
康定木兰在成都夏季高温期的光合特性研究
2020-07-09 07:17闫敬林刘光立
四川林业科技 2020年2期
闫敬林,刘光立
四川农业大学风景园林学院,四川 成都 610000
康定木兰(Magnolia dawsoniana)为木兰科木兰属落叶乔木,是我国川西山区特有的一种原始古老孑遗树种,现存数量少,处于濒临灭绝边缘,属国家保护珍稀树种[1]。康定木兰分布区域极窄,主要分布于川西泸定县海螺沟、燕子沟、雨洒坪及石棉县田湾河、九龙县洪坝等贡嘎山周围海拔2 000 m~2 500 m 的湿润沟谷地带[2]。野生康定木兰数量稀少,目前已经很难看到大片树种集中分布的情形,往往只在人迹罕至之地零星点缀,康定木兰处境已不容乐观[3]。在位于崇州的四川农业大学现代农业研发基地内,已经有引种成功的康定木兰。但在进一步的观察研究中发现,引种的康定木兰在成都的夏季高温季节生长受到了一定的胁迫。在成都夏季高温期,康定木兰出现部分新叶干枯,中上部叶片部分出现日灼斑,但是在同期的紫玉兰和白木兰中均未发现这种现象。
伍杰[4]等研究了土壤pH 值对康定木兰幼苗生长的影响,发现pH 值为5.5~7.5 时,康定木兰幼苗的形态特性、株高生长量、地径生长量、叶面积都明显优于其他处理。其中pH 值为6.5 时,康定木兰幼苗的各项所测指标都达到了最大值。pH 值<5.5 或pH 值>7.5 时康定木兰幼苗生长受到明显抑制。在对康定木兰嫁接幼苗年生长规律的研究中发现,采用望春玉兰为砧木嫁接康定木兰,能够明显促进康定木兰的生长[5]。目前对于康定木兰的相关研究还比较少,对于高温适应性的研究更是没有。研究高温条件下康定木兰实生苗和嫁接苗,以及白玉兰和紫玉兰的光合特性指标日变化,比较4 种苗木光合特性的差异,探究康定木兰在成都平原夏季高温期的适应性具有重要意义。
1 材料与方法
1.1 试验地概况
试验地是位于崇州市桤泉镇的四川农业大学现代农业研发基地,地理坐标为东经103°07'-103°49'、北纬30°30'-30°53'之间。崇州属四川盆地亚热带湿润季风气候,年平均气温15.9 ℃,年平均日照时数为1161.5 h,年平均降雨量1 012.4 mm,年平均风速为1.3 m·s-1。平均霜日19 d,平均无霜期为285 d。年平均雪日3 d,且雪量较小。
