
高寒沙地不同龄级高山柳光合特性及叶性状研究
2020-07-09 07:17邓东周刘成贺丽鄢武先陈德朝李佳泳
四川林业科技 2020年2期
邓东周,刘成,贺丽,鄢武先,陈德朝,李佳泳
1. 四川省林业科学研究院,四川 成都 610081;
2. 四川省林业和草原调查规划院,四川 成都 610081
荒漠生态系统的灌木对干旱环境的适应能力很强,常作为沙区植被恢复、水土保持、防风固沙的先锋树种,在维持荒漠生态系统物种多样性、稳定性及生态服务功能方面发挥了重要的作用[1-2]。高山柳(Salix cupularis)是杨柳科柳属灌木或小乔木,耐寒抗旱,生存能力很强,在我国环境恶劣的地区,特别是在干旱、沙化严重的流动沙地仍能顽强生长,目前已成为川西北沙化地较理想的防护林带造林树种,对当地的流沙治理和生态环境恢复起到了重要的作用。再者川西北草地沙化日趋严重,就地起沙、风沙侵蚀现象严重,因此对川西北植被保护和恢复的研究刻不容缓。目前国内学者对高山柳的研究主要集中于扦插繁殖试验[3]及干旱河谷交错带高山柳光合生理对干旱胁迫的响应[4],对不同龄级高山柳光合能力及叶性状的差异尚未见报道。
光合作用是植物进行有机物积累和能量转换的主要形式,决定了植物生长发育的基础和生产力的高低[5],光环境在很大程度上决定了植物生长[6-8]。不同生物学特性的植物对光环境具有不同的适应策略,阳性先锋树种具有较强的光合碳同化能力、高光补偿点和高光饱和点等特征[9];阴性树种适应林下弱光照环境,具有较低的光补偿点和较高的表观量子效率,且对林下光斑利用能力较强[10]。近年来关于沙生植物光合作用研究得到了重视,如对梭梭的光合生理生态过程与干旱环境的关系研究表明,梭梭对干旱环境具有良好的适应能力,其净光合速率、蒸腾速率的日变化曲线呈双峰曲型[11];优良固沙植物塔里木沙拐枣随海拔的升高,其最大净光合速率及Pn、Tr、Gs和Ls的日均值降低,但光能利用率(LUE)和水分利用效率(WUE)却显著增加[12]。不同光环境条件下,不同龄级植物光合生理的差异研究日趋受到重视[13-14]。为了明确高山柳光合特性及叶形态特征方面如何与高寒、干旱环境相适应,不同龄级高山柳的光合能力、叶性状差异有多大,以及移栽至流动沙地的高山柳的光合特征、叶性状与苗圃地高山柳的差异。本文以若尔盖县苗圃不同龄级(1年生、2年生、8年生)高山柳及移栽至流动沙地生长的高山柳(11年生)为研究对象,通过测定其自然状态下光合作用参数及叶形态指标,对各龄级高山柳的光合作用参数及叶形态指标进行对比分析。以期深入了解不同龄级高山柳光合能力差异及叶性状的差异,为高寒沙化区高山柳人工林的植被建设、保护与经营提供参考。
1 材料与方法
1.1 研究区概况
