
山西太行黑山羊mtDNA D-Loop区遗传多样性分析
2020-06-19 04:49闫益波杜丽英张伯池宋献艺程俐芬李文刚曹宁贤
山西农业科学 2020年6期
闫益波,杜丽英,张伯池,张 凯,张 菁,宋献艺,程俐芬,李文刚,曹宁贤
(1.山西省农业科学院畜牧兽医研究所,山西太原030032;2.山西省农业科学院饲料兽药研究所,山西太原030031;3.山西省畜禽繁育工作站,山西太原030001)
线粒体DNA(Mitochondrial DNA,mtDNA)是动物细胞核外唯一的遗传物质,具有基因结构简单、稳定,碱基突变率高,遗传上具有自主性及遵循严格的母系遗传等特点,为系统发育和遗传结构研究的良好素材[1]。其中,片段长1 220 bp的mtDNA D-环控制区(D-Loop)是mtDNA基因组中最主要的一段非编码区,控制mtDNA的复制和转录,受到进化的压力较小,进化速率最高,最具多态区域,尤其在高变区;由于D-Loop受自然选择影响较大,存在巨大变异,所以广泛应用于哺乳动物群体遗传多样性、物种亲缘关系、种群间系统进化分析和分子系统发生等方面的研究[2-3]。山西太行黑山羊(左权黑山羊)是一个重要的地方品种,分布广而分散、体型外貌多样、生产用途不突出,受各种因素的影响,目前群体数量急剧减少。合理保护和开发山西太行黑山羊品种资源是山西省畜牧产业发展的重要任务。
本研究在前期选育研究的基础上[4],对其mtDNA D-Loop进行测序分析,从系统发生方面研究山西太行黑山羊的遗传进化,旨在为山西太行黑山羊遗传资源保护、开发和杂交利用提供理论依据。
1 材料和方法
1.1 样品采集
试验样品采自左权县新世纪农业科技有限责任公司黑山羊种羊场核心选育群,随机选取健康成年黑山羊35只(检查系谱记录,避免选择亲缘关系较近的个体),采集其耳组织样品于冻存管中,冷藏箱保存带回实验室,-80℃冰箱保存。
1.2 基因组DNA的提取、扩增和测序
采用AXYGEN动植物基因组DNA小量制备试剂盒(AP-MN-MS-GDNA-250)进行总DNA提取。由于山羊的D环区位于细胞色素B基因(CYTB)和12SrRNA基因中间(图1),所以需要在CYTB基因和12SrRNA基因的保守区设计引物(引物序列为F:TTGATACCTGCTCCTCTTAG和R:CAGTCG AACAYCCCTAYRTT)扩 增mtDNA D-Loop,片段大小约1 500 bp。
PCR扩增使用的是Vazyme Phanta Max Super-Fidelity DNA Polymerase扩增。首先对样品稀释到20 ng/μL,再进行PCR扩增。轻弹混匀,瞬时离心收集管壁上的液滴至管底,在PCR扩增仪上进行PCR反应。PCR反应总体系为50μL,包括基因组DNA(20 ng/μL)1μL,2×Phanta Max Buffer 25μL,Phanta Max Super-Fidelity DNA Polymerase(1 U/μL)1μL,dNTPMix(10mmol/L)1μL,正向引物(10μmol/L)2μL、反向引物(10μmol/L)2μL,ddH2O 18μL。PCR反应条件为:95℃预变性3 min;95℃变性15 s,52.3℃退火15 s,72℃延伸45 s,35个循环;72℃延伸5 min,4℃保存。反应完成后,取3μL PCR产物进行1%琼脂糖凝胶电泳检测,以确认PCR扩增片段。
