
鸢尾素对兔骨髓间充质干细胞成脂分化的影响
2020-07-06 09:57:54董科明郭红延徐红梅申叶春李金儒
武警医学 2020年6期
翟 羽,董科明,郭红延,刘 翠,徐红梅,申叶春,李金儒,梅 稳
干细胞是组织工程的种子细胞,在体外具有分化成包括成骨细胞和脂肪细胞在内等多种细胞的潜能。干细胞的分化方向是由它所处的微环境所决定的[1]。BMSCs是一种组织工程中常用的种子细胞,有多条信号通路调控BMSCs的分化[2,3],其中Wnt/β-catenin信号通路的作用尤为重要。激活Wnt/β-catenin信号通路可促进BMSCs成骨分化并抑制其成脂分化[4]。BMSCs的成脂分化又受到多种转录因子的网络调节,其中比较重要的转录因子包括氧化物酶体增殖物激活受体-γ(peroxisome proliferator-activated receptor-γ, PPARγ)和CCAAT/增强子结合蛋白α(CCAAT/enhancer binding protein α, C/EBPα)等[5],而PPARγ和C/EBPα又均受到Wnt/β-catenin信号通路的调控[6]。鸢尾素是新近发现的一种由运动诱导产生的肌源性细胞因子,在棕色脂肪组织中高表达[7]。最近有研究报道,鸢尾素可促进BMSCs成骨分化[8-10],且可抑制脂肪组织的分化、成熟[11]。但是,鸢尾素对BMSCs成脂分化的影响及其作用机制尚不清楚。因此,本研究拟以体外培养的兔BMSCs为研究对象,观察鸢尾素对BMSCs脂向分化的影响并探讨其信号机制。
1 材料与方法
1.1 实验材料、试剂与仪器 3个月龄雌性新西兰大白兔1只,购自军事科学院军事医学研究院实验动物中心。重组人鸢尾素(美国Phoenix Pharmaceuticals公司);α-MEM培养基、高糖DMEM培养基、胎牛血清、青霉素、链霉素(美国Gibco公司);胰蛋白酶(美国amresco公司);消炎痛、3-异丁基-1-甲基黄嘌呤(IBMX)、普通牛胰岛素、地塞米松、L-谷氨酰胺、二甲基亚砜(DMSO, 美国Sigma公司);油红O、异丙醇(上海国药集团);Trizol(美国Invitrogen公司);Wnt10a抗体(货号:(#sc-376028; 美国Santa Cruz公司);BCA测定试剂盒(南京建成);PVDF膜(美国Millipore公司)。
Herocell C1型CO2恒温培养箱(中国润度生物);BCM-1000A型生物净化工作台(苏州安泰);5702R型低速离心机、Biophotometer核酸蛋白测定仪(德国eppendorf公司);Multiskan型酶联仪(美国赛默飞·世尔公司);DYY-7C型电泳仪、DYCZ-40型转膜仪(六一仪器厂);LW300LFT型正置荧光显微镜(日本奥林巴斯公司)。
1.2 细胞增殖能力检测 取第2代兔BMSCs接种于96孔板,并将细胞分成2组:对照组仅采用含10%胎牛血清的α-MEM培养液培养;鸢尾素组则在α-MEM培养液中加入100 ng/ml的重组鸢尾素,剂量参考既往报道[8,11]。接种后1~11 d,每天采用MTT法检测BMSCs的增殖活性,检测方法参照文献[12]。