1.2 试验材料
试验材料为3~5年生康定木兰嫁接苗与实生苗,以及紫玉兰、白玉兰实生苗。康定木兰嫁接苗采用望春玉兰为砧木,嫁接时间为2015年3月1 日,野生康定木兰1~2年生木质化枝条为接穗,采用切接法嫁接,套袋保湿,4月初接穗萌芽10~15 cm 时解除绑扎,进入正常养护管理。4 种试验材料采用相同的栽培条件和措施。
1.3 试验方法
选择天气晴朗高温天,于中午11:00—14:00 时间段,在试验基地内每种材料随机选取3 株生长健壮,无病虫害的苗木进行叶样采集,选择树冠外围中部枝条完整叶片,冰盒保鲜,带回实验室放于-20 ℃冰箱保存用于叶绿素的测定。同时采用Li-6400 便携式光合测定系统,测量这12 株试验材料在8:30—19:30 光合指标的变化。
1.3.1 净光合速率日变化的测定
采用Li-6400 便携式光合作用系统对试验苗木进行光合日变化的测定。取向阳面的中部叶片进行测定,待系统稳定后,同时读取叶片瞬时净光合速率值(Pn)、光照强度(PARi)、蒸腾速率(Tr)、气孔导度(Gs)等相关指标。
从8:30—11:30 每1 h 测定1 次,从13:30—19:30 每2 h 测1 次,每次3 次重复,每次重复记录10 个观测值,最后取平均值。
1.3.2 叶绿素含量的测定
采用张宪政[6]丙酮乙醇混合液浸提法浸提叶片中的色素。将采集回来的木兰叶片清洗干净,用刀片在叶片中部,注意避开主要叶脉,切取0.1 g 叶样。再将叶样切成细丝,放入装有10 mL 的(丙酮:乙醇=1∶1)混合液的离心管中,避光浸提24 h,期间隔段时间摇匀1 次。浸提好后,用分光光度计测定663 nm、645 nm 处的吸光度,重复3 次。按照Amon的方法计算叶绿素a、叶绿素b 的含量。叶绿素a 的含量(mg·g-1)=(12.7D663-2.69D645)V/1 000×W。叶绿素b 的含量(mg·g-1)=(22.9D645-4.68D663)V/1 000×W。式中D633、D645分别为相应波卡下的光密度值,V 为提取液的体积,W 为叶片鲜重或面积。
