
控制水稻中胚轴伸长的QTL 定位
2019-12-05 03:34:22牛世朋吕育松邬亚文魏祥进圣忠华焦桂爱胡时开唐绍清
中国稻米 2019年6期
牛世朋 吕育松 邬亚文 魏祥进 圣忠华 焦桂爱 胡时开 唐绍清
(中国水稻研究所,杭州310006;*通讯作者:sqtang@126.com)
胚轴是构成幼苗形成过程以及植株生长过程中比较重要的部分。在水稻中,中胚轴的伸长能够为种子萌发破土出苗提供动力,提升水稻出苗率。中胚轴的长度在野生稻和栽培稻中的多样性呈现明显差异,具有优良的中胚轴伸长能力的种质将能推动水稻从传统的播种、移秧的耕种方式向节约劳动力和水资源的直播耕种方式的转变。长中胚轴品种一般具有较强的破土能力,这样的品种能够在深覆土的情况下保持较高的出芽率[1-2],有利于水稻直播,大大节省了水稻种植的劳动成本。与传统的栽种方式相比,直播稻栽培主要有以下四个优点:一是省去了准备秧田、育秧和插秧的过程,节约了人力物力,降低了水稻的栽种成本;二是省去了拔秧后插秧的返青过程,缩短了生育期;三是增加了单位面积分蘖,从而大大提高了水稻产量;四是方便进行种植和收获,有利于大规模种植[3]。因此,揭示控制中胚轴伸长的机制可以推动水稻种植向高效、节约型耕种模式转变。
在目前使用的水稻品种和种质资源中,具有长中胚轴特性的材料较少。水稻遗传特性在控制中胚轴伸长中起着决定性作用。林建荣等[4]对水稻中胚轴伸长特性进行了遗传分析,发现中胚轴伸长特性受两对隐性基因控制。杂草稻中的部分品种有较长的中胚轴是一直以来容易被忽略的重要种质资源。杂草稻具有较强的中胚轴伸长能力,充分利用杂草稻中胚轴长的性状对我国的直播稻育种有较大帮助[5]。
关于长中胚轴伸长QTL 的研究已有部分报道。林建荣等[4]研究表明,中胚轴伸长特性受2 对隐性基因控制,并与株高存在一定的连锁关系。曹立勇等[6]利用籼稻IR64 和粳稻Azuccena 产生的DH 群体在第1、3、6、7、8、12 号染色体上共检测到8 个控制中胚轴伸长的QTL。黄成等[1]利用沈农265 和丽江新团黑谷的RIL 群体为材料在水和赤霉素两种条件下在第1、2、3、6、11号染色体上检测到5 个QTL。LU 等[7]利用1 个469 个籼稻材料组成的群体进行SNP 标记进行关联分析,共检测到17 个位点与中胚轴伸长有关。WU 等[8]通过核心种质资源和抗旱材料在黑暗条件下培养,共检测到6 个中胚轴伸长的QTL 位点,并且与前人已定位的QTL 位置吻合,证明了位点的存在。目前已经定位到44 个控制中胚轴伸长的QTL 位点,但至今还未克隆出控制中胚轴伸长的主效基因。
