
白桦MYB基因响应激素及盐旱处理的表达研究
2020-07-14 01:00:18刘佳欣刘慧子石晶静
植物研究 2020年5期
刘佳欣 刘慧子 石晶静 于 颖 王 超
(林木遗传育种国家重点实验室,东北林业大学,哈尔滨 150040)
已知植物茎的伸长生长受多种内源激素的调控,如生长素吲哚乙酸(IAA)、赤霉素(GA)、油菜素内酯(brassinosteroid,BR)和乙烯(ethylene,ETH)等都促进茎的伸长生长[9]。而外源激素的施加能够改变内源激素的含量,并影响茎生长发育。IAA、GA3和IBA浸种的两两交互作用对云南松胚轴和根长具有极显著或显著的差异影响[10]。外源激素GA3与ABA对烤烟茎尖处内源激素GA、ABA、IAA含量以及株高、节间距伸长产生影响[11]。喷施外源激素GA3可调节矮秆基因型烟草茎尖中各内源激素的含量,与正常株高基因型“K326”的各激素含量相接近,从而促进茎生长[12]。从外界信号到性状改变的过程是如何通过一系列的分子响应完成的?哪些基因参与了其调控的分子途径?分子机制如何?是研究激素调控植物生长发育的重要研究内容之一。
MYB转录因子是植物转录因子家族中最大的家族之一[1],不仅在植物生长发育调控中调控生长发育的节奏以及器官的形成,而且可以响应非生物胁迫诱导从而影响代谢产物的形成与激素信号传导等[2~4]。转录因子的主要作用方式时通过与下游启动子上的顺式作用原件相结合的方式来调控靶细胞基因的表达从而形成同源、异源二聚体,也可与其他蛋白互作成为某种活化形式参与JA(jasmonate acid)、SA(salicylic acid)、ABA(abscisic acid)等信号传导途径,形成基因表达的调控网络。遗传学分析发现ABA诱导花青素合成部分依赖于MBW复合体中的核心转录因子,如TTG1、TT8及MYB75等[5]。水曲柳FmMYB5基因对IAA(indole-3-acetic acid)、ABA、GA3(gibberellin)、JA和SA 5种激素诱导都存在显著响应且都是正调控作用[6]。核桃JrEFM1可被ABA、JA、SA处理不同程度地诱导[7]。拟南芥AtMYB51受到水杨酸强烈诱导,AtMYB34则对茉莉酸甲酯最为敏感,处理后表达水平显著提高[8]。这些研究说明植物MYB转录因子广泛参与外源激素调控的植物发育中。种子萌发到下胚轴和胚根的形成是植物发育、形态建成的重要生物学过程,受到不同激素的调控,研究该过程响应激素处理的MYB转录因子表达模式,对研究木本植物发育及形态建成的分子调控具有一定的研究价值。
白桦(Betulaplatyphylla)是我国北方重要的经济和园林绿化树种之一[13],其具有材质洁白细腻,抗旱耐瘠薄的优良特性,适于做纸浆材,单板材和观赏树木。是研究林木重要性状形成分子机制的良好材料,也是重要的改良树种。本研究以白桦为材料,选择MeJA(Methyl jasmonate)、ETH、ABA和KT对野生型白桦种子进行喷洒处理,比较不同处理条件下白桦苗与对照苗胚轴及根发育情况;利用实时荧光定量PCR对经过不同激素喷洒、NaCl和Mannitol处理下的白桦BpMYBs基因的表达量进行分析。分析其响应激素处理的表达模式,为了进一步明确BpMYB家族基因的信号调控及功能提供依据。