PCR产物用AxyPrep DNA凝胶回收试剂盒回收,具体操作按试剂盒说明书进行。在得到适合测序的PCR产物后,首先对其进行直接测序。测序PCR反应选用的试剂为ABI公司的BDT3.1测序试剂盒(BigDyeTerminator v3.1),测序反应均按照BDT 3.1使用手册进行。测序PCR产物的纯化采用乙醇沉淀方法,干燥后4℃避光保存,加入10μL甲酰胺,使纯化后的干粉充分溶解,随后置于PCR仪中进行变性反应,将变性后的样品采用3730XL测序仪进行测序。
1.3 测序数据分析
测序结果经DNAMAN比对、拼接和校正后,得到完整的序列。格式转换(Bioedit软件)后,用Clustalx 1.83软件进行序列同源性比对,以MEGA文件格式保存。用分子进化遗传分析软件MEGA 7.0确定单倍型、多态位点,构建Neighbor Joining(NJ)系统发育树。再利用DNASP4.0软件对核苷酸多样度(Pi)、单倍型多样度(Hd)以及平均核苷酸差异数等进行统计分析。
2 结果与分析
2.1 PCR扩增结果
对35只山西太行黑山羊mtDNA D-Loop区序列进行PCR扩增,经1%琼脂糖凝胶电泳检测,可见PCR扩增产物条带明亮清晰,无拖尾(图2)。扩增获得大小约1 200 bp的片段,与试验设计扩增片段长度大小相符,说明扩增效果良好。
2.2 山西太行黑山羊与其他山羊品种mtDNA DLoop区的核苷酸组成分析
对35只山西太行黑山羊个体D-Loop区序列碱基组成情况进行分析,结果表明(表1),山西太行黑山羊的D-Loop区序列长度为1211~1215bp,其中,1 211 bp 1只、1 212 bp 25只、1 213 bp 6只、1 214 bp 2只,1 215 bp 1只,T、C、A、G这4种碱基平均占比分别为28.44%、26.22%、31.82%、13.52%,C+G的占比为39.74%(表2)。说明山西太行黑山羊mtDNAD-Loop区富含A/T碱基,具有一定的偏倚性。20个黑山羊群体中,大部分个体的mtDNA D-Loop区片段长度分布在1 212~1 213 bp范围内(表3)。

表2 山西太行黑山羊mtDNA D-Loop序列的碱基组成情况

表3 山西太行黑山羊与其他山羊品种mtDNA D-Loop序列的碱基组成比较
2.3 山西太行黑山羊mtDNA D-Loop区序列多样性分析
