
大花序桉优树家系苗期性状的遗传变异
2020-07-09 07:17陈小中张临萍陈炙刘欢欢黄振李佳蔓杨汉波
四川林业科技 2020年2期
陈小中,张临萍,陈炙,刘欢欢,黄振,李佳蔓,杨汉波,5*
1. 四川省林业工作站,四川 成都 610084;
2. 四川省林业和草原调查规划院,四川 成都 610081;
3. 四川省林业科学研究院,四川 成都 610084;
4. 四川农业大学林学院生态林业研究所,四川 成都 611130;
5. 长江上游林业生态工程四川省重点实验室,四川 成都 611130
大花序桉(Eucalyptus cloezianaF. Muell.),又名昆士兰桉,木材纹理直,基本密度大,材积生长率高,心材黄褐色至浅红褐色,是极具培育价值的中大径材树种之一[1-3]。我国从1972年开始引种大花序桉,1989年建立了大花序桉11 个种源的区域试验林[4]。大花序桉在我国南方的发展空间较大,其最适分布区主要集中在广东、广西、海南和福建沿海[5]。其生长较迅速,材质优良,其木材品质与黄花梨相当,如17年生大花序桉平均基本密度达0.706 g·cm-3,目前已在广西、福建等地广泛用于实木用材林的营建[6,7]。四川省林业科学研究院于1983年开始大花序桉引种试验,在黑龙潭基地,33年生大花序桉优树树高28.5 m、胸径55.5 cm,被筛选为生长良好且极具生长潜力的树种之一。2012年四川省林业科学研究院在宜宾市建立大花序桉种源试验林,6年生试验林平均树高、胸径和单株材积分别达10.2 m、11.4 cm和0.059 m3,表现出相当大的生长潜力,在当前具有重要的研究、开发和推广利用价值。
大花序桉作为实木材和中大径材的培育对象,逐渐成为桉树研究领域的热点[8]。国内外林木育种学家围绕大花序桉开展了一系列的研究,Bootle[9]、Dickinson[10]、Phillips[11]、Muneri[12]等对大花序桉的幼龄林材和成熟材的木材密度进行研究。李昌荣等[6]分析了广西东门林场11 个种源的17年生大花序桉木材基本密度遗传变异。杨汉波等[13]开展5 个大花序桉种源引种的生长性状遗传变异和早期评价。邓紫宇等[14]利用SSR 分子标记技术研究了大花序桉4 个主要分布区的群体遗传多样性,并将其划分为北部和南部两大类。余玉珠等[8]通过木材树皮率、横向全干缩率和体积全干缩率等性状对6年生大花序桉9 个种源木材物理性质在种源、树干高度及径向上的变异规律进行了研究。玉首杰[15]参照红木干燥方法开展了大花序桉用于家具制造的探索,发现其木材含水率达到红木制作家具成品木材的标准,制成的家具高端大气,有红木古典家私的高雅风范,成功解决了家具加工的工艺问题。迄今为止,对大花序桉良种选育主要集中在种源和无性系的选择上,对大花序桉半同胞家系苗期特性还缺乏必要了解。苗期性状及遗传特性是林木良种家系选择的有效途径,具有加快育种进程、提早获得林木改良经济回报、方法简单、投产迅速且增益较高等优点[16,17]。因此,本文以27 个半同胞家系的大花序桉1年生苗为研究对象,通过对大花序桉半同胞家系苗期生长性状的测定,比较不同半同胞家系苗期生长指标的差异,旨在初步筛选出优良的大花序桉半同胞家系,以期为大花序桉优良家系的苗期选育提供一定的理论依据。
1 材料与方法
1.1 试验材料
试验地位于四川省成都市郫都区唐昌镇现代化育苗基地(103.779041°E,30.937384°N),属亚热带季风性湿润气候,夏无酷暑,冬无严寒,雨量充沛。年均气温16℃,降水量979.4 mm,日照1 014 h。