研究区地处若尔盖高原,位于四川省的西北部,平均海拔3 500 m,属黄河水系,高原亚寒带湿润气候,冬长无夏,无明显的四季可分。气候严寒,多年平均气温0.7 ℃,最高气温24.60 ℃,最低气温-33.7 ℃,昼夜温差大。11月至来年4月降水稀少,空气干燥,5月至10月空气湿润,多雷雨、冰雹天气。年均降水量657 mm,其中,11月至来年4月降雨量占全年降雨量的14%,5月至10月降雨量占全年降雨量的86%。相对湿度68%,日照时间长,年均日照时数2 300~2 400 h[15]。土壤主要是泥炭沼泽土、高原褐土、草甸土和沼泽土,此外有少量风沙土和人工草地土壤。研究区内的植被主要分为草甸植被、灌丛植被、沼泽植被,此外少量分布着森林植被类型。
1.2 实验材料
研究地点位于若尔盖县苗圃地和若尔盖县阿西乡阿西村协玛坚的流动沙地,研究对象为若尔盖苗圃地不同龄级(1年生、2年生、8年生)高山柳扦插植株和从苗圃地移栽至协玛坚流动沙地的11年生植株。扦插的株行距为15 cm×25 cm,深度40 cm,栽植行距为2 m×3 m。
1.3 光合作用参数的测定
1.3.1 光合日变化的测定
野外测量实验在2014年8月进行,均为晴朗天气。光合日变化测量时间为8:00—18:00,每隔2 h测定1 次。测量时各龄级各5 株,选取树冠南向枝条顶端以下第3 片完整展开叶进行光合参数日变化,每叶片测定3 次[16],每次连续读取10 个稳定的数据,计算其平均值作为测定结果。根据测定的指标计算叶片气孔限制值(Ls)、水分利用效率(WUE)[17-18]。计算公式为:
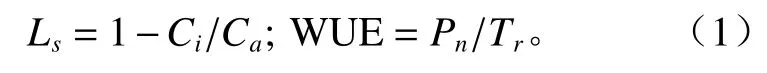
1.3.2 光响应曲线的测定
各龄级高山柳选择3 株各待测样株树冠南向中上部中等大小、健康的叶片(3 片)作为标准叶样。于2014年8月9 日,采用便携式光合作用系统红蓝光源叶室对待测叶片进行光响应曲线的测定。光合有效辐射分别设定为2 000、 1 800、 1 500、1 200、1 000、800、600、400、200、100、80、30、0 μmol·m-2·s-1共计13 个梯度,根据沙漠腹地高温、高辐射的环境气候特点,测定时叶温设置为28 ℃,参比室CO2浓度为380 μmol·m-2·s-1,光合测定仪的流速设定为500 μmol·m-2·s-1。测 定 时 间 为8:30—11:30,测定时以叶的中部作为测试部位,3 次重复,结果取平均值。测定时每一光强下至少停留200 s。然后光合有效辐射(PAR)为横轴、Pn为纵轴绘制光合作用光响应曲线(PAR-Pn曲线),最大净光合速率(Pnmax)、光饱和点(LSP)、光补偿点(LCP)等根据叶子飘直角双曲线修正模型(2010)采用SPSS 13.0 软件进行拟合[19],并计算光响应特征参数,模型表达式为:
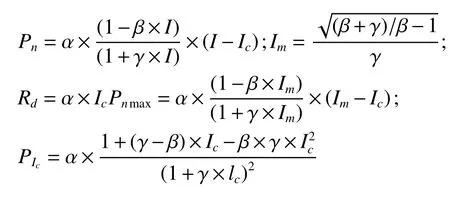
式中:Pn为净光合速率;I为光合有效辐射(PAR);Ic为光补偿点(LCP);α、β、γ为大于零的系数;Im为光饱和点(LSP);Pnmax为最大净光合速率;Rd为暗呼吸速率;PIc为表观量子效率(AQY)。
1.4 叶形态指标的测定
各龄级随机采集10 片完全展开并且健康的叶片,将叶片标记装袋,带回实验室用运用根系扫描仪(EPSON Expression 10000XL,USA)进行叶面积的测定,然后将扫描后的叶片置入水中,4 ℃暗处理12 h 后,迅速用吸水纸吸干叶片表面的水分,在天平(±0.0001 g)上称其饱和鲜重,最后将叶片及剪下的叶柄放入80 ℃烘至恒重,称干重然后计算其比叶面积(SLA,cm2·g-1,为叶面积与叶烘干重的比值)及叶干物质含量(LDMC,为叶干重比叶饱和鲜重)。
1.5 数据处理
采用Excel2007 和SPSS13.0 处理和分析数据,单因素方差分析(One-way ANOVA)进行不同龄级高山柳光合、叶形态指标间的差异显著性检验,若差异显著,采用LSD 法进行多重比较,方差非齐性的用Tamhane’s T2法多重比较。所有图采用SigmaPlot10.0 绘制。
2 结果与分析
2.1 不同龄级高山柳光合生理参数日变化比较
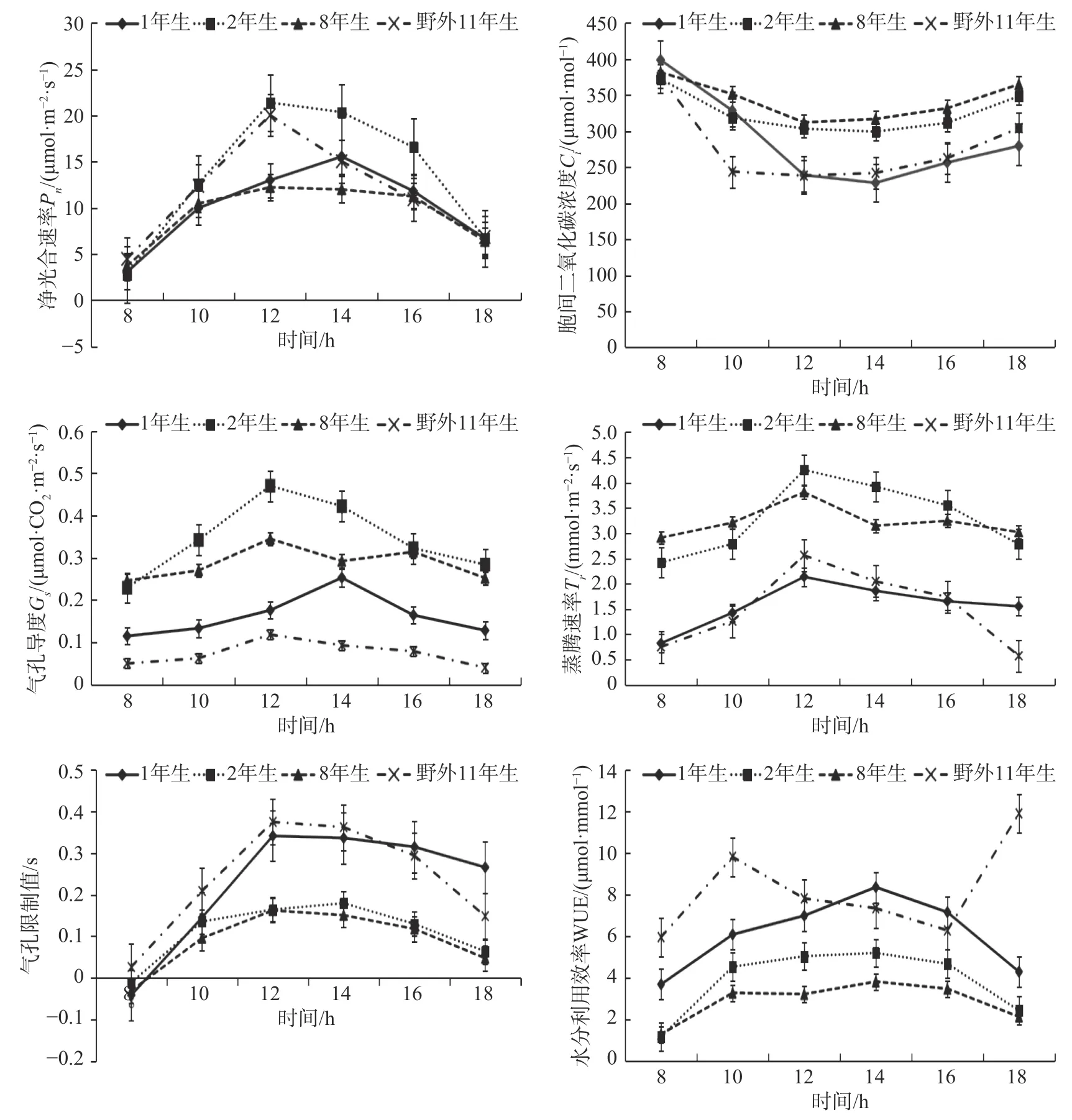
图1 不同龄级高山柳光合生理参数日变化Fig.1 Diurnal variations of photosynthetic physiological parameters of S. cupularis with different stand ages
一天中不同龄级高山柳Pn等光合特征参数随外界环境因子的变化而发生变化,但表现出不同的变化规律(见图1)。各个龄级高山柳净光合速率(Pn)日变化均呈单峰曲线,除了苗圃地1年生植株峰值出现在14:00 外,苗圃地2年生、8年生及野外11年生高山柳均在12:00 左右出现峰值,日均值大小表现为2年生>野外11年生>1年生>8年生。胞间二氧化碳浓度(Ci)值表现为早晚较高,与Pn日变化趋势相反,4 个龄级Ci的日变化趋势一致,但各龄级高山柳Ci最低值大小和出现时间各异,苗圃2年生和8年生日均值大小没有显著性差异,野外11年生与苗圃1年生日均值差异不显著。
各龄级高山柳Tr日变化均呈单峰曲线,但峰值大小不同,2年生峰值最高,为4.25,1年生峰值最低,为2.13。各龄级出现峰值的时间均在12:00 左右。Gs与Tr的日变化趋势一致,且各龄级Gs出现峰值的时间除1年生在14:00 出现峰值外均与Tr相同。