1.3 qPCR检测成脂相关基因的转录表达 将第3代BMSCs以1.0×104/孔的密度接种于24孔板,待细胞长满瓶底面积70%时,换为自行配制的成脂诱导培养液(在高糖DMEM培养基中加入10%胎牛血清、1000 nmol/L地塞米松、10 mg/L 牛胰岛素、60 μmol/L消炎痛和0.5 mmol/L IBMX)培养,同时加入0 ng/ml(对照组)或者100 ng/ml的鸢尾素(鸢尾素组)。 两组细胞分别成脂诱导培养7、14 d后提取总RNA,再将RNA反转录为cDNA并进行扩增。然后,检测PPARγ和C/EBPα等成脂相关基因的转录表达。扩增条件为:95 ℃预变性3 min,96 ℃变30 s,58 ℃退火30 s,73 ℃延伸45 s,扩增30个循环,最后73 ℃ 延伸10 min。基因引物由上海申工生物合成,序列见表1。

表1 成脂相关基因引物序列
注:GAPDH为内参
1.4 油红O染色及定量分析 对照组和鸢尾素组BMSCs在成脂诱导14 d后,倒掉诱导液,PBS缓冲液漂洗干净,10%中性甲醛固定30 min,然后进行油红O染色并于显微镜下拍照。最后,用异丙醇振荡洗脱至油红O脂滴全部脱落,并于490 nm处测定吸光度值。
1.5 Western blot检测Wnt10a的表达 对照组和鸢尾素组BMSCs在成脂诱导7 d后,依据Western blot实验的常规步骤[13]裂解细胞并提取总蛋白,凝胶电泳分离蛋白、转膜、显影后,采用Image J软件分析胶片灰度。

2 结 果
2.1 鸢尾素对BMSCs增殖活性的影响 BMSCs的增殖曲线呈S形,细胞在前5 d增殖缓慢,第6~7天增殖最快,此后细胞生长处于平台期。而鸢尾素对BMSCs增殖无明显影响(图1),表明鸢尾素无细胞毒性。
2.2 鸢尾素对BMSCs成脂相关基因表达水平的影响 qPCR检测结果显示:在BMSCs成脂分化的第7天,与对照组相比,鸢尾素明显抑制成脂相关基因PPARγ[(0.20±0.06)vs(1.00±0.10)]和C/EBPα[(0.40±0.06)vs(1.00±0.12)]的转录表达(P<0.05);甚至在BMSCs成脂分化的第14天,鸢尾素仍能显著抑制PPARγ[(0.61±0.06)vs(1.00±0.15)]和C/EBPα[(0.70±0.09)vs(1.00±0.18)]的转录表达(P<0.05)。
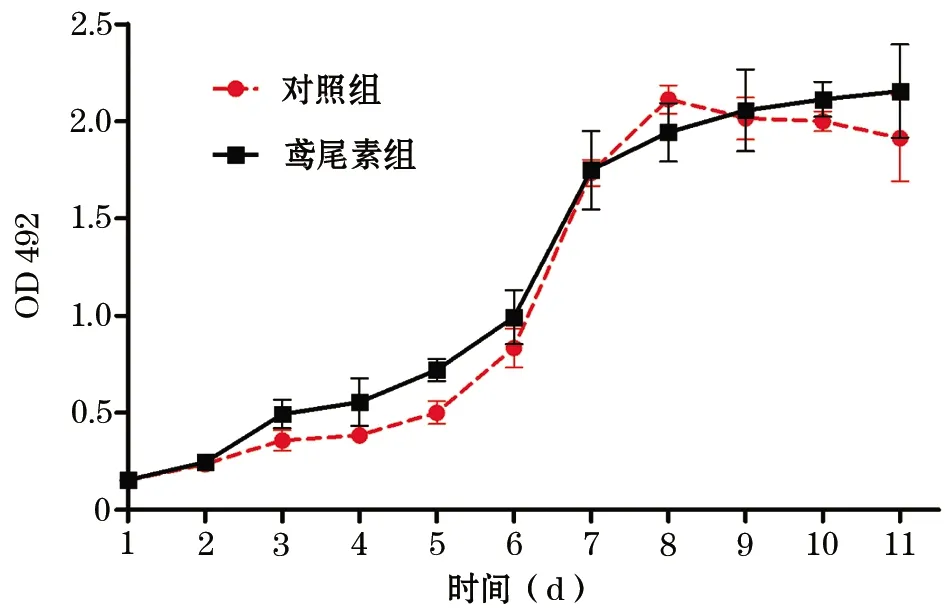
图1 BMSCs增殖曲线
2.3 鸢尾素对BMSCs成脂分化的影响 对各组BMSCs成脂诱导14 d后行油红O染色,对照组细胞中可见大小混合型的脂滴布满整个视野,而鸢尾素组细胞中脂质沉积明显减少。进一步对细胞中的油红O进行半定量分析,结果也证实鸢尾素组油红O的OD值显著低于对照组,差异有统计学意义[(0.45±0.05)vs(0.95±0.10);P<0.05]。
2.4 鸢尾素对Wnt10a蛋白表达的影响 Western blot结果显示:在BMSCs脂向分化的过程中,鸢尾素能显著提高Wnt10a蛋白的表达水平,与对照组相比差异有统计学意义[(5.01±0.78)vs(1.00±0.25);P<0.05]。
3 讨 论
本研究以兔BMSCs为细胞模型,发现在BMSCs成脂分化的过程中,鸢尾素能显著提高Wnt10a信号蛋白的表达,进而下调成脂相关基因PPARγ和C/EBPα的转录表达,并最终抑制BMSCs成脂分化。