1 材料与方法
1.1 实验材料
将野生型白桦种子用流水冲泡2 d,然后种到人工营养土壤(腐殖质∶蛭石∶珍珠岩3∶1∶1)中覆膜保持湿度(3 d后将膜揭掉),放置于生化培养箱内,第二天开始进行激素处理,分别用10 μmol·L-1的MeJA、100 μmol·L-1的ETH、20 μmol·L-1的ABA和50 μmol·L-1的KT对白桦种子进行喷洒处理,同时以喷洒清水处理作为对照,期间每隔1 d喷洒1次。待白桦苗长到4周大小时,将不同激素处理后的白桦苗完整的从土壤中取出,用清水将根部的土壤洗净,平铺在干净的玻璃平板上进行观察,并照相。用于基因表达分析的材料迅速用液氮处理,放于-80℃冰箱内保存备用。取8周大小的白桦苗,分别由200 mmol·L-1NaCl和100 mmol·L-1Mannitol处理6、12、24和48 h,用水处理白桦苗作对照进行基因表达分析。
1.2 RNA提取及cDNA合成
使用CTAB法提取白桦中的总RNA,利用反转录试剂盒(Takara,日本)进行一链cDNA的合成。反应体系为20 μL:2×TS Reaction Mix10 μL,TransScript® RT/RI Enzyme Mix 1 μL,Anchored Oligo(dT)18 Primer(0.5 μg·μL-1) 1 μL,Total RNA 50 ng至5 μg,gDNA Remove 1 μL,补充RNase-Free Water至20 μL。反应条件:在PCR仪(Biometra)中,42℃条件下,反应30 min;85℃条件下,反应5 s。实验结束后,将cDNA于-20℃冰箱保存备用。
1.3 基因克隆及生物信息学分析
对白桦应拉木转录组数据进行分析,筛选鉴定出转录组中的BpMYB基因序列,利用NCBI的ORF finder程序进行编码区分析并鉴定其完整的CDS序列。以野生型白桦cDNA为模板,根据白桦MYB基因CDS序列设计特异引物(见附表1)进行RT-PCR克隆。克隆产物送华大(北京)公司进行测序。利用MEGA(V.6)对白桦MYB基因编码的蛋白与拟南芥已知MYB蛋白序列进行系统进化分析。
1.4 荧光定量RT-PCR分析
根据白桦MYB基因CDS序列,选取200 bp左右长度序列设计特异引物(见表1)进行荧光定量PCR克隆。分别以不同胁迫处理和水对照的白桦苗cDNA为模板进行荧光定量PCR试验,其中以Tubulin(GenBank number:FG067376)和Ubiquitin(GenBank number:FG065618)为内参引物,设置3次重复(包括3次生物学重复以及3次技术重复)。