
湿地松产脂量与生长及树冠性状多点遗传相关及通径分析
2020-07-01 00:24:44冷春晖孙世武蒋祥英
核农学报 2020年7期
冷春晖 张 露 易 敏 孙世武 蒋祥英 赖 猛
(1江西农业大学,江西特色林木资源培育与利用2011 协同创新中心/江西省森林培育重点实验室,江西 南昌 330045;2吉安市白云山林场,江西 吉安 343062;3景德镇市枫树山林场,江西 景德镇 333000)
产脂量是衡量松属(Pinus)树种产脂能力非常重要的指标,受树体因子、林学因子和气象因子等诸多因素的影响[1]。目前,在生产上采脂工选择高产脂植株仍较盲目,通过了解产脂量与其他性状内在的相关性,可以进行松脂性状与其他性状的间接选择或联合选择,大大提高良种选择的效率。研究表明,产脂量与树体表观形态因子紧密相关[2]。随着高产脂选育研究工作的推进,产脂量与生长性状、树冠性状等表观形态因子之间的关系正逐渐成为育种学家关心的重点问题[1,3]。如胸径可作为思茅松(P.kesiyavar)、马尾松(P.massoniana)和湿地松(P.elliottii)等松类高产脂优树野外选择的主要指标[4-6]。树冠性状中冠幅、冠长、树冠轮廓与产脂能力之间均存在较高的遗传相关性,这3 个因子对产脂量的影响程度依次增强,当树冠比率在40%~50%时,松脂产量和树木生长量将维持最佳状态,树冠长度与树高之比低于1/3 时松脂产量明显减少[5,7-8]。此外,树皮厚度、分枝角、侧枝粗度等因子也是影响产脂量的重要因素,且与产脂量存在显著的线性关系[9-10]。
湿地松原产于美国东南部,在我国已有70 多年的引种历史,其松脂主要由松节油和松香组成,具有质量好、松香不易结晶、杂质少、β-蒎烯含量高等特点,广泛应用于工业生产、化妆品、食品和医药等领域[11-12]。近年来,我国已开展湿地松产脂量的遗传测定和产脂量相关因子分析等工作,但现有研究方法仍存在许多不足,如在产脂量预测模型的建立上,大多以单个指标或单类性状预测为主,预测精度不足,效率较低;全年称脂测量法虽然可获得精准的产量数据,但消耗的人、财、物力过大,不适宜大规模推广。因此,系统掌握湿地松产脂量与树体因子间的关系及其控制途径,并以此为基础建立产脂量预测模型,既是选育湿地松高产脂优树的前提,也是科学开展湿地松采脂林定向培育的基础。本研究以赣北、赣中和赣南等多种典型立地条件下的28年生湿地松家系试验林为研究对象,应用遗传相关及通径分析的方法研究生长性状和树冠性状与产脂量之间的相关关系及其对产脂量的遗传控制途径,构建主要性状与产脂量之间的关系模型,为脂用目标湿地松优良品种选育提供间接评价因子,这对加快湿地松高产脂选育进程和提高育种效率具有重要意义。
1 材料与方法
1.1 试验林概况
试验地分别位于景德镇枫树山林场、吉安白云山林场和赣州九龙林场,各试验地基本概况如表1所示。试验林均采用随机区组设计,均包括同样的112 个从美国引进的高产脂湿地松家系,分5 个区组,4 次重复,1989年春育苗,1990年春造林。