各龄级高山柳Ls日变化曲线均为早晚较低,1年生、8年生、11年生在12:00 左右出现峰值,2年生出现峰值在14:00 左右,且低于苗圃1年生和野外11年生高山柳,与苗圃8年生高山柳Ls差异不明显。WUE 日变化曲线只有野外11年生表现为先升高后降低再升高的趋势,其余3 个龄级均呈先升高后降低的趋势,峰值出现的时间和大小各异,野外11年生高山柳WUE 峰值最高,为11.91 μmol·mmol-1,日均值也显著高于其余各龄级高山柳。
2.2 不同龄级高山柳光响应曲线的差异
光是光合作用的主导因子,对每种绿色植物均可作出光合作用对光的响应曲线。通过光响应曲线,可以确定光LCP、LSP、AQY、Pnmax等指标。强,Pn迅速上升。PAR 大于200 μmol·m-2·s-1时,不同龄级高山柳Pn表现出较大的差异,且上升缓慢,达到一定值后基本稳定。当PAR 达到1 800 μmol·m-2·s-1时,苗圃2年生高山柳Pn值降低,出现光抑制现象,这可能与光照强度大植物叶片叶绿体中系统Ⅱ(PSⅡ)受到损伤,电子传递活性下降有关[20]。
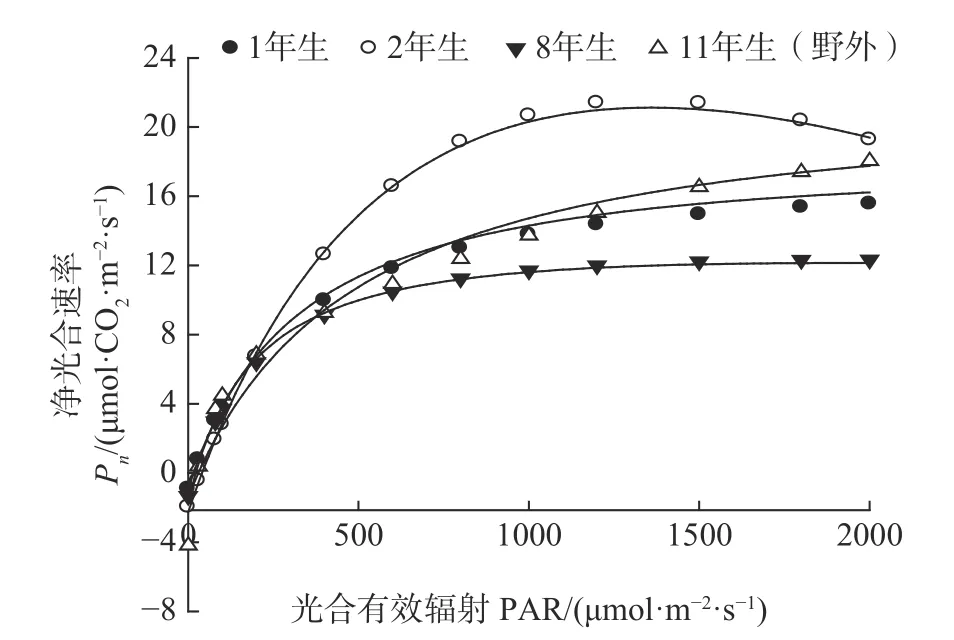
图2 不同龄级高山柳光合-光响应曲线Fig.2 Photosynthesi-Light response curve of S. cupularis with different stand ages
从表1 中可以看出苗圃2年生高山柳的Pnmax显著高于苗圃其余两个龄级,分别比1年生、8年生高山柳高出36.36%、59.17%,但与野外11年生高山柳Pnmax差异显著。苗圃1年生AQY 值显著高于苗圃其余各龄级和野外11年生高山柳,这与其LCP在4 个龄级中最低相吻合,表明对弱光的利用能力最强。11年生高山柳LCP、LSP 值显著高于苗圃各个龄级的高山柳,说明该植物利用强光的能力较强,对弱光的利用能力较弱。苗圃1年生高山柳Rd显著低于其余各龄级,表明其生理活性较弱,有利于有机物的积累。
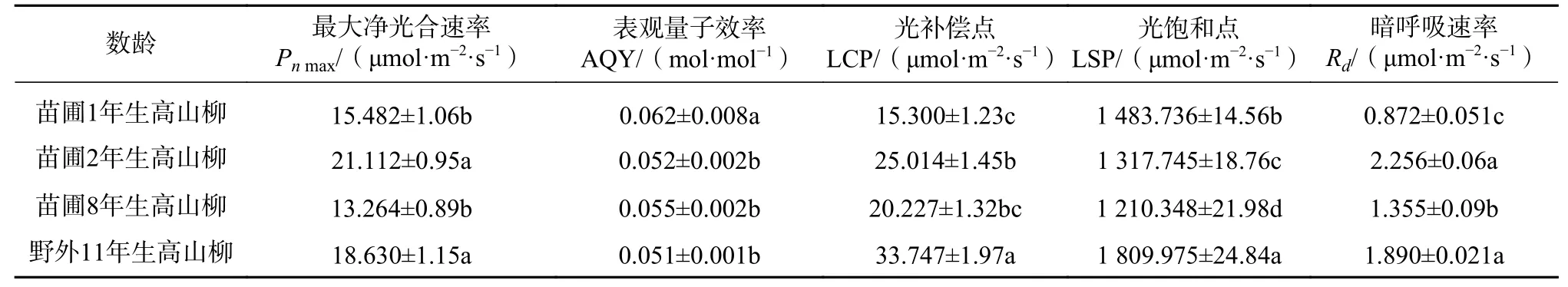
表1 不同龄级高山柳个体光合作用光合作用参数Tab.1 Photosynthetic parameters of light response curves of S. cupularis with different stand ages
2.3 不同龄级高山柳叶片特征
4 个龄级高山柳叶面积、叶干重、比叶面积、叶干物质含量大小如表2 所示。从表中可以看出,除叶干物质含量随年龄的增加呈逐渐上升的趋势外其余叶形态参数均呈先上升后降低的趋势。多重比较表明,野外11年生高山柳叶干物质含量均显著高于苗圃其余各龄级;苗圃1年生与野外11年生高山柳叶面积、叶干重、比叶面积差异不显著,但均显著低于苗圃2年生和8年生高山柳的叶面积、叶干 重、比叶面积大小。