供试的27 个大花序桉半同胞家系于2017年10月全部采自宜宾市6年生大花序桉引种试验林的优良单株(见表1),该大花序桉种源试验林种源由澳大利亚CSIRO 中心提供,6年生时试验林生长良好,保存率在79%以上,平均树高、胸径和单株材积分别为10.2 m、11.4 cm 和0.059 m3[13]。大花序桉种子即将成熟且少部分已脱落时采集。在2018年3月,将采集的大花序桉种子采用随机区组实验设计分别进行播种育苗。幼苗出土后,适时除草、松土,按常规方法进行育苗和苗期管理。
1.2 性状调查方法
生长指标的测定方法:2019年7月对大花序桉1年生苗的株高(H)和地径(DGL)的生长状况进行测量。测定时在每小区随机选取10 株植株,重复3 次,用钢卷尺和游标卡尺分别测量苗木的H 和DGL(距地面大约2 cm 处的幼苗直径)。
1.3 数据统计分析方法
使用EXCEL 2013 进行数据分析处理和图表制作。使用林木单地点半同胞子代测定分析软件HalfsibSS 1.0 计算每个性状的方差分量、方差分量的假设检验统计量、家系遗传力、单株遗传力和性状间的遗传关系[18]。
统计分析模型为:

式中,yijk为第i个区组第j个家系第k个单株的数量性状值;μ为总体平均值;Bi为第i个区组的固定效益,i=1,2,3,…,b;Fj为第j个家系的随机效应,j=1,2,…,f;BFij为第i个区组第j个家系的随机效应;eijk为第i个区组第j个家系第k个单株的随机误差。
家系重复力采用公式:R=1-1/F,式中,F为方差分析的F值[19]。
变异系数:CV=SD/×100%,式中,CV为变异系数,SD为各性状的标准差,为各性状均值。
一般配合力(GCA)采用公式:g=x-μ,式中,g为亲本的一般配合力,x为亲本的某个交配组合在某个性状的子代平均值,μ为这个性状所有组合的子代总平均值。
用隶属函数法综合各项指标进行评价[20],隶属函数值计算公式为:

式中,i表示某个家系,j表示某项指标,Tij表示i家系j指标的隶属函数值,Xij表示i家系j指标的测定值,Xjmin表示所有家系j指标的最小值,Xjmax表示所有家系j指标的最大值。某一个体某一指标的隶属函数值越大,表明该指标越靠近最大值。
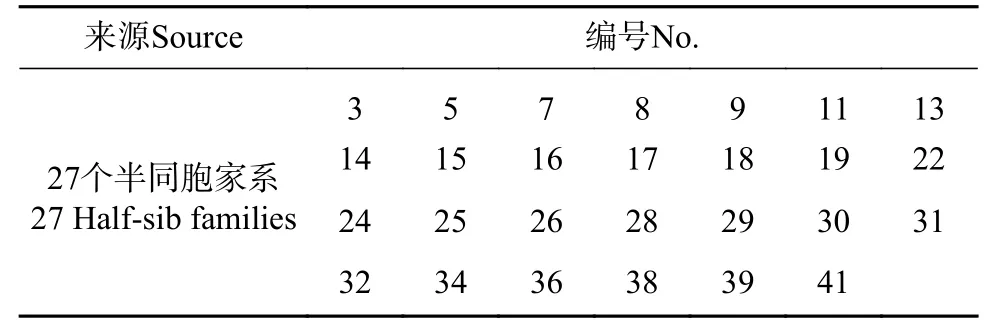
表1 试验材料Tab.1 Experimental materials
家系现实遗传增益估算公式:
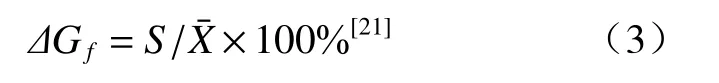
式中,S为选择差,为各性状平均值。
单株遗传增益估算公式:ΔGs=(R/)×100%R=h2·S[21]。式中,R为选择反应,S为选择差,表示各性状平均值。
2 结果与分析
2.1 生长性状的变异分析
参试的27 个家系苗高(H)和地径(DGL)性状调查情况(见表2)表明:苗高平均9.12 cm,地径平均3.06 mm,其最小值仅为1.37 cm 和0.78 mm,最大值为27.00 cm 和18.00 mm。苗高和地径表型变异系数分别高达44.01%和65.96%。说明1年生大花序桉家系各性状间存在极为丰富的遗传变异,在家系和单株水平上具有巨大的遗传改良潜力。
2.2 生长性状的方差分析及遗传参数