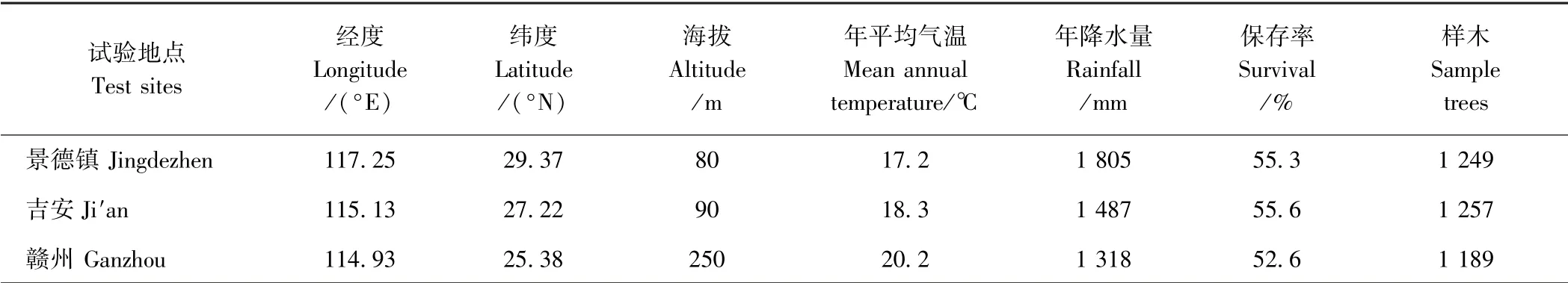
表1 试验地基本概况Table 1 Basic information of the test sites
1.2 试验测定及方法
2016年8月,对三片试验林进行全林产脂量(resin yield,RY)、生长及树冠性状的测定。采用下降式单刀采脂法进行产脂量的测定,割脂高度在样木约1.5 m 处,倾斜树干呈45°割面,负荷率约为40%,侧沟深入木质部5 mm,步距2 mm,割脂频率两天一刀。8月共割脂15 刀,月末收脂并记录每株林木的单株产脂量,称量时先将松脂收集袋中的水倒出,然后将松脂收集袋挂在弹簧称上直接称量。测量生长性状:树高(height of the tree,HGT)、胸径(diameter at breast height,DBH)、枝下高(height under live crown,HLC);树冠性状:冠幅(crown width,CW)、冠长(crown length,CL)、树冠表面积(crown surface area,CSA),其中树冠表面积由冠幅和冠长计算得出[13]:
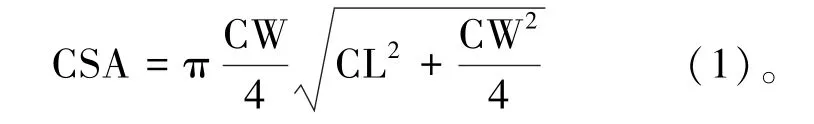
1.3 建模方法
利用线性混合模型(mixed linear model,LMM),进行多点遗传分析,单变量分析模型如下[14]:
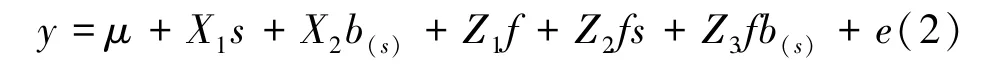
式中,y是观测值向量,μ表示该性状观察值的总体平均值向量,s、b(s)是固定的地点和地点内区组效应向量,f、fs、fb(s)和e是随机的家系、家系-地点交互、家系-地点内区组交互及残差效应向量。X和Z分别是固定和随机效应的关联矩阵。模型假设f~ N(0,,其中和分别是家系、家系-地点交互、家系-地点内区组交互及残差方差,Ⅰ是单位矩阵。
以多点单变量遗传分析模型为基础,利用扩展的双变量模型进行性状间的遗传相关分析,模型如下:
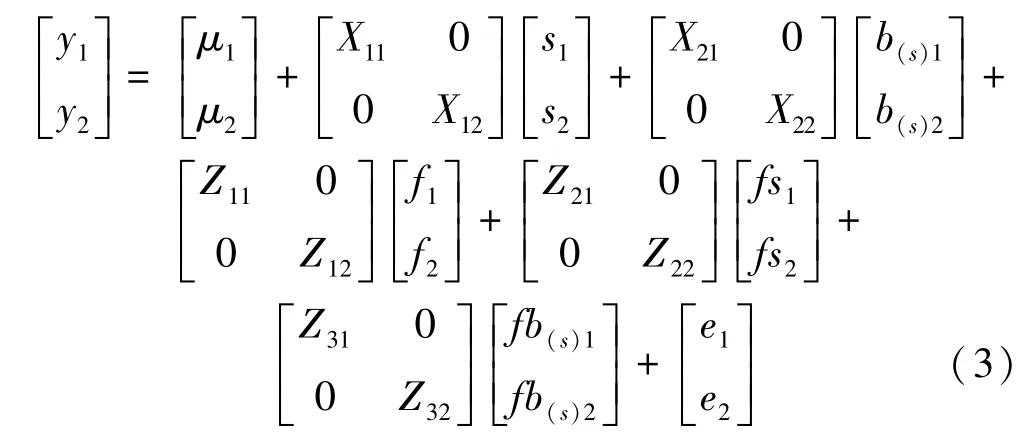
利用公式计算相关系数[15-16]:
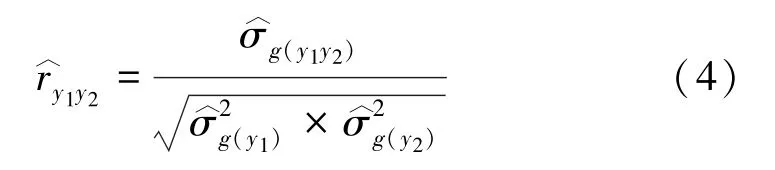
模型拟合及参数估计均通过ASReml-R(V3.0)统计分析软件进行[17]。其中,方差估计采用限制性最大似然估计法(residual maximum likelihood,REML)[18]。利用t检验进行相关系数显著性的假设检验,其零假设是相关系数r=0。
基于遗传相关系数,利用最小二乘法进行生长及树冠性状与产脂量的遗传通径分析[19]。其分析过程如下,设有变量x1,x2,…,xn与y,它们之间存在着线性关系,其回归方程为:
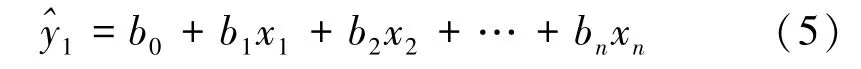