实时荧光定量RT-PCR反应体系20 μL,cDNA 2.0 μL,Primer F(10 μmol·L-1) 1.0 μL,Primer(10 μmol·L-1) 1.0 μL,SYBR Green Realtime PCR Master mix 10.0 μL,补充ddH2O至总体积20 μL。实时荧光定量PCR反应程序:94℃ 30 s;94℃ 12 s,58℃ 15 s,72℃ 40 s,79℃读板1 s,共45个循环。反应结束后,运用ΔΔCt方法对基因的相对表达量进行定量数据分析。

表1 BpMYB家族基因定量引物序列

图1 不同激素处理的白桦幼苗表型分析 a.20 μmol·L-1 ABA;b.50 μmol·L-1 KT;c.100 μmol·L-1 ETH;d.10 μmol·L-1 MeJAFig.1 Phenotype of birch seedlings treated with different hormones a.20 μmol·L-1 ABA;b.50 μmol·L-1 KT;c.100 μmol·L-1 ETH;d.10 μmol·L-1 MeJA
2 结果与分析
2.1 激素处理下白桦表型观察
对白桦种子分别用10 μmol·L-1的MeJA、100 μmol·L-1的ETH、20 μmol·L-1的ABA和50 μmol·L-1的KT进行喷洒处理,4周后观测其生长性状。从苗的表型上来看(见图1),经过ETH喷洒处理的白桦苗变化最为明显,它的下胚轴比未处理的白桦下胚轴短,根长显著减小,约是未处理白桦苗的20%;经过ABA处理的白桦苗,下胚轴比未处理的白桦苗的下胚轴稍长,根长显著增加,比未处理的根长30%;经过KT处理的白桦苗的下胚轴和根均比未处理的白桦苗的下胚轴和根长;经过茉莉酸处理的白桦苗下胚轴和根比对照稍长。
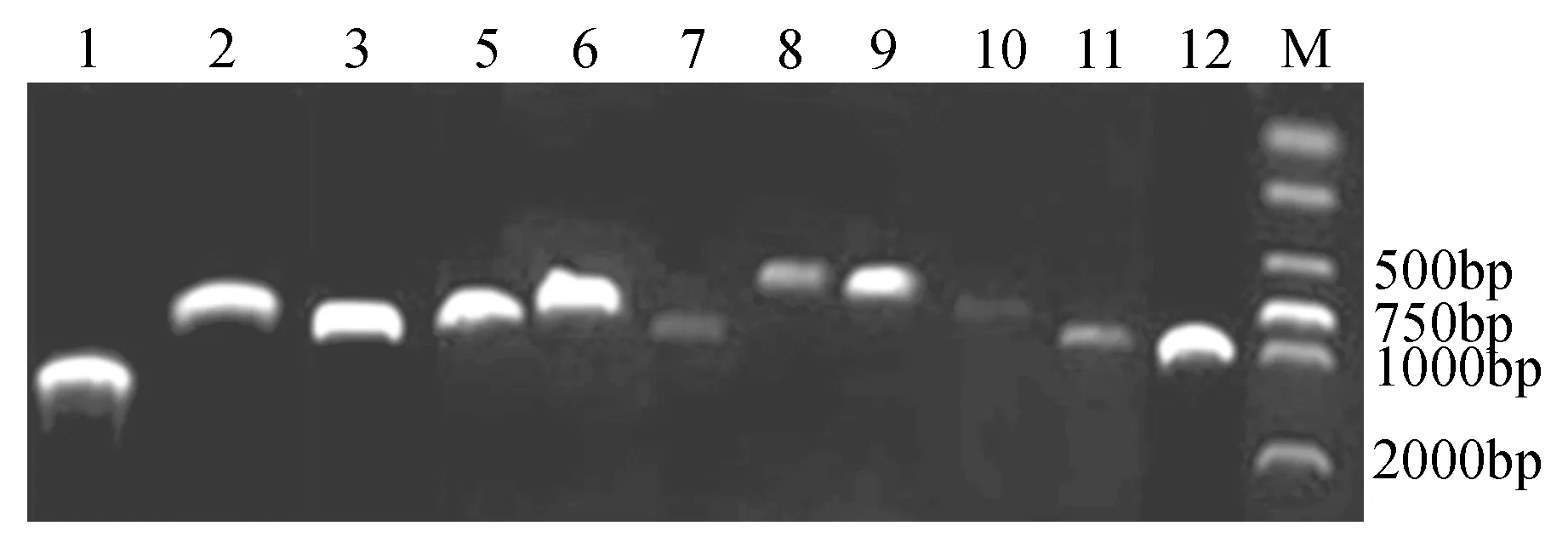
图2 白桦MYB基因RT-PCR克隆Fig.2 RT-PCR cloning of birch MYB gene
2.2 白桦MYBs基因克隆及系统进化分析
通过对所选白桦MYB基因进行RT-PCR,成功扩增出11条目的片段,将这11条MYB目的片段利用胶回收试剂盒(Omega)进行回收,用PMD18-T载体进行连接并且成功转化大肠杆菌,从中随机挑取阳性菌斑,利用11条MYB基因的特异性引物进行菌液PCR,PCR结果显示目的基因条带正确(见图2),将菌液送至公司测序,测序结果与基因序列比对一致,成功克隆得到11条具有完整ORF的白桦MYB序列,这些基因编码区碱基数在624~1 203 bp。根据克隆顺序命名为BpMYB1、2、3、5、6、7、8、9、10、11和BpMYB12。本研究增加了一条研究前期克隆的BpMYB4,共12条。将这12条基因编码的蛋白与107条拟南芥MYB家族基因进行系统进化分析,结果显示可以分为6大组,其中BpMYB1和BpMYB6聚为一组,BpMYB3、10、12聚为一组,BpMYB2、4、5和BpMYB8蛋白序列相似性最高,BpMYB7、9和BpMYB11进化关系较近。