采用DNAsp 5.0软件对35条序列进行分析,结果表明,山西太行黑山羊共有34种单倍型(表4),单倍型多样度(Hd)为0.998,平均核苷酸多样度(Pi)为0.018 65,平均核苷酸差异度(k)为22.0;其余18个山羊品种258只个体中共发现单倍型210种。对19个山羊品种的mtDNA D-Loop区全序列进行Tajima中性显著性检验可知,其值变化范围为-1.297 52~1.787 86,均不显著,符合中性突变(表5)。
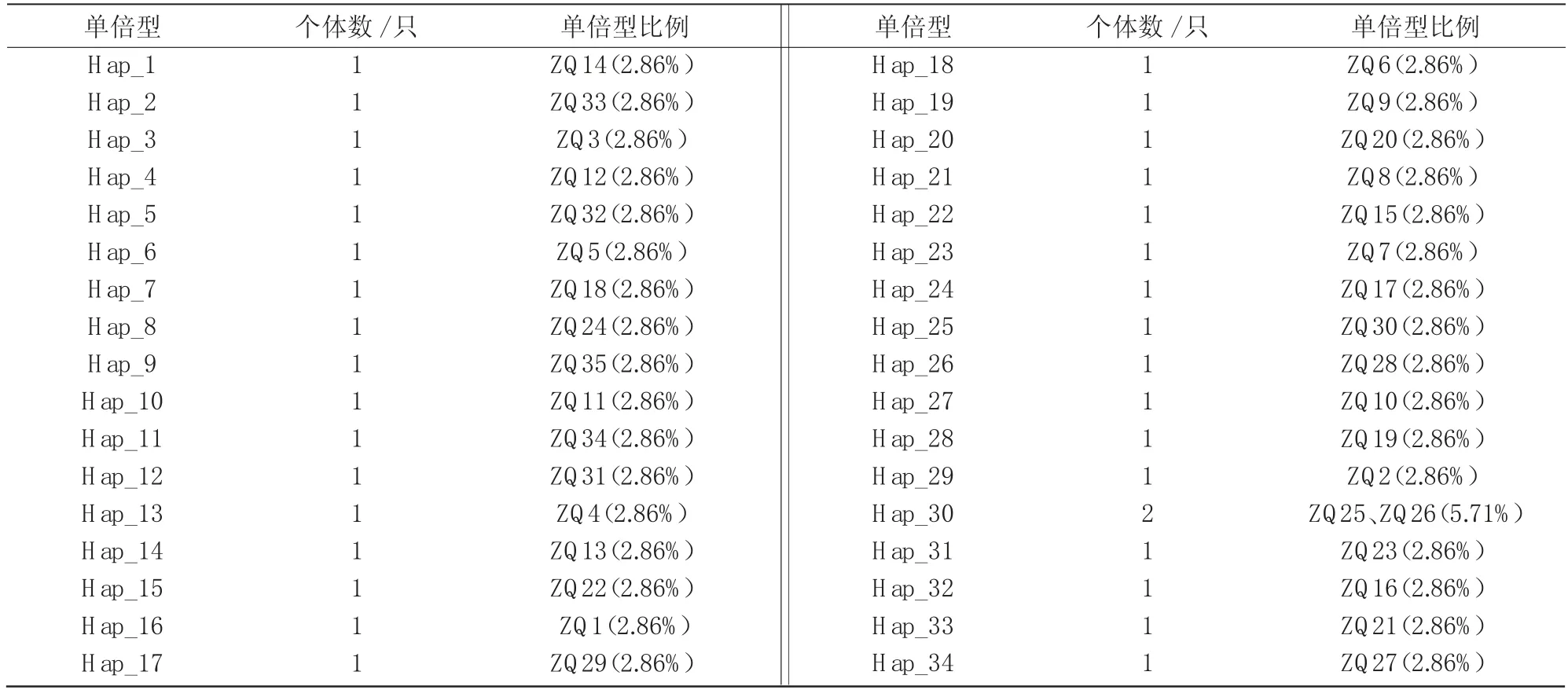
表4 山西太行黑山羊mtDNA D-Loop序列的单倍型分布情况
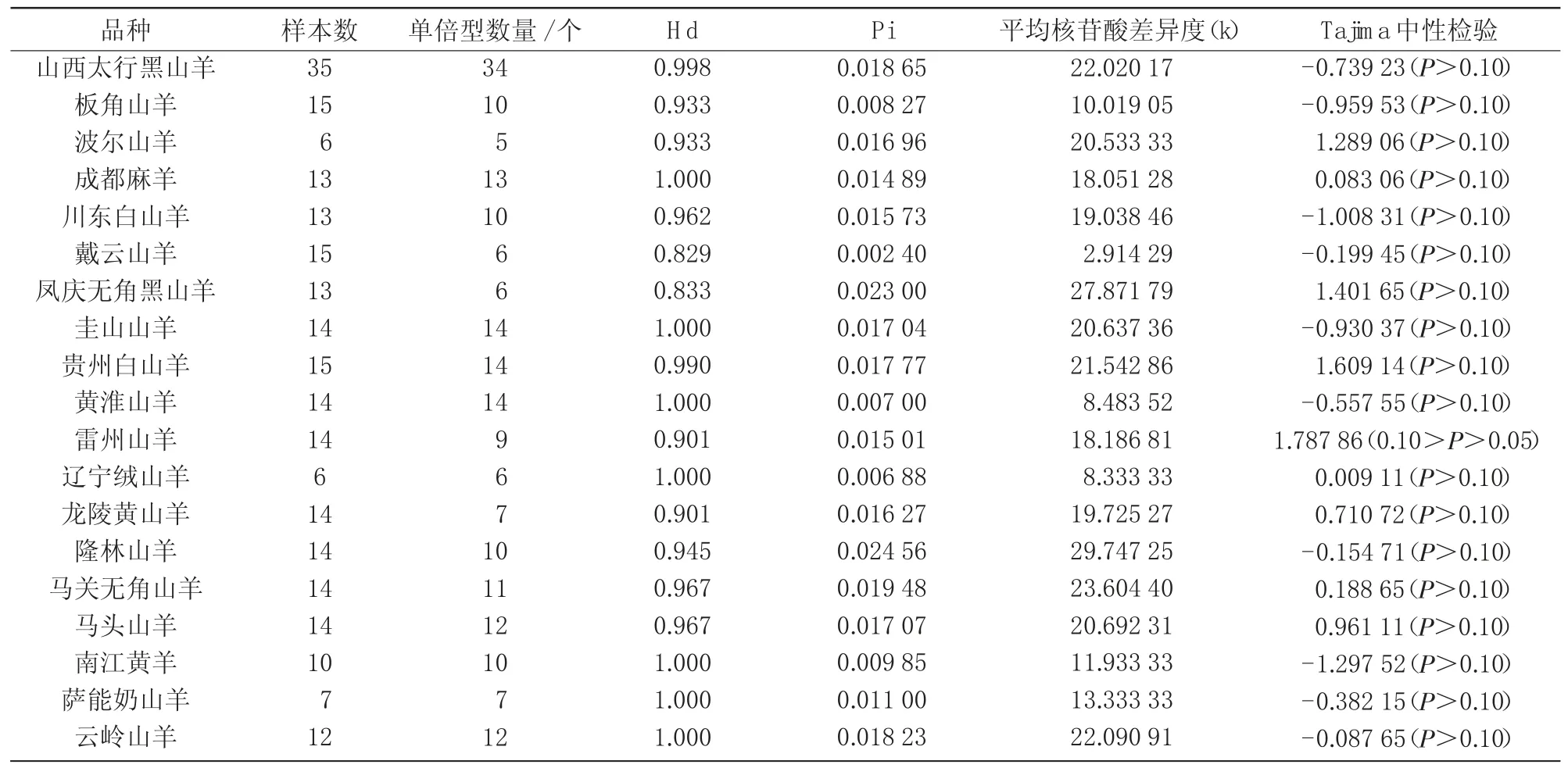
表5 山西太行黑山羊与其他山羊品种mtDNA D-loop序列遗传多样性
2.4 山西太行黑山羊与中外部分山羊品种的遗传距离分析
遗传距离是衡量种群间遗传分化程度和差异大小的重要指标。本研究中,对20个山羊品种260条序列利用MEGA 4.0软件分析得到20个群体品种间遗传距离(表6),结果表明,20个山羊群体品种间的遗传距离为0.003~0.033;辽宁绒山羊与凤庆无角山羊之间的遗传距离较小,为0.003;凤庆无角山羊与安哥拉山羊的遗传距离最远,为0.033。与山西太行黑山羊遗传距离由小到大的品种分别为辽宁绒山羊、黄淮山羊、戴云山羊、萨能奶山羊、成都麻羊、圭山山羊、板角山羊、马头山羊、川东白山羊、马关无角山羊、龙陵黄山羊、云岭山羊、贵州白山羊、雷州山羊、南江黄山羊、波尔山羊、凤庆无角黑山羊、隆林山羊、安哥拉山羊。山西太行黑山羊与辽宁绒山羊、黄淮山羊和戴云山羊的遗传距离最小,为0.014,表明其亲缘关系较近;山西太行黑山羊与安哥拉山羊的遗传距离最大,为0.029,表明二者亲缘关系较远。

LNC MG ZQ YL SA NJ MT离LL距传Nei's遗LLY的间LZ种品羊HH山他其外GZW中与羊山GS黑行太FQ西山6 DY表CDW CDM BOR BJ AN 0.029种 AN BJ品0.019 0.032 BOR 0.020 0.017 0.028 CDM 0.022 0.020 0.013 0.011 0.025 0.018 0.019 0.014 0.020 0.016 0.023 0.019 0.023 0.021 0.016 0.010 0.022 0.017 0.030 0.025 0.033 0.028 CDW DY FQ GS 0.017 0.020 0.014 0.021 0.019 0.018 0.017 0.016 0.017 0.011 0.020 0.021 0.021 0.021 0.018 0.020 0.018 0.021 0.020 0.020 0.012 0.024 0.024 0.021 0.021 0.024 0.022 0.021 0.022 0.021 0.022 0.017 0.018 0.021 0.020 0.017 0.022 0.025 0.018 0.020 0.021 0.022 0.021 0.021 0.014 0.018 0.017 0.021 0.015 0.019 0.010 0.017 0.014 