检测不到其活性。因此可以通过胞外溶液电导率及β–半乳糖苷酶活性变化反映细菌细胞膜的通透性和完整性[20]。由图6可知,未添加大蒜有机硫化物的腐败解淀粉芽孢杆菌培养液电导率变化较为平缓,而添加大蒜有机硫化物的腐败菌培养液电导率随着培养时间的增加而逐渐提高,浓度为30 mg/mL的GOS处理细菌8 h后,菌株DY1a和DY1b的培养液电导率分别升了0.060和0.031 mS/cm,显著高于对照。由图7可知,在0 h时,培养液中β–半乳糖苷酶活性大小几乎相同,添加大蒜有机硫化物30 min后,2株解淀粉芽孢杆菌培养液中的酶活性快速增加,30 mg/mL大蒜有机硫化物处理DY1a与DY1b菌株 5.0 h 后,胞外β–半乳糖苷酶活性提高,D420nm分别提高了0.047与0.016,显著高于对照;且培养液β–半乳糖苷酶活性高低与大蒜有机硫化物添加浓度呈正相关。此外,解淀粉芽孢杆菌DY1b的培养液电导率和β–半乳糖苷酶活性的上升幅度小于菌株DY1a,这也与前述胞外蛋白酶和代谢活性的试验结果一致,表明菌株DY1a的菌体代谢和细胞结构对大蒜有机硫化物更敏感。

图6 大蒜有机硫化物(GOS)处理2株解淀粉芽孢杆菌的胞外电导率变化Fig. 6 Changes of extracellular electrical conductivity in two strains of Bacillus amyloliquefaciens treated with garlic organic sulfide (GOS)

图7 不同浓度大蒜有机硫化物(GOS)处理2株解淀粉芽孢杆菌的胞外β–半乳糖苷酶活性变化Fig. 7 Changes of extracellular β-galactosidase activity in two strains of Bacillus amyloliquefaciens treated with different concentrations of garlic organic sulfide (GOS)
2.7 解淀粉芽孢杆菌的形态结构变化
从图8可以看出,未经大蒜有机硫化物处理的解淀粉芽孢杆菌菌体细胞没有异常变化,形态饱满,表面光滑,细胞与细胞之间分界清晰(图8A、8D),质量浓度为30 mg/mL的大蒜有机硫化物处理5.0 h后的菌体细胞变形,表面出现皱缩,孔洞式塌陷(图 8B、8E),质量浓度为 480 mg/mL 的大蒜有机硫化物处理后,菌体细胞与细胞之间出现黏聚、破裂现象(图8C、8F)。细胞表面皱缩和孔洞现象进一步证明了大蒜有机硫化物处理可破坏细胞壁、细胞膜的通透性和完整性,导致胞内物质泄漏至胞外,甚至最终使细胞死亡。

图8 不同浓度大蒜有机硫化物(GOS)处理2株解淀粉芽孢杆菌细胞表面形态Fig. 8 Cell surface morphology of two strains of Bacillus amyloliquefaciens treated by different concentrations of garlic organic sulfide (GOS)
3 讨论与结论
解淀粉芽孢杆菌是预浸泡豆杆制品的优势腐败菌,具有耐高温、产蛋白酶能力强的特点。大蒜作为重要的香辛料,具有抑菌谱广、杀菌能力强的优势,在食品绿色抑菌保鲜领域具有重要的应用潜力。已有研究发现,大蒜提取物对蜡样芽孢杆菌Bacillus cereus[14]、枯草芽孢杆菌Bacillus subtilis[21-22]等芽孢细菌具有良好的抑制作用。目前国内外关于大蒜提取物对豆制品腐败菌−解淀粉芽孢杆菌的抑菌研究鲜见报道。本研究结果表明,大蒜有机硫化物对导致豆杆腐败的解淀粉芽孢杆菌菌株DY1a、DY1b具有良好的抑菌杀菌作用,最小抑菌浓度均为30 mg/mL,最小杀菌浓度分别为240和480 mg/mL。同时,大蒜有机硫化物对解淀粉芽孢杆菌胞外蛋白酶这一重要腐败因子具有很强的抑菌活性。这些结果显示出大蒜有机硫化物在预浸泡豆杆腐败菌杀菌控制方面具有很大的应用潜力和价值。
本试验对最小抑菌浓度下2株解淀粉芽孢杆菌的代谢活性与胞外蛋白酶活性变化进行了研究,结果发现,在最小抑菌浓度下,2株解淀粉芽孢杆菌仍然保持一定的活性,可见,低浓度下大蒜有机硫化物对解淀粉芽孢杆菌的作用方式并非直接杀死细菌,而是抑制细菌的生理活动,这与宋卫国[23]的研究结论一致。大蒜有机硫化物处理后,细菌胞外碱性磷酸酶、β–半乳糖苷酶活性及胞外溶液电导率升高,因而可推断细菌细胞壁、细胞膜的通透性和完整性受到不同程度的影响,导致胞内电解质与内容物外泄渗出。进一步通过扫描电镜观察发现,此时细菌细胞表面形态出现皱缩、坍塌和破裂现象,这进一步印证了大蒜有机硫化物的抑菌机理与破坏细菌细胞壁和细胞膜的完整性有关,这与胡蕾[24]和Feng等[25]的研究结论一致。
生长曲线试验结果显示15 mg/mL大蒜有机硫化物处理的菌株DY1b的对数生长期早于同处理下的菌株DY1a;胞外蛋白酶试验结果显示,在相同处理下,大蒜有机硫化物对菌株DY1b的蛋白酶抑制率低于DY1a;此外,细菌代谢活性、胞外电导率及β–半乳糖苷酶试验结果表明,解淀粉芽孢杆菌DY1b对大蒜有机硫化物胁迫更具有耐受性。
大蒜有机硫化物对解淀粉芽孢杆菌DY1a和DY1b具有良好的抑菌、杀菌作用,并能抑制胞外蛋白酶活性,其抑菌机理与破坏细菌细胞壁、细胞膜的通透性和完整性,干扰细菌生理代谢活动有关。大蒜有机硫化物在控制预浸泡豆杆优势腐败菌中具有较强的应用潜力。
猜你喜欢
金属热处理(2022年10期)2022-10-25
安徽建筑大学学报(2022年4期)2022-09-27
昆明理工大学学报(自然科学版)(2022年4期)2022-09-07
湖南饲料(2021年4期)2021-10-13
昆明医科大学学报(2021年4期)2021-07-23
当代水产(2021年3期)2021-07-20
食品安全导刊(2021年30期)2021-02-15
生物化工(2020年5期)2020-10-30
农业环境科学学报(2020年9期)2020-10-09
农药科学与管理(2019年6期)2019-11-23