影响中胚轴伸长的因素有很多,包括自身因素和外界因素两大类。马殿荣等[9]研究发现,中胚轴长度与种子内源α-淀粉酶活性、β-淀粉酶活性、可溶性糖含量相关。杂草稻发芽初期淀粉酶活性强,可溶性糖含量高,为中胚轴伸长提供物质能量,导致杂草稻中胚轴明显长于野生稻。除了内部环境外,外界环境对中胚轴的伸长也有很大影响。在所有外界调控中胚轴伸长的因子中,光照影响中胚轴伸长效果最显著。杜金友等[10]研究发现,光对中胚轴的长度有明显的抑制作用,是明显的光调控途径。王莹等[11]研究发现,杂草稻和栽培稻的中胚轴在光照环境下均不伸长,证明了光照对中胚轴的伸长有抑制作用。激素调控途径在中胚轴伸长中也起着重要作用。XIONG 等[12]在日本晴突变体中发现了长中胚轴的突变体gy1,GY1基因通过结合OsEIL2,调控MHZ7/OsEIN2 信号转导通路的下游,进而抑制茉莉酸的生物合成相关基因的表达,促进中胚轴伸长。另外,外界环境会与内源物质协同调控中胚轴的伸长,比如光和油菜素类固醇调控水稻光形态发生从而调控中胚轴伸长[13]。

表1 Asominori 和IR24 群体检测到控制中胚轴伸长QTL
本研究以Asominori 和IR24 重组自交系(RIL)对水稻控制中胚轴伸长性状进行QTL 定位分析,利用Asominori 为遗传背景、籼稻IR 24 为染色体片段供体的片段代换系群体(CSSLs)验证得到的QTL 位点,并与前人的QTL 定位结果进行了比较,旨在为长中胚轴和强顶土能力品种的选育及其分子标记辅助选择(MAS)育种提供一些信息。
1 材料与方法
1.1 实验材料
Asominori/IR24 重组自交系群体(RIL),由172 个家系组成[14];Asominori 为背景的IR24 全基因组片段置换系群体(CSSLs)包含64 个家系,由12 条染色体上的116 个RFLP 多态性分子标记进行辅助选择[15]。两套群体以及分子数据由南京农业大学万建民老师课题组提供。
1.2 实验方法
培养方法参考陈冰孀培养玉米中胚轴方法,并适当修改[16]。
1.2.1 浸种、催芽
每个家系挑选30 粒饱满的种子放在浸种袋中,30℃条件下浸种24 h;取出,用湿毛巾包好,然后30℃条件下催芽24 h,催芽过程中保持毛巾湿润。
1.2.2 固体培养基制备
称取8 g 琼脂放在1L 双蒸水(ddH2O)中,混匀;灭菌锅中121℃灭菌15 min;灭菌完成后冷却到60℃,混匀,均匀倒入8 cm×15 cm 广口瓶中,150 mL/瓶。
1.2.3 播种
挑选20 粒处理好的出芽一致的种子,均匀摆放在固体培养基上。
1.2.4 培养
置于人工气候室内黑暗培养,温度30℃,湿度75%RH。
1.2.5 测量
人工气候室培养7 d,用毫米刻度尺测量各家系的中胚轴长度,并做好记录。去掉偏差较大的数据,选取中胚轴长度集中的10 株然后取平均值作为每个家系中胚轴长度。
1.3 数据分析
利用WinQTLCart 软件[17]对RIL 表型值进行中胚轴伸长QTL 的显著性和定位分析。将参数设置为自动,选择前景控制因子方法,设置排列组合次数为1 000 次来确定LOD 阈值。当实际求得LOD 大于阈值时,就认为该区段存在1 个QTL。将LOD 值2.0 作为判断QTL 存在的阈值,并分析其贡献率、加性效应以及效应来源。遵照McCouch 原则[18]进行QTL 命名。