运动是预防肥胖和代谢性疾病的有效策略,不仅仅是因为运动能直接增强骨骼肌的强度,而且运动能诱导骨骼肌产生肌源性细胞因子,例如鸢尾素、白细胞介素-15和成纤维细胞因子21等[14]。这些细胞因子通过内分泌或旁分泌等方式调节机体的代谢,对机体器官的代谢紊乱起到间接的保护作用。因此,肌源性细胞因子是联系骨骼肌和机体其它系统的重要纽带[15]。鸢尾素是运动诱导骨骼肌产生的一种非常重要得细胞因子,它依赖激活PGC1α起作用[16]。分泌型鸢尾素的主要功能是通过上调解偶联蛋白1(UCP1)的表达,促进白色脂肪棕色化[7],进而通过增加能量代谢来缓解代谢性疾病[17]。例如,有研究表明,鸢尾素能通过减少脂肪酸合酶的生成抑制人脂肪细胞内脂质的沉积[18]。本研究发现,鸢尾素能抑制BMSCs成脂分化。这间接地佐证了干细胞分化平衡理论,即促进干细胞向成骨细胞分化就会抑制干细胞向脂肪细胞分化,因为鸢尾素能促进BMSCs向成骨细胞分化已被广泛证实[8-10]。与本研究相似,有实验表明,鸢尾素也能抑制成熟的脂肪细胞分化[19]。
经典的Wnt信号通路是调控成骨与成脂的重要信号通路[20]。有研究报道称,鸢尾素可以通过下调sclerotin(一种Wnt/β-catenin信号通路的特异性抑制剂)来促进骨生成与矿化[21]。本实验发现,在抑制BMSCs成脂分化的过程中,鸢尾素能促进Wnt10a的合成。当然,在经典Wnt信号通路还存在其它配体,如Wnt10b和Wnt6等,未检测鸢尾素对其它配体表达的影响是本实验的一个局限性。Wnt信号通路调控成脂关键转录因子PPARγ,C/EBPα和脂肪酸结合蛋白4(fatty acid binding protein 4, FABP4)的表达,PPARγ和C/EBPα主要调控成脂早期的细胞分化,而FABP4主要调控成脂末期的细胞分化。本实验发现,无论在成脂早期(7 d)还是终末期(14 d),鸢尾素都能显著抑制PPARγ和C/EBPα的转录表达。新近有研究表明[11],无论是内源性还是外源性的鸢尾素均能抑制脂肪组织的成熟、分化,并且鸢尾素的抑制作用与激活Wnt信号通路相关,这与本研究的结果一致。
鸢尾素是纤维连结蛋白Ⅲ型域包含蛋白5(type III domain-containing protein 5, FNDC5)的分泌形式,FNDC5体内超表达可以显著升高循环中鸢尾素的水平[22],而直接在BMSCs中超表达FNDC5能否抑制其自身成脂分化值得进一步探讨。本研究未进行剂量-反应实验,因此并不清楚其它干预剂量的鸢尾素对BMSCs成脂分化的影响,后续研究应设计多个干预剂量。
综上所述,本研究发现鸢尾素能够在体外抑制兔BMSCs脂向分化,并且鸢尾素的这种作用与激活Wnt信号通路有关。鸢尾素有望被开发成抑制BMSCs脂向分化,进而促进BMSCs成骨分化的一种新型药物。
猜你喜欢
江南诗(2023年4期)2023-08-06 19:54:11
中老年保健(2021年7期)2021-08-22 07:44:28
甘肃科技(2020年20期)2020-04-13 00:30:48
中成药(2017年10期)2017-11-16 00:49:54
中成药(2017年10期)2017-11-16 00:49:52
中小企业管理与科技·中旬刊(2017年4期)2017-05-16 19:42:19
课程教育研究·学法教法研究(2017年1期)2017-03-21 23:40:21
罕少疾病杂志(2016年2期)2017-01-11 12:27:34
华南农业大学学报(2016年5期)2016-08-05 05:35:04
山西省政法管理干部学院学报(2016年2期)2016-07-31 18:19:32