表3 表明:大花序桉苗高和地径在家系间差异极显著。苗高和地径具有较高的重复力,分别为0.895 和0.985,这表明大花序桉家系生长性状遗传变异显著,受较高强度的遗传控制。性状间遗传相关分析结果表明:苗高和地径的遗传相关两两之间呈负相关关系,但不显著(r=-0.5778,P>0.05),说明大花序桉苗期苗高和地径性状可能是独立进行遗传。苗高和地径均具有高的家系遗传力,均超过0.8,地径的家系遗传力高达0.985。从单株遗传力来看,苗高和地径的单株遗传力均相当大。

表2 参试家系各性状测定值Tab.2 Measured values of various traits in the tested families

表3 大花序桉家系各性状方差分析及重复力估算Tab.3 Variance analysis and estimation of repeatability of traits in E. cloeziana F. Muell. families
2.3 各性状一般配合力分析
各性状一般配合力(GCA)见表4。苗高一般配合力变化范围为-6.989(家系19)-5.909(家系38),其中18 个家系的一般配合力为正值,9 个家系的一般配合力为负值。地径一配合力变化范围为-1.151(家系34)-7.777(家系39),其中5 个家系的一般配合力为正值,22 个家系的一般配合力为负值。6 个家系的苗高一般配合力较高,均在2.0 以上。2 个家系的地径一般配合力较高,分别为3.777(家系19)和7.777(家系39),其余均在1.0 以下。其中家系13 和25 苗高和地径一般配合力均较高,其亲本可选作优良亲本材料。
2.4 优良家系选择
为选育生长量大的家系,采用隶属函数法共筛选出5 个家系39、19、38、25 和13(见表5)。与整体平均值相比,筛选出的家系生长性状都有不同程度的提高,其中苗高的增益最大为64.75%,地径的增益最大为254.47%(见表6)。家系39 和家系19 的地径增益巨大,分别为254.47%和123.59%,但它们的苗高增益为负值,可用作培育高胸径生长量潜力优良家系的定向培育,或作为优良的杂交亲本,以获得其优良的地径生长性状。
3 讨论和结论
遗传变异是选择的基础,对变异来源、特点和规律的研究是进行树种改良并获得遗传增益的理论前提[22,23]。陈益泰[24]综合分析了不同树种、指标的评价结果,认为在林木苗期开展早期选择是有效的、可行的。苗高和地径是反映苗木质量最直观的
指标[25]。本研究通过对1年生27 个大花序桉家系生长性状进行统计分析发现,苗高和地径在家系间差异极显著,说明家系水平具有较大的遗传改良潜力。生长性状遗传力、重复力估算结果表明,苗高和地径受高强度的遗传控制,家系生长遗传改良具有可行性。这与桢楠、白桦及麻栎苗期评价和选择研究的结论基本保持一致[26-28]。

表4 各家系不同性状一般配合力Tab.4 General combining ability values of different traits among different families

表5 大花序桉家系不同性状隶属函数值及排序Tab.5 Membership function values of different traits of E. cloeziana families
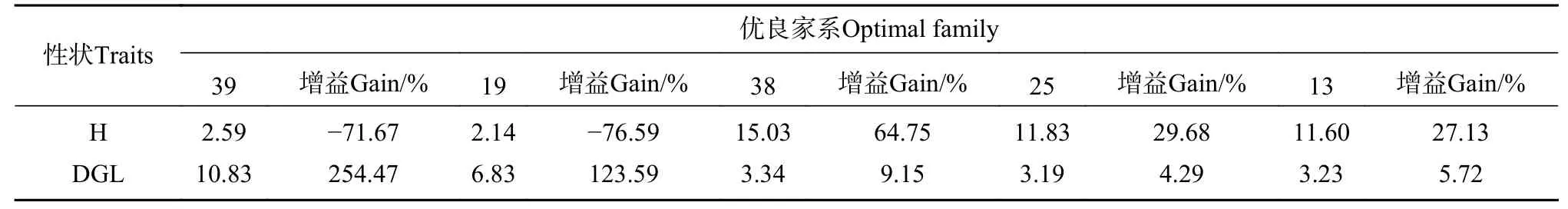