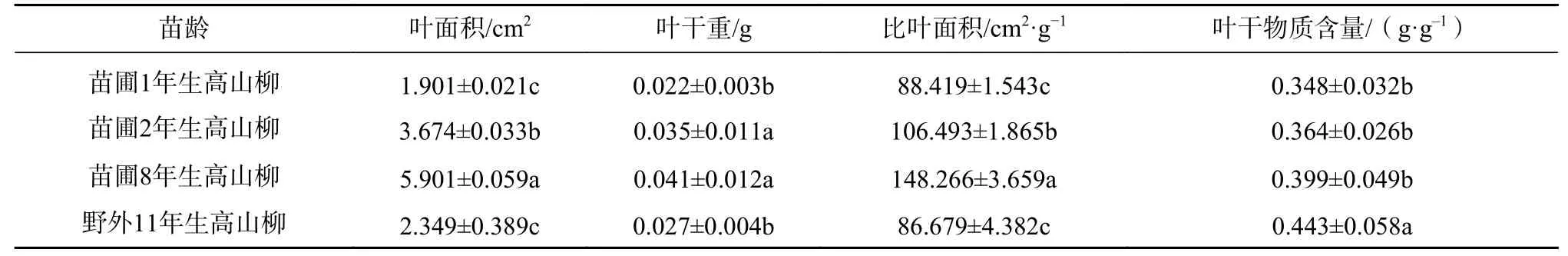
表2 不同龄级高山柳叶形态特征Tab.2 Leaf morphological characteristics of S. cupularis with different stand ages
3 结论与讨论
植物叶片对光能的吸收、传递与利用能力不仅与外界环境因子有关,也与自身生理状态密切相关[21-22]。4 个龄级高山柳的Pn呈单峰曲线,不存在“午休”现象。各龄级高山柳Pn日均值间产生了显著差异,苗圃2年生最高,野外11年生其次,2年生高山柳处于生长快速阶段,积累更多的有机物用于各构件的生长,11年生高山柳生长在流动沙地上,同样有较高的光合速率,表明高山柳能适应干旱的流动沙地环境。
水分利用效率作为植物气体交换的一个重要指标。很多研究认为,荒漠植物具有较高的水分利用效率[23-24]。不同龄级高山柳表现出不同的水分利用策略:苗圃1年生和2年生高山柳WUE 较高,能充分地利用有限水分;野外11年生高山柳也表现出较高的WUE 和较低的Tr,同样也能较充分的利用有限的水分,这与高山柳能在干旱的沙地上较好的生长相吻合;苗圃8年生高山柳Pn与WUE 均处于较低水平,这可能是由于植株冠幅较大,叶片相互遮阴过多,导致其光合速率降低。这种处于不同生长阶段的高山柳所采取各自的水分利用方式是对当地特殊环境的适应,反映了不同龄级高山柳为了维持自身的碳水平衡,获取最大碳收益而采取的不同的生态适应对策。
有研究表明Ci的变化趋势与Pn相反,这与许多植物光合日进程变化规律一致[25-26]。判断植物体Pn降低的原因是气孔还是非气孔因素,主要看Ci和Ls的变化方向:Ci降低、Ls升高表明气孔因素是主要原因;Ci增高、Ls降低表明非气孔因素起主导作用[27]。在午间高温、高光的环境条件下,4 个龄级高山柳Pn均有不同程度的下降,但是降低原因各异:苗圃1年生高山柳14:00 时Pn降低主要是受非气孔因素的影响,即叶肉细胞羧化能力降低或电子传递链受阻造成同化力不足而导致Pn降低,此时植物的光合器官可能已经受到损伤;2年生高山柳12:00 时Pn降低主要是由于气孔限制造成的,14:00 时Pn降低主要由非气孔因素造成的;8年生和11年生高山柳12:00Pn降低主要是由非气孔因素造成的。植物通过调节气孔的关闭程度来适应外界的光环境,这是植物体的一种保护调节机制[28]。
植物叶片的LCP 和LSP 是判断植物耐阴性的一个重要指标。研究发现11年生高山柳LCP 平 均 值 为33.747 μmol·m-2·s-1,LSP 平 均 值 为1 809.975 μmol·m-2·s-1,在自然光强下阳性树种的光补点(平均值30 μmol·m-2·s-1)和光饱和点的范围(1 200-1 400 μmol·m-2·s-1)[29],可初步推断高山柳为阳生性、且耐阴性较强的树种。