利用最小二乘法求方程组,求得性状x1,x2,…,xn对性状y 的直接通径系数p:

式中,[A]为各控制性状x1,x2,…,xn与性状y的遗传相关系数向量,[B]为各控制性状间的遗传相关系数矩阵。

式中,qiΠj为xi通过xj对性状y的间接通径系数;rij为xi与xj的遗传相关系数;pj为xj直接通径系数。
根据通径分析结果,确定对产脂量贡献率最大的因子,应用SPSS 22.0 建立多元回归模型,并对模型进行预估精度检验[20]。其中,每个家系随机挑选两组数据,112 个家系,3 个试点共672 组数据作为模型的检验样本,其余数据作为拟合样本,模型预估精度检验公式如下:
式中,yi为实测值,为预估值,n为样本数,tα为置信水平为0.05 时t的分布值,T为回归模型中的参数个数,为预估值的均值。
2 结果与分析
2.1 湿地松家系试验林各测定性状的表型变异特征
由表2可知,景德镇、吉安和赣州(由北向南)3 个试验点的产脂量均值分别为1.17、1.16 和1.25 kg,赣州产脂量均值大于其他两试验点,且与景德镇和吉安的产脂量均值间呈显著差异,吉安产脂量最低但变异系数最高达35.57%,说明该点产脂量具有更大的选择潜力。胸径均值由北向南逐渐增加,分别为25.26、27.82 和28.05 cm,赣南点(赣州)的年均气温和无霜期的天数均较其他两试验点高,这可能是其胸径均值更大的原因之一;树高、枝下高、冠幅和树冠表面积均值均以景德镇为最大,而冠长均值则以吉安表现最优。
2.2 产脂量、生长及树冠性状间的遗传相关性分析
由表3可知,除枝下高外,产脂量与各性状间均呈显著或极显著的遗传正相关关系;其中,与产脂量遗传相关系数排名前二位的性状分别为胸径(R=0.93)和树冠表面积(R=0.83),说明胸径和树冠表面积对产脂量的影响要高于其他因子,在进行间接选择时,胸径和树冠表面积越大,单株产脂量越高。除个别性状外,生长性状与树冠性状呈显著或极显著遗传正相关关系。在各树冠性状的相关关系中,整体来看,冠幅、冠长和树冠表面积三者间均呈极显著遗传正相关关系,枝下高与冠幅及树冠表面积的遗传相关系数不显著且与冠幅呈负相关。
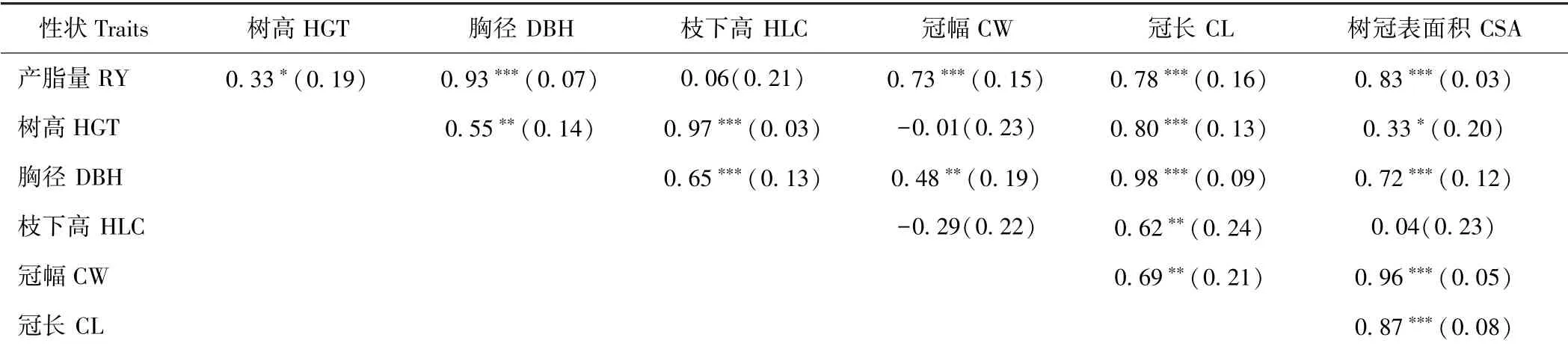
表3 产脂量、生长及树冠性状间的多点综合遗传相关分析Table 3 Estimated intertrait genetic correlations among resin yield, growth traits and crown traits from site combined analysis
2.3 生长及树冠性状与产脂量的遗传通径分析