表6 综合选育的优良家系特征描述Tab.6 Comprehensive selection of optimal family characteristics
遗传力表示亲本某一性状遗传给子代能力的大小,是估算遗传增益的重要参数[29]。本研究中,苗高和地径家系遗传力均高于6年生大花序桉种源[13]、1年生赤桉家系[30],所有性状的家系遗传力均高于0.8,属高遗传力,表明所选材料各性状能稳定遗传[31]。大花序桉苗高和地径的单株遗传力分别为0.985 和2.935,均超过杨汉波等[13]对6年生大花序桉种源、尚秀华等[30]对1年生赤桉家系的研究结果。单株遗传力较高,表明可在家系内进行优良单株的筛选,可为优良种质的发掘、优良亲本的选配和种子园营建提供材料[32]。本研究中各性状单株遗传力均高于家系遗传力,明显高于赤桉、邓恩桉种源、家系的研究结果[30,33],表明各性状在单株水平上的遗传能力强于家系,单株在各性状的遗传差异具稳定的遗传能力。配合力反映亲本优良性状传递给子代的相对能力,其大小与具体性状有关[34]。除家系25 和13 各性状一般配合力均较高,其亲本可作为优良亲本材料外,其余家系不同性状一般配合力高的家系差异较大,难以进行联合选择,需结合隶属函数法进一步对家系进行评价选择。家系39 和19 地径性状一般配合力较高,家系38 苗高性状的一般配合力较高,可作为特异亲本或材料加以选择、开发。
育种目标决定育种方向,大花序桉是极具培育价值的锯材树种,但作为中大径材培育的周期较长,为缩短育种周期,选育高生长量的优质大花序桉资源,本文结合大花序桉实际情况,采用隶属函数法对各家系进行苗期评价。本研究在隶属函数法评分的基础上,选择分数靠前的5 个家系作为优良家系,入选率为18.5%,其中家系39 和19 苗高和地径遗传增益形成两个相反的极端,地径遗传增益极高,分别为254.47%和123.59%,而苗高遗传增益分别为-71.67%和-76.59%,这两个家系可考虑作为特异优良亲本培育,用于杂交育种有望获得高杂种优势的杂交种质。家系38、25 和13 苗高和地径均存在较高的遗传增益,高于红松[32]、桢楠[26]、麻栎[28]等针、阔叶用材树种,表明利用苗期生长性状选择对大花序桉进行遗传改良具有较好的效果,入选的家系在生长上存在明显的优势,若用优良家系进行生产造林,有望减少林分工艺成熟时间,进而提高经济效益[35]。
1年生大花序桉家系苗高和地径性状存在丰富的遗传变异,且受到高的遗传控制,有非常好的遗传改良潜力。综合生长指标结合隶属函数法,初步选定家系39、19、38、25 和13 为优良家系。本次试验是对1年生大花序桉幼苗进行测定和评价,其结果仅为大花序桉优良家系早期选额提供参考,初选的可靠性和稳定性还有待对区域造林试验的表现进行跟踪观测和验证分析。然而,本次研究至少为大花序桉早期选择提供了一定的理论依据和创新思路。
猜你喜欢
养猪(2022年4期)2022-08-17
养殖与饲料(2021年11期)2021-11-15
园艺与种苗(2021年8期)2021-09-23
农技服务(2020年1期)2020-12-17
山东农业科学(2019年11期)2019-12-24
现代农业科技(2019年1期)2019-07-11
现代农业科技(2017年24期)2018-01-22
湖北畜牧兽医(2015年11期)2016-01-11
湖北畜牧兽医(2015年11期)2016-01-11
园艺与种苗(2015年8期)2015-12-28