0.017 0.006 0.017 0.017 0.022 0.024 0.017 0.019 0.020 0.021 0.021 0.021 0.015 0.020 0.013 0.020 0.019 0.022 0.018 0.021 0.015 0.019 0.018 0.019 0.012 0.022 0.021 0.023 0.021 0.025 0.023 0.022 0.021 0.021 0.023 0.022 0.021 0.014 0.007 0.014 0.017 0.020 0.013 0.017 0.011 0.017 0.014 0.017 0.003 0.019 0.014 0.019 0.022 0.025 0.018 0.022 0.018 0.022 0.020 0.022 0.010 0.020 0.013 0.020 0.017 0.021 0.019 0.019 0.014 0.017 0.018 0.018 0.013 0.019 0.020 0.019 0.021 0.024 0.020 0.019 0.021 0.022 0.022 0.021 0.020 0.017 0.013 0.016 0.020 0.023 0.016 0.019 0.017 0.020 0.019 0.020 0.010 0.030 0.025 0.031 0.030 0.033 0.029 0.031 0.027 0.031 0.029 0.030 0.025 GZW HH LZ LLY LL MT NJ SA YL ZQ MG LNC无庆;FQ.凤羊山白东;CDW.川羊山角无关;MG.马羊山山;GS.圭羊羊。山山头绒;MT.马宁山;LNC.辽羊羊白山州;GZW.贵云;DY.戴羊羊山山岭奶;YL.云能羊;SA.萨山羊黄山江淮;NJ.南;HH.黄羊羊山山黑角权;ZQ.左;BJ.板羊羊山山州林;LL.隆;LZ.雷羊羊山麻尔都;CDM.成;BOR.波羊羊山山黄拉陵哥:AN.安;LLY.龙羊注山黑角
2.5 山西太行黑山羊系统进化分析
根据遗传距离,采用邻接法(NJ)构建山西太行黑山羊与其他19个山羊品种间的系统发生树,以便直观地了解20个种群间的亲缘关系,结果显示(图3),20个种群被分为两大支,安哥拉山羊独自形成一支;以我国的地方山羊为主的19个品种属于一个大支,其又分为2个亚支:龙陵黄山羊与马关无角山羊首先聚在一起,然后与贵州白山羊和南江黄山羊聚在一起形成其中一个亚支;山西太行黑山羊与其他14个品种聚为一个亚支,但山西太行黑山羊单独聚为一支。
2.6 mtDNA D-Loop区系统发生树分析
利用NETWORK软件对260只个体绘制了网络关系分析图(图4)。从图4可以看出,20个山羊群体分为A、B、C、D共4个不同的支系,且每个支系呈现“星状”,分布清晰,没有交叉。
3 结论与讨论