1.3.3 数据处理
采用Excel 和SPSS 对数据进行处理和绘图。
2 结果与分析
2.1 高温期光合日变化
2.1.1 环境因子日变化规律
研究表明,光合有效辐射(PAR)日变化为单峰曲线,8:30—11:30 持续上升,到11:30 达到峰值1 317.35 umolm-2·s-1,之后开始下降,到19:30 时已经下降至386.55 umolm-2·s-1。大气CO2浓度(Ca)从8:30 的较高水平逐渐降低,而后又从11:30 到19:30 呈持续上升趋势。大气温度的日变化范围在28 ℃到36 ℃之间,其中,从8:30 到15:30 随着时间的推移,大气温度都在持续上升,总共上升了8 ℃,在15:30 达到最大值36 ℃。随后,大气温度开始下降,但是下降的幅度并不大,到19:30 大气温度仍然较高,为34 ℃。大气相对湿度的日变化呈单峰曲线,8:30 开始上升,在10:30 达到峰值79,随后大气相对湿度开始持续下降,在10:30—11:30 时间段下降幅度最大,到19:30 时大气相对湿度为51%。
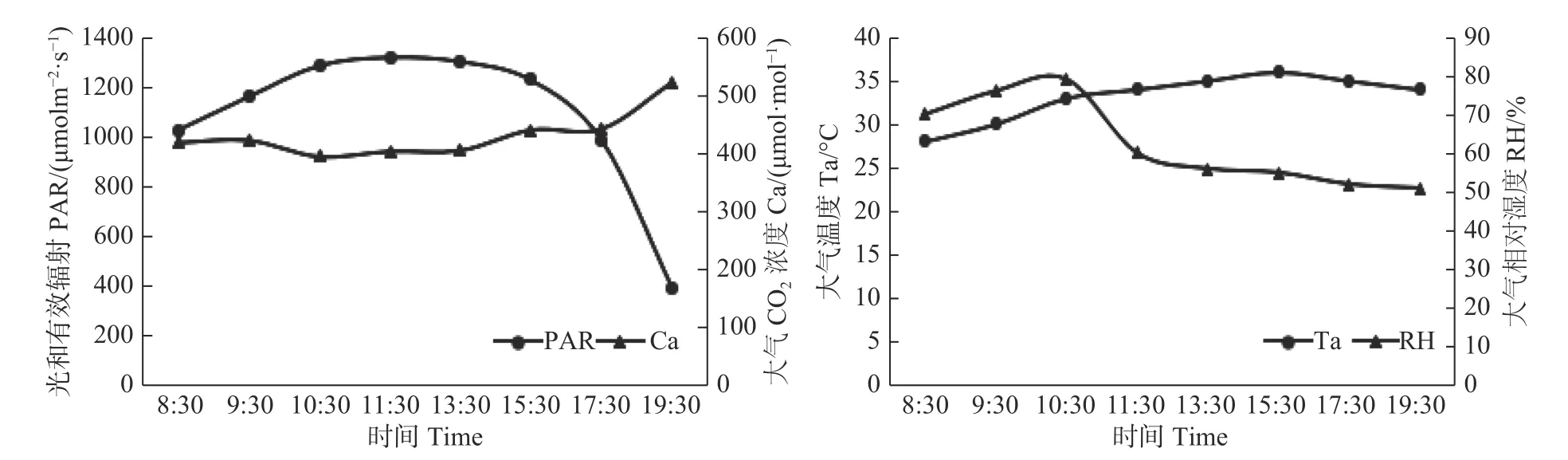
图1 试验地环境因子日变化Fig.1 Diurnal variations of environmental factors in the experimental site
2.1.2 净光合速率和蒸腾速率的日变化特征
研究表明,4 种木兰苗的Pn均值大小关系为:嫁接苗>紫玉兰>白玉兰>实生苗。4 种木兰苗的净光合速率日变化都呈双峰曲线,但是峰值出现的时间有所不同。其中,嫁接苗和紫玉兰净光合速率均是8:30 开始持续显著上升,到10:30 达到第一个峰值,嫁 接 苗 峰 值 为17.80 umolCO2·m-2·g-1,紫 玉 兰 为14.18 umolCO2·m-2·g-1,随后速率开始大幅度的持续下降,到15:30 达到谷值,嫁接苗的谷值甚至为负值。