由图1 和表4可知,在湿地松生长及树冠性状与产脂量的通径分析中,不同生长及树冠性状对产脂量的直接控制作用不同。树高对产脂量的直接作用很强烈,直接通径系数高达1.537,但绝大部分被反向间接作用(间接通径系数达-1.207)所抵消,故两者相关系数仅为0.330。同样,冠长对产脂量具有很强烈的直接作用(直接通径系数为-1.098),然而冠长通过影响树高、胸径、冠幅及树冠表面积等性状对产脂量产生了更加强烈的间接正效应,因此冠长与产脂量呈现出正相关关系,且两者关系达极显著水平。枝下高对产脂量的直接作用和间接作用大小非常接近,但方向正好相反,因此,两者的相关系数很小,仅为0.060。胸径、树冠表面积和冠幅的直接通径系数分别为0.599、0.511 和0.329,间接通径系数分别为0.331、0.319 和0.401,直接作用和间接作用均表现出相同的正向效应,因此这3 个性状与产脂量均表现出极显著正相关关系。
由表5可知,6 个生长及树冠性状对单株产脂量影响明显,在各性状之间的相互作用下,其对产脂量的决定系数贡献之和为0.820,同时亦说明还有其他因素影响林木的产脂量,如生长环境、松脂的分泌与运输机制及控制松脂合成基因的选择性表达等因素。由各性状对决定系数的贡献程度排序可知,胸径(0.557)、树高(0.507)、树冠表面积(0.424)和冠幅(0.240)通过直接作用和间接作用成为影响产脂量的主导因素。
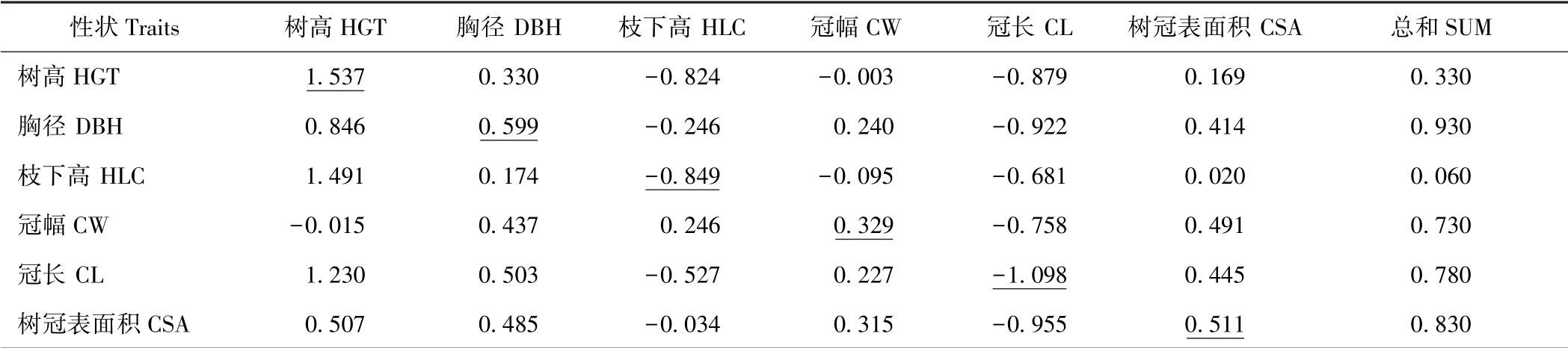
表4 生长及树冠性状对产脂量的遗传通径系数Table 4 Genetic path analysis coefficients of growth and crown traits on resin yield
2.4 产脂量与生长及树冠性状的多元回归分析
根据通径分析结果,得出对产脂量影响最大的4个因子分别为胸径、树高、树冠表面积和冠幅,将这4个因子与产脂量之间进行多元回归分析。每个家系随机挑选两组数据,112 个家系,3 个试验点共672 组数据作为检验样本,其余数据作为拟合样本;利用拟合样本建立产脂量与胸径、树高、树冠表面积和冠幅的回归方 程:y=- 0.255 +0.040x(DBH)+0.004x(HGT)+0.006x(CSA)+0.036x(CW),对回归方程进行显著性检验(F=955.907∗∗,R2=0.559),回归常数和回归系数均达到极显著水平。用672 组检验样本对上述方程进行预估精度计算,得到预估精度为99.64%,超过95%,因此应用该模型预测湿地松的产脂量是可行且可靠的。
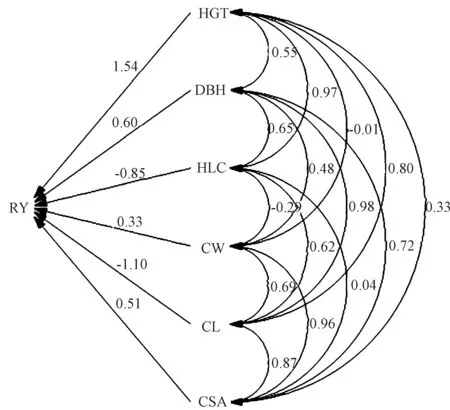
图1 生长及树冠性状对产脂量的通径图Fig.1 Path map of resin yield by growth and crown traits
3 讨论