2 结果与分析
2.1 亲本和重组自交系中胚轴的表型
人工气候室黑暗培养7 d 后,亲本和RIL 群体的生长情况出现明显差异。亲本Asominori 中胚轴长度比较短,仅为0.2 cm;IR24 中胚轴长度较长,达到了0.6 cm(图1a)。在RIL 群体中,中胚轴长度差异比较大,长度从0 cm 到3.5 cm 不等(图1b)。
2.2 水稻中胚轴的QTL 分析
运用WinQTLCart 软件共监测到3 个QTL 位点,分别位于第2、第3 和第7 号染色体上,分别命名为qML2、qML3、qML7,其 对 应 的 标 记 区 间 为RM425 ~RM138、RM7097 ~RM3816 和RM214 ~RM560,LOD 值分别为2.34、3.48 和3.41,贡献率分别为7.25、11.07、8.86。前2 个位点来自于IR24,第3 个来自于Asominori(表1)。
2.3 利用片段代换系验证QTL 位点
本实验检测到的3 个位点中,qML2和qML7在Asominori 为背景的IR24 片段置换的片段代换系群体中有对应的代换家系,qML3没有对应的代换家系。因此利用Asominori 为背景的IR24 片段置换的片段代换系对qML2和qML7进行验证。

表2 代换家系CSSL12 和CSSL13 代换情况及中胚轴表型

表3 代换家系CSSL37 代换情况及中胚轴表型
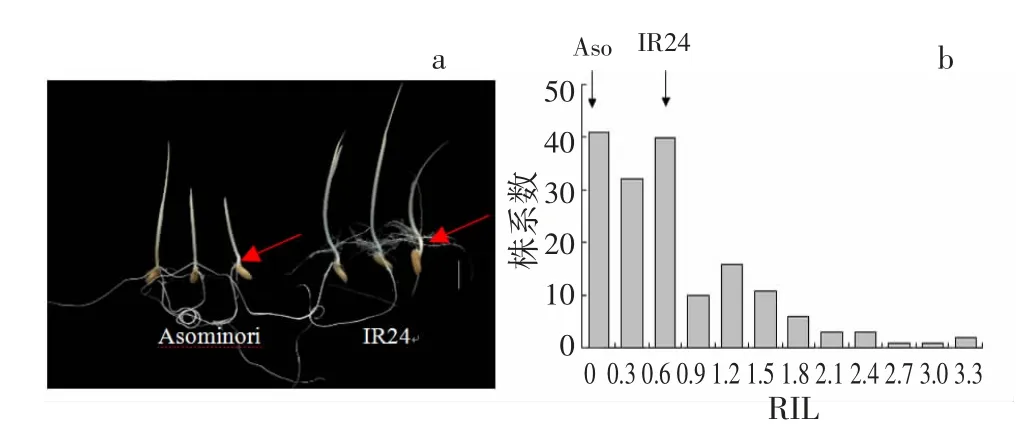
图1 亲本及RIL 群体中胚轴表型
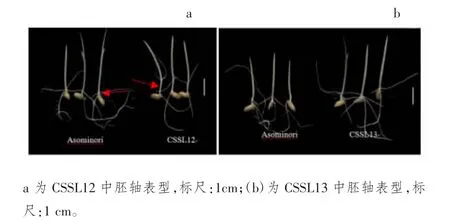
图2 Asominori、代换系CSSL12 和CSSL13 中胚轴表型
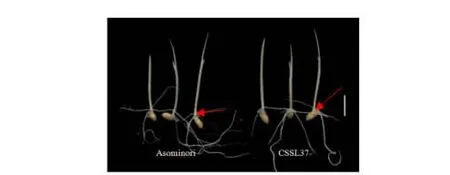
图3 Asominori、CSSL12 和CSSL13 中胚轴表型,标尺:1 cm
2.3.1qML2位点代换系进行验证
在qML2位点,有2 个独立的代换家系CSSL12 和CSSL13 发生了代换(代换情况如表2 阴影部分)。利用2 个独立的代换系验证qML2位点:与亲本Asominori相比,CSSL12 代换系中胚轴极显著伸长(图2a),CSSL13 中胚轴伸长不显著(图2b),说明在RM6~RM425 之间存在控制中胚轴伸长的QTL 位点。
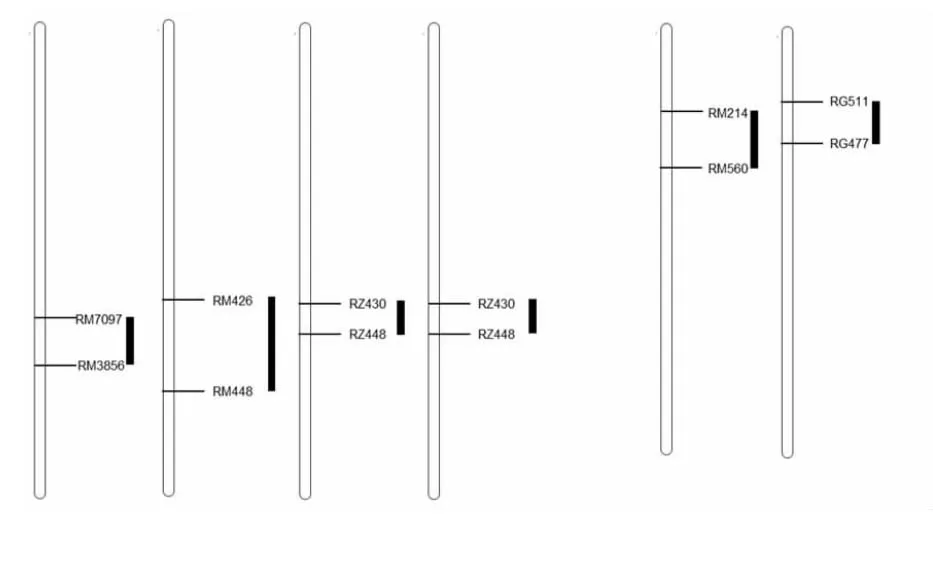
图4 qML3 和qML7 位点与前人研究比较
2.3.2qML7位点代换系进行验证