15:30 到17:30 速率上升,达到第二个峰值,17:30到19:30 速率下降,紫玉兰的第二个峰值是四种苗木中最小的,紫玉兰在一天高温的后期净光合速率有显著下降,出现的抑制效果最明显。而实生苗和白玉兰净光合速率则是从8:30 开始上升,到9:30 就已经达到第一个峰值,实生苗峰值为9.58 umolCO2·m-2·g-1,白玉兰为8.58 umolCO2·m-2·g-1。随后,速率持续下降到15:30 达到谷值,实生苗同样出现净光合速率为负值的情况。实生苗在15:30 到17:30 时间段净光合速率上升幅度较大,峰值超越同时段处于峰值的白玉兰和紫玉兰。说明随着高温胁迫的持续,实生苗面对高温胁迫产生了一定的适应性。
蒸腾作用是植物体内的水分以气体的形式散失在大气中的过程,动力来源于PAR,通过影响叶面温度,促进水分的蒸发,Tr在一定程度上反映其调节植物水分损失的能力及适应干旱环境的方式。四种木兰苗的蒸腾速率(Tr)日变化曲线也都为双峰型,且变化趋势基本一致,但是嫁接苗的变化幅度是最大的,嫁接苗两次峰值出现的时间分别是10:30 和17:30。木兰苗在15:30 有明显的谷值,此时Tr降低,可能是树体关闭气孔以达到适应午间高温、防止叶片过度蒸腾水分的一种对环境变化的应对策略。
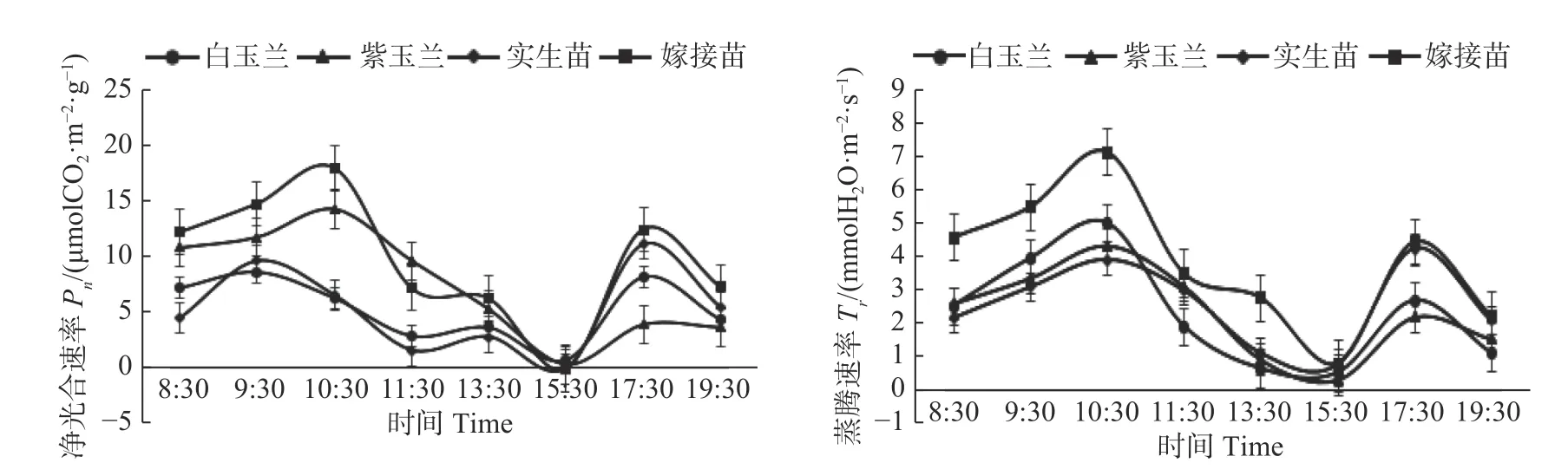
图2 净光合速率(Pn)和蒸腾速率(Tr)日变化Fig.2 Diurnal variations of net photosynthetic rate (Pn) and transpiration rate (Tr)
2.1.3 胞间CO2浓度和气孔导度的日变化特征
Ci是光合作用的重要因素之一,Ci反映了叶片进行光合作用的过程[7],研究表明,康定木兰嫁接苗的Ci浓度日变化呈现先降后升再降再升的双峰曲线,在11:30 达 到 最 小 值297.89 umolCO2·mol-1,在19:30 达到最大值480.03 umolCO2·mol-1,而实生苗的Ci日变化浓度较为复杂,在11:30 达到最小值286.56 umolCO2·mol-1,在19:30 达 到 最 大 值496.57 umolCO2·mol-1。