BpMYB1与AtMYB61、AtMYB50进化关系比较近,属于R2R3类型的家族成员;BpMYB12与编码叶轴MYB结构域蛋白AtMYB91进化关系比较近;BpMYB4l与AtMYB4进化关系比较近;BpMYB5与种子萌发过程中的原花色素(proanthocyanidin)相关基因AtMYB123进化关系比较近;BpMYB7与影响气孔开关的AtMYB60进化关系比较近;BpMYB11与AtMYB30进化关系比较近;BpMYB9与AtMYB84进化关系比较近(见图3)。
2.3 白桦MYBs基因响应激素处理的表达模式
以不同激素、NaCl和甘露醇处理白桦苗的cDNA为模板,以同期未经处理的白桦苗为对照,利用实时荧光定量PCR技术对白桦中BpMYBs基因的表达量进行分析。
经过20 μmol·L-1ABA的处理后,只有BpMYB4、8上调表达,但差异不显著,其他BpMYB基因全都下调表达,除BpMYB11和BpMYB12下调表达差异非常显著外,其他基因下调的倍数较茉莉酸和乙烯利处理下调的少。经过50 μmol·L-1KT处理后,除了BpMYB11依然处于下调表达以外,其他的BpMYB基因全部呈现出上调表达,其中BpMYB2、8上调倍数最高。经过100 μmol·L-1ETH处理后,所有基因的表达模式出现较为明显的统一趋势,全部都呈现出下调表达,除了BpMYB2与BpMYB8下调表达量差异不显著外,其他的BpMYB基因全都高度下调表达。其中BpMYB11的下调表达量差异最大。经过10 μmol·L-1MeJA处理后, 除了BpMYB2和BpMYB8上调表达

图3 白桦与拟南芥MYB蛋白系统进化分析 拟南芥MYB家族蛋白序列来源于TAIR网Fig.3 Phylogenetic analysis of MYB protein in birch and Arabidopsis The protein sequence of MYB family in Arabidopsis is derived from TAIR website

图4 激素处理下白桦MYBs基因的表达模式分析Fig.4 Analysis of expression pattern of MYBs gene in birch under hormone treatment
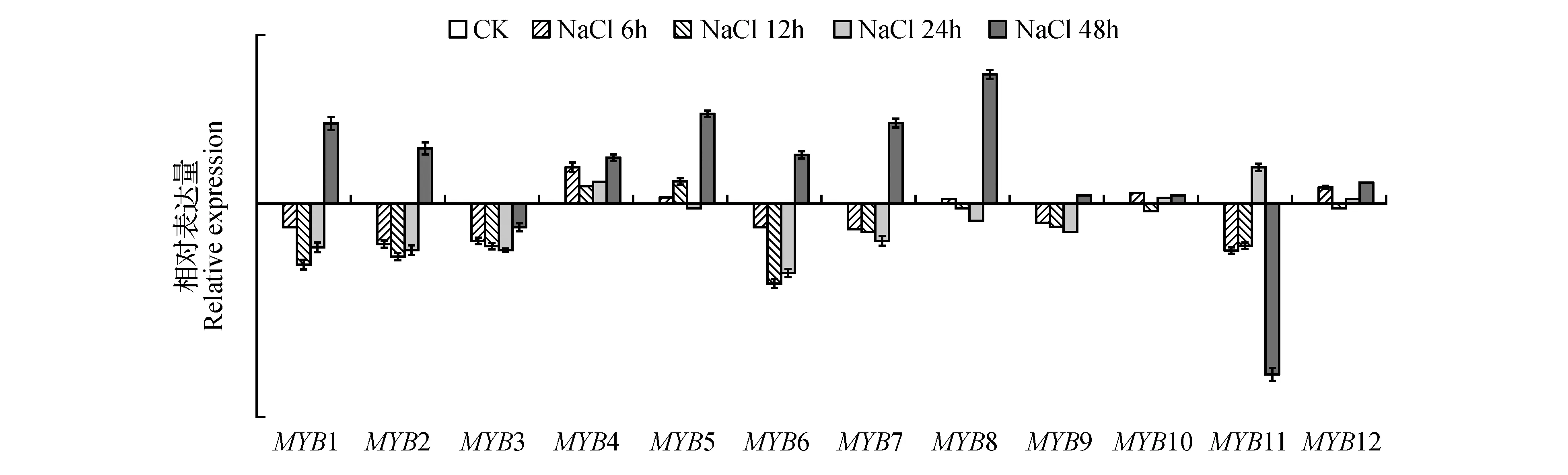
图5 200 mmol·L-1 NaCl处理下白桦MYBs基因的表达模式分析Fig.5 Analysis of the expression pattern of MYBs genes in white birch under 200 mmol·L-1 NaCl treatment

图6 100 mmol·L-1 Mannitol处理下白桦MYB基因的表达模式分析Fig.6 Analysis of the expression pattern of MYBs gene in birch under 100 mmol·L-1 mannitol treatment
