
亚麻荠CsDGAT3基因的鉴定与表达分析
2020-06-19 04:49高慧玲张宏江刘宝玲薛金爱苑丽霞李润植
山西农业科学 2020年6期
高慧玲,张 飞,张宏江,高 宇,刘宝玲,薛金爱,苑丽霞,李润植
(1.山西农业大学分子农业与生物能源研究所,山西太谷030801;2.晋中学院生物科学与技术学院,山西晋中030600)
亚麻荠(Camelina sativa(L.)Crantz)是十字花科植物中一种古老的油料作物。与其他十字花科植物相比,亚麻荠是一种低投入、易栽培的环保型经济作物,而且其具有需肥量低、耐旱性强以及抗斑点落叶病和其他病虫害能力强等特性,符合现代农业的可持续发展要求[1-4]。亚麻荠种子中含油量占干质量的37%~45%,其中,大多数为多不饱和脂肪酸,富含亚麻酸和亚油酸。目前,亚麻荠油已成为国际上推崇的最佳食用油[5-6];同时,亚麻荠也被应用于食品、饲料和工业等多个领域,且在生物燃料和植物油的生产上也具有巨大的农艺潜力[7]。随着亚麻荠油市场需求的大幅增长,通过基因工程和基因组编辑方法提高亚麻荠油产量和质量等成为研究热点[8],深入了解油脂的生物合成及其生物过程的内在分子机制将为亚麻荠的遗传改良和高价值产品的可持续商业化奠定基础。
植物种子油主要为三酰甘油(Triacylglycerol,TAG),作为能量储存脂质,TAG在植物种子萌发和幼苗形成过程中提供能量和碳来源[9];此外,TAG也是食品、饲料、燃料和其他油脂化工中碳分子的主要来源。大量报道表明,二酰甘油酰基转移酶(Diacylglycerol acyltransferase,DGAT)是参与TAG生物合成的最后一步酰化反应的限速酶[10-13]。目前,已发现DGAT酶的4种类型:DGAT1、DGAT2、DGAT3和WS/DGAT。其中,DGAT1和DGAT2已经在大多数真核生物中得到广泛研究。一般认为,植物DGAT1在种子油脂的合成与积累中起主要作用[14];而DGAT2更侧重于积累特殊脂肪酸,如在油桐[15](Vernicia fordii)、蓖麻[16](Ricinus communis)和斑鸠菊[10](Vernonia anthelmintica)中发现DGAT2可以促进特殊脂肪酸的合成,但是二者的作用并不相互排斥。而WS/DGAT和DGAT3是分别从拟南芥[17]和花生[18]中最新鉴定出来的,在植物中的研究还相对比较少。拟南芥WS/DGAT虽可促进少量TAG的合成,但其主要催化蜡酯的合成[17]。CsDGAT3蛋白无跨膜区,为可溶性酶蛋白,最初从发育中的花生(Arachis hypogaea)子叶中分离并验证[18]。目前,在拟南芥、大豆和亚麻荠等油料作物中发现了DGAT3的同源序列[11,18-20],但仍未对其功能进行验证。
本研究采用生物信息学工具,从亚麻荠全基因组数据库中筛选、鉴定亚麻荠CsDGAT蛋白,并对其进行生物信息学分析,检测亚麻荠CsDGAT3基因的表达谱,以期深入了解植物DGAT3功能的多样性以及为其在生物技术的进一步应用提供新的思路。
1 材料和方法
1.1 材料
本研究所用的亚麻荠由山西农业大学分子农业与生物能源研究所种植、储藏,所选的材料包括亚麻荠的根、茎、叶、花及种子的不同时期(早期种子(开花后的4~20 d)、中期种子(开花后的22~30 d)和晚期种子(开花后的32~40 d))。
1.2 CsDGAT3基因的鉴定
由于DGAT3基因的功能已在花生中得到验证,以花生AhDGAT3(AAX62735.1)蛋白序列为检索序列,在亚麻荠基因组数据库(www.ncbi.nlm.nih.gov/genome/?term=camelina+sativa)下载获得亚麻荠基因组gDNA、CDS和蛋白序列,随后在亚麻荠转录组中筛选并获得目的基因。
1.3 亚麻荠CsDGAT蛋白的基本结构分析