在qML7位点,CSSL37 发生代换(代换情况如表3 阴影部分)。利用CSSL37 去验证qML7位点的存在:与亲本Asominori 相比,CSSL37 中胚轴伸长受到抑制(图3),中胚轴极显著缩短(表3),说明在RM214~RM346 之间存在控制中胚轴伸长的QTL 位点。
2.4 控制中胚轴伸长QTL 位点与前人研究进行比较
与前人的研究结果比较发现,qML3在不同群体的重复性很好,本研究中定位到的qML3位点与黄成[1]、曹立勇[6]和Redoa[19]位点基本重合(图4 a),说明qML3是一个比较稳定的位点,在不同群体中可以重复检测到,可以作为一个目标位点进行下一步的基因克隆。qML7位点与曹立勇等的结果基本一致[6](图4b),说明了qML7的存在。在先前的研究中,qML2位点未被检测到,利用片段置换系验证qML2位点,qML2位点的片段代换(CSSL12)导致中胚轴极显著伸长证明了qML2的存在,可作为一个新位点进行下一步的基因克隆。
2.5 水稻中胚轴伸长与株高之间的关系
为了验证株高与中胚轴长度之间的关系,考察了2018年杭州大田重组自交系各家系的株高,运用Win-QTLCart 软件进行控制株高QTL 定位。共监测到2 个QTL 位点,位于第2、第7 号染色体上,分别命名为qPH2和qPH7,对应的标记区间为RM71~RM5390 和RM82~RM180(表4)。由控制株高QTL 定位结果可以看出,控制中胚轴伸长位点(图5 黑色部分)与控制株高位点(图5 浅灰色部分)未发生重合,表明在Asominori 和IR24 群体中控制中胚轴伸长性状和控制株高性状具有不同的遗传基础,这将有利于我们在育种中选育长中胚轴且株高适宜的品种。

表4 Asominori 和IR24 群体检测到控制株高QTL

图5 Asominori 和IR24 群体检测到控制中胚轴伸长和控制株高QTL

图6 Asominori 和CSSL12 出土情况(a 为3 d 出土情况;b 为

图7 Asominori 和CSSL12 出芽率
2.6 中胚轴伸长对破土能力的影响
代换系表型考察发现,与亲本Asominori 相比代换家系CSSL12 中胚轴极显著伸长(图2a)。选用Asominori 和CSSL12 家系进行破土实验,检验中胚轴对破土能力的影响。结果发现,与Asominori 相比,代换系CSSL12 出苗更迅速,出苗率更高、苗长势更均匀(图6、图7),说明CSSL12 中片段的代换促进了中胚轴的伸长,进而提升了水稻的破土能力。浅层土覆盖时水稻出苗主要由中胚轴伸长提供动力,本研究与预期结果符合。我们可以通过分子标记辅助方法将控制中胚轴伸长QTL 进行聚合,再结合农艺性状筛选,培养适合直播的优良水稻品种。
3 讨论