外,其余基因全部为下调表达,其中BpMYB5和BpMYB12的下调表达量相对较少,其他基因都呈现出高度下调表达趋势。
经过高浓度NaCl胁迫处理后,BpMYB4基因最先响应胁迫诱导。BpMYB9、10和BpMYB12表达差异不明显。BpMYB11主要表现出显著的下调表达模式,表达量最高可下调317倍。其他大部分基因表现出先下调后上调的表达模式,在48 h表达上升,包括BpMYB1、2、3、6和BpMYB7。BpMYB8主要在处理48 h被诱导。
在甘露醇胁迫处理6 h时,除了BpMYB3、6、8、11外其余基因均为上调表达,但表达量不高,最高的也仅仅为3.1倍;在甘露醇胁迫处理12 h的时候,只有BpMYB3轻度上调表达,其余MYB基因全部都呈下调表达模式,且为高度下调表达;在甘露醇胁迫处理24 h的时候,只有BpMYB11呈高度下调表达,虽然BpMYB1、2、9也呈下调表达,但表达差异较小,其余基因均为上调表达;经过甘露醇胁迫处理48 h的时候,除了BpMYB4、8、10为上调表达外,其余基因均为下调表达,且为高度下调表达,但是,整体下调表达的强度比胁迫12 h的下调表达强度弱。BpMYB11在胁迫12 h后,都呈现出显著的高度下调表达。
3 讨论
在不同激素处理下,白桦4周幼苗表现出了不同的生长表型,ABA和KT处理,能够促进白桦幼苗下胚轴及根的伸长,而ETH处理,显著抑制白桦幼苗下胚轴及根的伸长,但是促进了下胚轴的增粗及侧根的生长。MeJA处理效果不明显。有研究表明高浓度(10-6~10-4mol·L-1)的ABA和MeJA均抑制花生根和下胚轴的生长;低浓度(10-7mol·L-1)的ABA作用不明显,MeJA在低浓度下对生长表现出明显的促进效应[14]。高浓度(10 μmol·L-1)ABA做为胁迫因素在短时间内对2种藜科抗逆植物的种子萌发和幼苗生长均有促进效应,而长时间作用则使2种植物产生不同的适应策略,这种差异可能由于2种种子大小不同所导致[15]。较低温度下(5、10℃),植物材料的发芽率、根长、芽长均随着ABA浓度的提高而降低,20℃时,1、5 mg·L-1的ABA处理对种子萌发均有促进作用[16]。本研究10 μmol·L-1ABA和50 μmol·L-1KT处理对白桦下胚轴和根生长起到促进作用,可能是因为白桦种子的特异性及早期短时间处理造成的。植物激素乙烯(ETH)调节植物生命周期许多方面,如种子萌发、根毛发育、根瘤形成、花衰老、凋谢和果实的成熟。ETH对胚轴伸长的作用依赖幼苗生长在光下或暗中,暗中ETH及ETH合成前体AC(C1-氨基环丙烷1-羧酸)抑制胚轴伸长[17]。本研究白桦种子播种于土壤中进行ETH处理,起到了抑制胚轴伸长的作用。