判断叶片Pn降低的原因是非气孔因素还是气孔因素,主要根据Pn、Ci和Ls的变化,当Pn、Ci降低、Ls增大时,Pn的降低主要是由气孔限制因素所引起,当Pn降低,Ci增大、Ls降低时,Pn的降低主要是由非气孔因素限制,也就是叶肉细胞的光合活性下降[8]。嫁接苗和实生苗在10:30 到11:30PPn降低,Ci也降低,Pn与Ci变化方向相同,Ls升高,说明这个时间段Pn下降的原因是气孔关闭。在17:30 到19:30,Pn和Ls均下降,而Ci上升,变化方向相反,说明Pn降低是由于光照强度的降低造成的。
2.1.4 WUE 日变化特征
植物的WUE 指植物消耗单位水量生产出的同化量,是一个比较稳定的衡量碳固定与水分消耗关系的指标[9]。研究表明,四种苗木的水分利用效率日均值大小关系为:紫玉兰>嫁接苗>白玉兰>实生苗,白玉兰和紫玉兰在13:30 的水分利用效率明显高于同时段的嫁接苗和实生苗。嫁接苗的WUE 日变化趋势大致为降-升-降,在15:30 达到最小值,最大值出现在19:30。实生苗的日变化趋势大致为升-降-升-降-升,15:30 后水分利用效率有了明显提升。
2.1.5 净光合速率和环境因子的相关性
植物叶片Pn的大小是衡量植物光合作用能力强弱的重要指标。研究表明,木兰苗的Pn与环境因子的相关性具有较高一致性,其中Pn值均与Ci、Ca呈现显著负相关(P<0.05),而与Tr呈现显著正相关(P<0.05)。嫁接苗的Pn与Ls和WUE 的相关性都要高于实生苗。
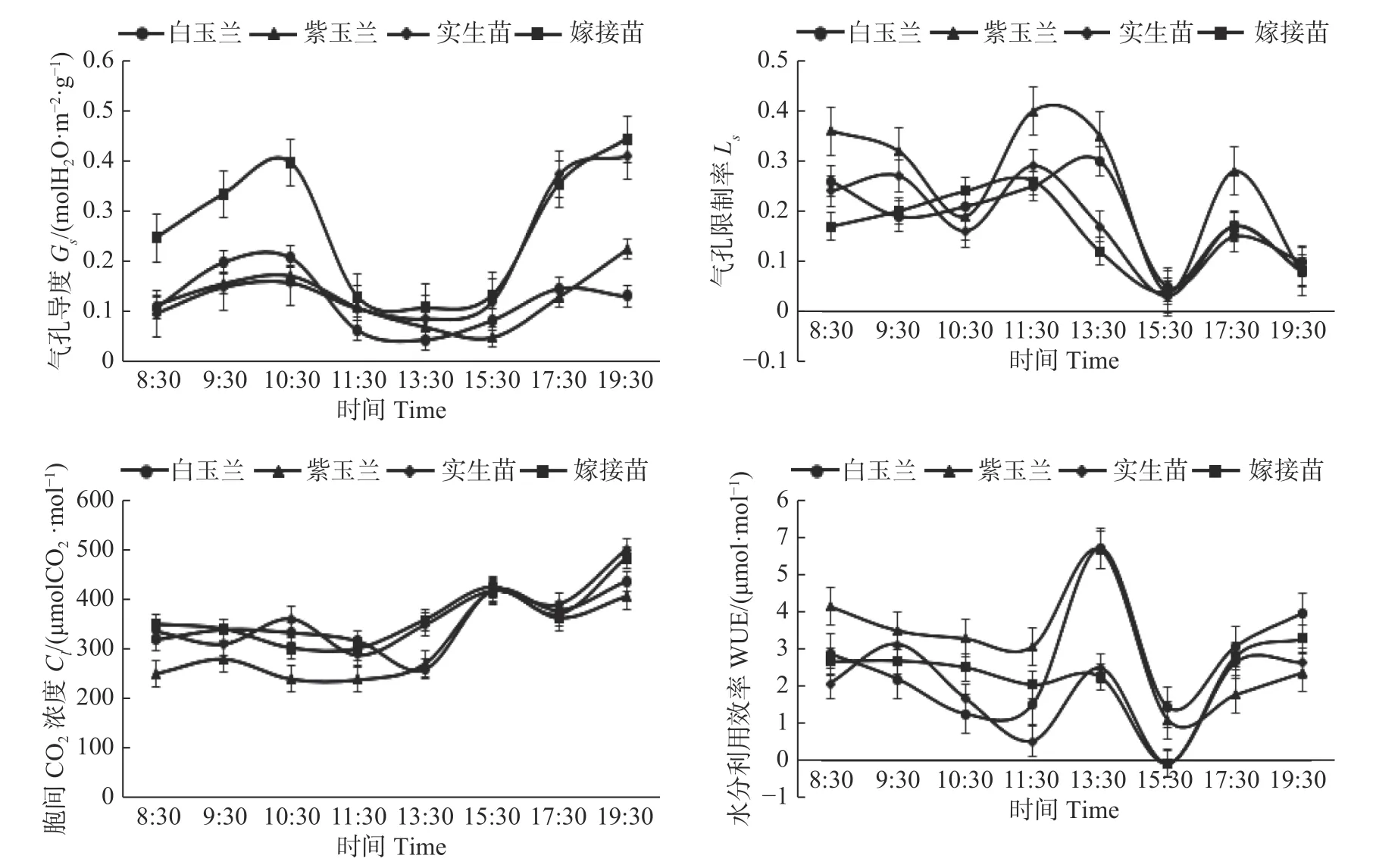
图3 叶片气孔限制值(Ls)、水分利用率(WUE)、气孔导度(Gs)和胞间CO2 浓度(Ci)日变化Fig.3 Diurnal variations of leaf stomatal limitation (Ls),water use efficiency (WUE),stomatal conductance (Gs) and intercellular CO2 concentration (Ci).