左权县现存的山西太行黑山羊是在当地特殊地理及生态环境条件下,经过长期的人工选择形成的地方类群。一般认为,该类群从分布上属于太行山羊的范畴,但太行山区纬度跨度大,羊群分布广,从而形成了特点鲜明的多种类型,比如从毛色可分为黑、褐、青、灰、白等类型,从生产类型可分为偏绒用和偏肉用2种。已有研究表明,山西太行黑山羊具有抗逆性和适应性强的优点,尤以产肉性能著称于国内市场;生产的羊肉具有脂肪分布均匀、肉质细嫩、味道鲜美、几乎没有难以接受的膻腥味、口感极好,而且蛋白质含量高、脂肪及胆固醇含量低、滋补作用强。由于市场对黑山羊的需求逐年增加,加之封山禁牧政策的影响和长期缺乏系统科学的选育,从而导致该品种群体规模急剧下降,其珍贵的遗传多样性和独特的遗传特性正受到破坏和影响,急需对其进行保护、选育及科学利用。
mtDNA D-Loop区多态性常被作为群体内、群体间遗传多样性分析方法,本研究通过对山西太行黑山羊线粒体D-Loop序列与GenBank中收录的部分品种山羊线粒体D-Loop区序列进行比对分析,从系统发生方面研究山西太行黑山羊的遗传进化,旨在为山西太行黑山羊遗传资源保护与开发利用提供理论依据。
本研究通过对35只山西太行黑山羊个体mtDNA D-Loop区进行测序及与其他19个中外山羊品种的mtDNA D-Loop区全序列进行比较分析,并对其遗传多样性及起源进化情况进行深入分析,结果显示,山西太行黑山羊mtDNA D-Loop区全序列中,A、T、C、G的含量分别为31.82%、28.44%、26.22%和13.52%,碱基组成分析显示,G相对缺乏,C+G的含量为39.74%,A+T的含量为60.26%,A+T的含量明显比G+C的含量多,说明山西太行黑山羊mtDNA D-Loop区富含A/T碱基,具有一定偏倚性。这与王杰等[5]对我国7个山羊品种mtDNA D-Loop区碱基组成的研究结果较为相似。可能是由于CG含量越丰富序列越趋于稳定,而mtDNA D-Loop变异率较高,所以富含A/T碱基[6]。20个黑山羊群体中,大部分个体的mtDNA D-Loop区片段长度分布在1 212~1 213 bp范围内,与已有研究报道中的1 212~1 213 bp范围趋于一致[7]。另外,本研究在35只山西太行黑山羊中共检测到34种单倍型,单倍型多样性及核苷酸多样性均较高,也证明了山西太行黑山羊具有较高的遗传变异程度,遗传多样性丰富,这与王杰等[5]的研究结果基本一致。武建亮等[8]利用PCR-SSCP技术对8个山羊品种的分析结果显示,单倍型多样度以太行黑山羊最高,为0.800 3,与本研究的结果(0.998)接近。山西太行黑山羊与我国众多的地方山羊品种一样,都具有一定的选择潜力,遗传多样性丰富。
一般认为,群体中mtDNA核苷酸多样度(Pi)和单倍型多样度(Hd)是衡量一个群体遗传分化的重要指标,Pi值和Hd值越大,群体的多态程度越高,遗传多样性越丰富,大多数哺乳动物的Pi值均大于0.01,0.001~0.007的Pi值范围被认为是缺乏种内变异的物种指标范围[9]。本研究中,山西太行黑山羊Pi和Hd值分别为0.018 65和0.998,显示出较高的群体多态性,表明山西太行黑山羊群体遗传多样性丰富。