从整体趋势来看,本研究的白桦MYB基因在4种不同激素的处理下的表达模式的整体趋势基本一致:在10 μmol·L-1MeJA、100 μmol·L-1ETH以及20 μmol·L-1ABA的喷洒处理后,基因大部分都处于下调表达,但是其表达量有所差异,其中在脱落酸处理下的下调表达量较其他两个激素处理后的下调表达量低,而经过乙烯处理后的差异表达量最高,呈高度下调表达模式,这种表达模式暗示了在ETH处理下,BpMYB基因的下调表达,在白桦幼苗下胚轴及根的生长抑制中起作用。在50 μmol·L-1KT的喷洒处理后,基因的表达量全部上调,说明白桦应答KT诱导的BpMYBs基因呈现为正向的调控模式,有研究表明,过表达拟南芥MYBH基因能够促进拟南芥下胚轴的伸长[19~20],本研究中这些上调的BpMYB基因可能在KT处理下白桦幼苗下胚轴及根的伸长中起作用。本研究BpMYBs基因对于不同外源激素信号响应不同,说明它们在不同激素信号途径中起作用。其中有两个表达模式较为特别的基因,一个是BpMYB2,在4种不同激素的处理下都处于一个上调表达的模式,但表达量有所差异,在激动素(KT)的处理下BpMYB2的表达量高度上调,可能与KT处理促进下胚轴及根发育相关;BpMYB2与BpMYB8在激素处理下的表达模式相似,这可能与它们系统进化分析聚为一组相关。另外一个基因是BpMYB11,它与BpMYB2正好相反,在4种不同激素的处理及盐和旱胁迫处理下都处于一个下调表达的模式,而且均处于一个高度下调表达的趋势,尤其是在100 μmol·L-1ETH喷洒处理后下调表达量极高,说明该基因受到外源激素信号的抑制。MYB转录因子在植物抗逆胁迫过程中根据是否依赖ABA信号途径可将其分为依赖于ABA和独立于ABA信号通路两类[14]。本研究发现用20 μmol·L-1脱落酸(ABA)的喷洒处理后,BpMYB11、12表现为强烈的下调表达,说明这两个基因可能依赖于ABA信号通路。在高浓度NaCl胁迫处理情况下,本研究大部分BpMYBs基因在胁迫处理初期下调表达,而在处理48 h被显著诱导上调表达;BpMYB4、5和BpMYB12在盐和旱胁迫处理早期即被诱导,并持续上调表达,可能和之前的信号传导途径有关[18]。总体上,系统进化分析聚为一组的基因有着相似的表达模式。

附表1 BpMYB家族基因克隆的引物序列
猜你喜欢
《学习方法报》小学语文四年级(2023年26期)2023-04-29 00:44:03
热带海洋学报(2022年6期)2022-12-05 12:53:52
环球时报(2022-09-20)2022-09-20 15:18:57
今日农业(2020年24期)2020-12-15 16:16:00
中国稻米(2019年6期)2019-12-05 03:34:22
文苑(2019年20期)2019-11-20 02:12:33
文苑·经典美文(2019年10期)2019-10-16 02:47:16
上海农业学报(2017年4期)2017-04-10 12:40:22
江西农业学报(2017年12期)2017-02-03 21:49:41
兽医导刊(2016年12期)2016-05-17 03:51:50