水稻是世界上最主要的粮食作物之一,全球一半以上的人口以稻米为主食。但在实际种植过程中,水稻的育秧、取秧、插秧等工作耗费了大量的人力物力,因此水稻直播栽培也渐渐受到重视。水稻直播出苗率与水稻秧苗中胚轴及胚芽鞘的长度有极大的关联[20]。因此,对水稻中胚轴及胚芽鞘伸长机制的研究有极其重要的意义。不同水稻品种中胚轴伸长能力存在差异,既受遗传因素的影响,也极容易受外界环境的影响。影响中胚轴伸长的外界因素主要包括水分、温度、CO2浓度、光照等,以往中胚轴实验是用滤纸放在培养皿里面进行培养,而本实验也革新了先前的培养方式,采用适度硬度的琼脂培养基(0.8%)进行培养,这样更容易控制培养的湿度,排除水分的变化对中胚轴伸长的影响。
水稻中胚轴长度受到基因型与环境共同作用,不同品种之间差异较大且可以稳定遗传,但也因外界环境的变化而产生很大差异[21]。曹立勇等利用籼粳交IR64/Azucena 的DH 群体进行了控制中胚轴伸长的QTL 定位,检测到的qml3(与本实验qML3位点重合)效应来源于中胚轴长的亲本IR64,检测到的qml7-1(与本实验qML7位点重合)效应来源于短中胚轴亲本Azucena,与本实验检测到的qML3和qML7效应来源一致,说明在不同的群体中qML3 和qML7有相同的效应来源。另外,qML2与先前的控制中胚轴伸长的位点未发生重合,且qML2加性效应来源于长中胚轴亲本IR24,在qML2位点的片段代换系验证中,与亲本Asominori 相比,qML2对应的代换家系CSSL12 中胚轴明显伸长,说明IR24 的qML2位点片段的代换导致中胚轴极显著伸长。
已有研究发现,影响水稻中胚轴的基因与控制株高以及控制节间长度的基因可能具有某种关联,这给育种工作带来了一定的困扰[4]。由于株高和节间长度关系到水稻抗倒伏能力的强弱,株高越高、节间长度越长,抗倒伏能力越弱,而株高越矮、节间长度越短的材料一般中胚轴会越短。因此在直播稻育种过程中,选育中胚轴长的材料往往会导致选育品种株高过高而不适合水稻直播。我们利用Asominori 和IR24 重组自交系群体进行株高和中胚轴伸长的QTL 定位时发现,控制株高和控制中胚轴伸长的基因座未发生重合,说明2个性状有不一样的遗传基础,这也为选育长中胚轴表型但株高合理的材料提供了可能。
猜你喜欢
中国糖料(2023年4期)2023-11-01 09:34:46
热带海洋学报(2022年6期)2022-12-05 12:53:52
上海农业学报(2021年4期)2021-08-28 04:13:00
农村百事通(2019年17期)2019-10-08 02:24:55
中国果业信息(2019年11期)2019-01-05 20:47:24
上海农业学报(2017年4期)2017-04-10 12:40:22
现代农业科技(2017年1期)2017-03-06 23:08:18
江苏农业科学(2016年8期)2017-02-15 19:19:29
江苏农业科学(2016年8期)2017-02-15 19:19:15
江西农业学报(2017年12期)2017-02-03 21:49:41