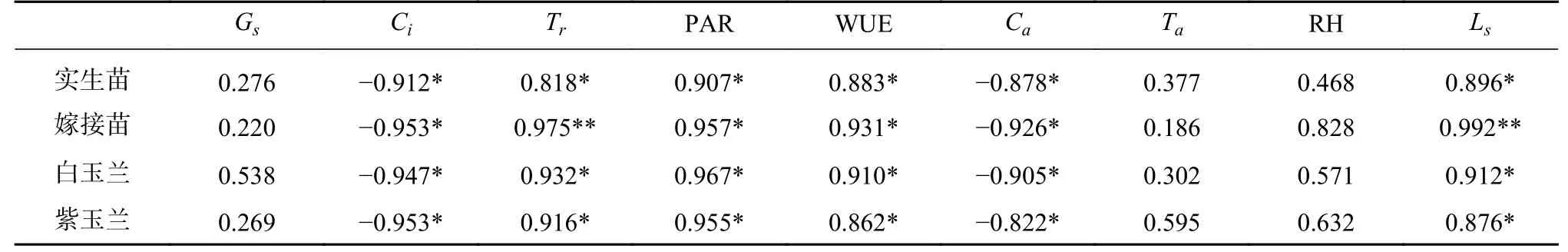
表1 木兰苗叶片净光合速率(Pn)与环境因子相关性Tab.1 Correlation between net photosynthetic rate (Pn) and environmental factors
2.2 高温期叶片叶绿素含量
研究表明,4 种苗木的叶片叶绿素含量关系为嫁接苗>实生苗>白玉兰>紫玉兰。其中,嫁接苗和实生苗的叶片叶绿素含量差别不大,嫁接苗为41.698 mg·L-1,实生苗为39.779 mg·L-1,白玉兰为25.773 mg·L-1,紫玉兰为17.029 mg·L-1。
3 结论与讨论
研究结果表明,在相同的栽培环境中4 种木兰苗的光合特性不尽相同。康定木兰嫁接苗的Pn值最大,4 种木兰苗都有明显的光合午休现象。在中午Pn下降是由于气孔关闭造成的,在17:30 到19:30间Pn值的降低是光照强度的降低导致叶肉细胞活性下降的非气孔因素引起。4 种木兰苗的Tr的日变化与净光合速率呈明显正相关,都为双峰型,嫁接苗的曲线变化更为剧烈,其调节水分蒸腾的能力更强,有利于降低叶温,维持细胞膜的稳定性,能更好地适应高温环境。嫁接苗和实生苗的叶绿素含量较为接近,且均高于白玉兰和紫玉兰。
植物光合作用是植物生产过程中物质积累与生理代谢的基本过程。通常用净光合速率反映植物光合能力的大小,自然条件下,植物光合作用的日变化曲线有单峰、双峰和不规则3 种类型[10]。4 种木兰苗的净光合速率日变化都为双峰型,在中午高温期净光合速率持续下降,说明在高温环境下光合作用均受到抑制。光合作用中的光照强度,空气中CO2浓度、温度、湿度等环境因子与植物适应生态环境存在密切联系,而光合特性是揭示植物对生存环境生态适应性的有效途径[11,12,13]。本研究结果表明,木兰苗的净光合速率与环境因子的相关性具有较高的一致性,蒸腾速率、胞间CO2浓度对Pn 影响较大。
有研究表明,嫁接苗的根系活力、光合作用能力均优于播种苗和扦插苗。如曾义安等[14],郑阳霞等[15]研究发现,嫁接苗有较强的光合能力,其叶绿素含量、净光合速率均高于扦插苗。同时,光合效率与叶绿素含量、叶片成熟度等特性密切相关,植物光合作用的强弱反映了植物本身的适应能力[16]。康定木兰原产于高海拔地区,那里气温常年处于较低水平,植物对高温的适应的能力较差。此研究结果表明,康定木兰嫁接苗的日均净光合速率和叶绿素含量都最大,而实生苗的日均净光合速率最小,说明嫁接在一定程度上提高了植物对高温环境的适应性。在康定木兰今后的引种驯化过程中,嫁接应成为引种中重要的处理方法。
猜你喜欢
中国蔬菜(2022年6期)2022-06-22
车迷(2022年1期)2022-03-29
农业与技术(2021年8期)2021-05-13
地震研究(2021年1期)2021-04-13
河南农业科学(2021年1期)2021-03-09
阿来研究(2020年1期)2020-10-28
阿来研究(2020年1期)2020-10-28
农家顾问(2016年4期)2016-11-19
现代农业研究(2014年9期)2014-08-16