近年来,随着松脂价格逐年上涨,湿地松松脂产业为林农带来了丰厚的经济回报。然而,目前松脂生产仍主要靠大面积人工采脂来完成,采脂工作强度大、劳动效率低。虽然高产脂选优工作已有一定进展,但在选优过程中,因无可靠鉴别高产脂湿地松的因子,采脂工选择高产脂采脂树时具有较大的盲目性,选择效率低,误选率较高。如果采取全年称脂测量法,虽然可以提高选择精度,但会消耗大量的人力、物力及财力,严重阻碍高产脂湿地松选育研究进展。本研究以湿地松多点家系试验林为研究对象,具有广泛的代表性,由不同试点产脂量可知,湿地松单月单株产脂量均值在1.16~1.25 kg 之间,与马尾松、海岸松(P.pinaster)等松类树种产脂量略有差异[10,21],造成此现象的原因可能是松属内不同树种间的种间差异,此外,立地环境、取样年龄、取样方法也会对产脂量产生重要影响[22-23]。整体来看,赣南(1.25 kg)的产脂量高于赣北(1.17 kg)和赣中(1.16 kg),这主要是因为赣南的年平均气温高于赣中和赣北,而年降雨量又低于这两点,适度的高温和干旱能够提高产脂树种分泌松脂的能力[21,24]。湿地松生长及树冠性状在赣北、赣中和赣南3 个典型立地条件下表现出不同的特点,在三点中赣北的树高、枝下高、冠幅和树冠表面积最大。赣中的冠长表现最优,而赣南则胸径最大,土壤和区域气候条件对生长及树冠性状均有一定影响,关于该结论已有相关报道[25-26]。
林木生长性状与其产脂性状关系较为复杂,既可能为正相关又可能为负相关关系。有研究认为,胸径大的林木产脂量高,主要是因为胸径大的植株其树脂道数量较多,而树脂道数量又与产脂量呈正相关关系[1-2,21]。本研究中,通过产脂量与生长性状的遗传相关关系可知,胸径与产脂量的遗传相关系数高达0.93,是影响松脂产量最重要的因子之一,与上述结论基本一致。然而,另有研究表明,针叶树的生长性状与其产脂能力(单位胸径的产脂量)呈负向相关[27-29]。生长/分化平衡假说(growth/differentiation balance hypothesis,GDBH)对此的解释为,在环境胁迫条件下,植物体内的碳水化合物优先保障具有防御功能的萜烯、酚酸和松脂等次生化合物的合成,而植物体内的碳水化合物是相对有限的,当碳水化合物优先分配给次生代谢物时,其分配给植物生长需求的碳水化合物则会相应减少,从这点来看,生长性状与其产脂力为负相关关系[24,30]。Hood 等[3]对美国黄松(P.ponderosa)的研究认为生长性状与产脂性状的相互关系取决于所选树脂道性状的度量单位。例如,生长较慢的美国黄松在单位面积内树脂道数目和单位面积内树脂道总面积要高于生长较快的林木,当以此为度量单位时,生长与产脂性状为负相关关系;然而,生长较快的美国黄松在单个树脂道平均面积、树脂道总数目和树脂道总面积要高于生长较慢的林木,而当以此为度量单位时,生长与产脂性状为正相关关系。产脂量与树冠性状的遗传相关分析结果表明,树冠表面积与产脂量的遗传相关系数为0.83,是影响松脂产量的另一重要因子。从生理角度来看,松脂是光合作用的次生代谢产物,而植物体内次生代谢物质积累与光合作用密切相关,说明光合能力是决定松脂产量高低的重要因素[30-32]。针叶作为松树的光合器官,林木树冠表面积越大,能够进行光合作用的针叶就越多,光合能力就越强,其相应的产脂量也越高[1,33]。