从山西太行黑山羊与其他19个山羊品种间的遗传距离来看,山西太行黑山羊与辽宁绒山羊、黄淮山羊的遗传距离最近,为0.014;与凤庆无角黑山羊和隆林山羊相对较远,遗传距离分别为0.023和0.024;与安哥拉山羊的遗传距离最远,为0.029,表明山西太行黑山羊在起源进化的过程中更多受到东北和华东山羊的影响,而这些地区冬季寒冷的气候对毛皮性状具有较大的选种要求,山西太行黑山羊与辽宁绒山羊、黄淮山羊都以产绒性状和板皮性状为优,因而群体间起源较近,群体间分化主要由地理隔离造成,分化程度较低。
《中国羊品种志》认为,我国家养山羊有2个野生祖先,一个是角骨羊(Capra aegagrus),另外一个为旋角野山羊(Capra falconeri)。张文丽等[10]研究发现,会理黑山羊与太行黑山羊、成都麻羊、贵州白山羊等我国地方品种山羊一起和角骨羊聚为一大分支,而旋角野山羊则单独聚为一个分支,角骨羊与我国山羊亲缘关系较近,对我国山羊贡献较大。本研究显示,山西太行黑山羊与辽宁绒山羊、黄淮山羊、戴云山羊、萨能奶山羊、成都麻羊、圭山山羊等多个地方品种存在亲缘关系,与上述研究结果有部分相似,提示山西太行黑山羊的母系起源为角骨羊,虽然生活在北方,但与南方山羊品种具有一定渊源,可能在长期进化中,由于饲养管理粗放、选育程度较低,受到了不同品种山羊的影响。
利用mtDNA D-Loop区单倍型构建群体系统发育树是研究种群遗传进化关系的常用方法[11]。本研究中,20个山羊品种系统发生树显示,其共分为4个支系,且每个支系呈现“星状”,与NJ树的分析结果一致,暗示我国山羊至少存在有4个独立的母系起源,“星状”分布清晰,无交叉,可进一步说明4个支系间没有复杂的遗传关系,表明我国地方山羊品种至少有4个母系起源。刘益平等[12]研究认为,类型A和B是在我国山羊品种中广泛存在的类型,而类型C和D则以低频率存在于我国部分山羊个体中,至于是否还有A、B、C、D类型以外的其他新的线粒体D环类型,各类型在我国山羊品种中的分布频率如何,仍需要测定更多的其他地方山羊品种的材料来检验和分析,这与本研究结论基本一致。
总体上,山西太行黑山羊群体的多态程度高,遗传多样性丰富,选育程度较低,应当注意加以保护,防止盲目杂交,避免基因库的混杂和我国地方品种某些优良基因丢失,以保持我国山羊品种丰富的遗传多样性。
猜你喜欢
中国人兽共患病学报(2022年9期)2022-10-19
云南畜牧兽医(2022年4期)2022-08-19
云南畜牧兽医(2021年6期)2021-12-24
今日农业(2021年15期)2021-11-26
云南畜牧兽医(2021年5期)2021-10-27
科学导报(2021年29期)2021-06-03
中国生殖健康(2020年4期)2021-01-18
科海故事博览·下旬刊(2019年6期)2019-04-16
作文周刊·小学一年级版(2017年46期)2018-01-16
小天使·一年级语数英综合(2017年4期)2017-04-18