研究结果表明野外栽植的高山柳对强光的利用能力较强,且有效光利用区间(LCP 与LSP 之间的差)较广,说明该植物对光的适应范围较广。当PAR 在0~200 μmol·m-2·s-1时,Pn迅速上升,这一阶段主要受非气孔限制,之后随光照强度的增加缓慢上升,这一阶段主要受非气孔限制[30]。AQY 值高,说明叶片光能转化效率高及利用弱光进行光合生产的能力强[31],对于长势良好的植物,AQY 一般在0.04~0.07 之间[32],本研究中高山柳AQY 值在Long等给出的范围值。4 个龄级高山柳Pnmax、AQY 的大小顺序与Pn大小的排列顺序一致,均为苗圃2年生>野外11年生>苗圃1年生>苗圃8年生,进一步证明了苗圃2年生和野外栽植的11 高山柳光合能力较强,同时两者的暗呼吸速率(Rd)较高,叶片代谢能力较强,有利于光合作用。
有研究认为植物SLA 和LDMC 可以反映植物获取资源的能力和对不同生境的适应特征[33-34],且多年生植物两者之间呈负相关性[35]。当SLA 较低时,植物能更好地适应资源贫瘠和干旱的环境;当SLA 较高时,可以更好地适应资源丰富的环境[36-37]。因此,本研究中的11年生高山柳SLA 较低,与其生长在土地资源贫瘠的流动沙地上有关,而LDMC 相对较高,由于其生长年限较长,钙离子和一些次生代谢物则积累较多,导致叶内组织密度增加,导致LDMC 增加,SLA 降低[38]。苗圃1年生、2年生、8年生高山柳SLA 显著上升,随着年龄的增加,通过增加比叶面积,获取更多的光资源用于光合作用,但3 个龄级间的LDMC 差异不显著。
综上所述,高山柳表现出阳性树种的特点,对光的适应范围广,能够适应干旱的流动沙地环境。在苗圃生长时,生长阶段初期,生长发育较快,对弱光的利用能力较强,表现出较低的LCP。这样可以根据不同龄级高山柳的生长策略,合理经营高山柳林。
猜你喜欢
航空材料学报(2022年4期)2022-08-04
贵州农业科学(2022年4期)2022-05-06
浙江农业科学(2022年2期)2022-02-25
干旱地区农业研究(2022年1期)2022-01-28
南方农业(2021年8期)2021-12-26
南方农业·中旬(2021年3期)2021-10-31
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
今日农业(2020年14期)2020-12-14
新疆农业科学(2020年1期)2020-02-14
科技创新与品牌(2019年12期)2019-02-06