和遗传相关相比,遗传通径分析能把性状间的遗传相关性分解成直接和间接作用,可更深入评价因变量对自变量具体的控制程度和控制机理,同时利用各性状的决定系数总贡献可表现影响程度,因此其在林木选育过程中具有十分重要的作用[34-35]。对产脂量与生长及树冠性状进行通径分析发现,除胸径和树冠表面积外,树高和冠幅也是对产脂量影响较大的因子。有学者认为,松树在进行采脂时,随着割面面积增加、树脂道破坏增多,出脂量也会随之增大,在其他因素一致的情况下,树越高,树木营养健康状况越好,生长越旺盛,作为新陈代谢产物的松脂产量也越大[36]。许建伟[5]在研究马尾松树体因子与产脂量的相关关系时发现,冠长对产脂量的影响作用强于冠幅,与本研究遗传通径分析结论不一致。根据遗传相关和通径分析结果,综合得出对产脂量影响最大的4 个因子分别为胸径、树高、树冠表面积和冠幅,其与产脂量构建的多元回归方程为y=-0.255 +0.040x(DBH)+0.004x(HGT)+0.006x(CSA)+0.036x(CW),此模型预估精度达99.64%。在高产脂选育过程中,通过树高、胸径和树冠表面积等较易测得的因子,应用上述多元回归模型可对产脂量进行初步预估;根据产脂量预估结果结合割脂精确称量开展高产脂选育工作,可大大降低误选率并提高选择效率,节约人力和物力。
4 结论
湿地松产脂量与主要生长及树冠性状(除枝下高外)呈显著或极显著的遗传正相关关系。遗传通径分析中,本研究选取的6 个生长及树冠因子可以解释产脂量影响作用的82%,其中,胸径、树高、树冠表面积和冠幅成为影响产脂量的主导因素,其他影响林木产脂量的因子有待进一步研究。建立的产脂量与胸径、树高、树冠表面积和冠幅的多元回归模型,预估精度为99.64%,能较好地预测林木的产脂量。在湿地松高产脂遗传改良过程中,通过对树高、胸径、树冠表面积和冠幅的快速测定,可达到预测产脂量的目的,大大减少工作量,以实现早期选育。
猜你喜欢
东北林业大学学报(2023年6期)2023-05-31 02:56:16
丽水学院学报(2022年2期)2022-04-19 01:22:10
农业与技术(2021年6期)2021-12-12 05:33:47
黑龙江气象(2021年2期)2021-11-05 07:06:58
林业科技情报(2021年3期)2021-09-01 02:01:26
机械工程与自动化(2021年4期)2021-07-30 09:37:02
机械工程与自动化(2020年6期)2020-12-28 11:58:26
林业与生态(2020年2期)2020-04-16 12:48:31
中南林业调查规划(2017年1期)2017-12-19 00:58:07
广西林业科学(2016年3期)2016-03-16 05:43:30