利用在线ExPasy网站中ProtParam工具(http://web.expasy.org/protparam/)分析亚麻荠CsDGAT3的基本理化性质;使用在线Target P 1.1 Server软件(http://www.cbs.dtu.dk/services/TargetP/)预测亚麻荠CsDGAT3蛋白的亚细胞定位;依据TMHMMServer v.2.0(http://www.cbs.dtu.dk/services/TMHMM/)分析CsDGAT3蛋白的跨膜区;采用保守结构域数据库(Conserved Domain Database,CDD)鉴定CsDGAT3蛋白的保守域;采用SWISS-MODEL(https://swissmodel.expasy.org/interactive)分析亚麻荠CsDGAT3蛋白的三级结构。
1.4 亚麻荠CsDGAT3蛋白的多序列比对和系统发育分析
使用MEGA 6.06软件对亚麻荠CsDGAT3和花生AhDGAT3蛋白的氨基酸序列进行Clustal W多序列比对;采用邻位连接法(NJ)对DGAT3蛋白(表1)构建系统进化树。
1.5 亚麻荠CsDGAT3的表达检测
根据鉴定结果,从转录组数据中获得了3个CDS序列,分别为CsDGAT3-1、CsDGAT3-2和CsDGAT3-3,并将其与亚麻荠基因组序列进行了比较。
qTR-PCR引物采用Primer 6.0设计,内参基因Actin引物和定量引物如表2所示。使用RNA提取试剂盒(TaKaRa)提取RNA。使用cDNA Synthesis Kit反转录试剂盒(Genstar公司)合成cDNA。使用2×增强型染料实时荧光定量PCR预混液(GenStar公司)进行qRT-PCR,以Actin作为内参;然后用2-ΔΔCt法进行基因表达分析。每个样品至少进行3次生物学重复。
2 结果与分析
2.1 亚麻荠CsDGAT3基因的鉴定
为了验证CsDGAT3基因在亚麻荠TAG生物合成中的作用,以已经鉴定出的花生AhDGAT3氨基酸序列为探针,在亚麻荠基因组数据库中筛选出3个具有完整开放阅读框(ORF)候选CsDGAT3基因,即CsDGAT3-1、CsDGAT3-2和CsDGAT3-3,且CDS序列都含有TRX-like Fd家族的典型结构域。现将其重新命名(表3),方便后续进行分析。
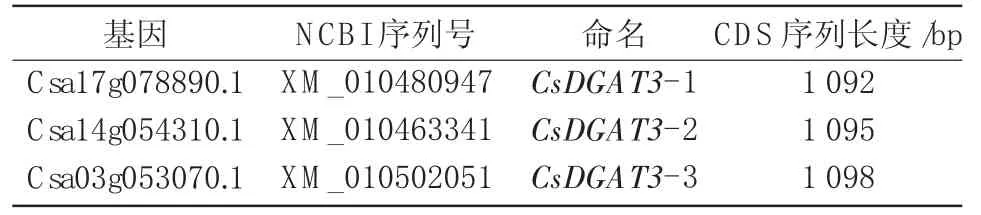
表3 亚麻荠CsDGAT3基因的序列信息
2.2 亚麻荠CsDGAT3蛋白的理化性质
从表4可以看出,CsDGAT3-1、CsDGAT3-2和CsDGAT3-3基因编码的蛋白质长度分别为361、364、365个氨基酸,其理化性质与花生AhDGAT3相似;脂肪酸指数和亲水性系数分析表明,Cs-DGAT3是水溶性蛋白质;跨膜区分析进一步显示,均无跨膜结构;亚细胞定位预测表明,CsDGAT3蛋白最有可能定位在细胞质中。

表4 亚麻荠CsDGAT3和花生AhDGAT3蛋白质理化性质分析
2.3 亚麻荠CsDGAT3蛋白的高级结构分析
通过保守结构域数据库(CDD)对CsDGAT3蛋白的保守功能域分析表明,CsDGAT3-1、CsDGAT3-2和CsDGAT3-3蛋白均含有TRX-like Fd家族的典型结构域,这类TRX-like-Fd结构域含有4个参与[2Fe-2S]簇结合的保守半胱氨酸,在DGAT3家族成员中检测到该结构域是保守的。以ID1m2b.1为模板对CsDGAT3蛋白进行三级结构分析,结果显示(图1),CsDGAT3-1、CsDGAT3-2和CsDGAT3-3与模板(ID1m2b.1)的相似度分别为25.00%、25.00%和23.68%,且均由2个亚基组成,并且均匀对称。
2.4 CsDGAT3蛋白氨基酸多序列比对及系统发育分析
使用MEGA 6.06软件对亚麻荠CsDGAT3和花生AhDGAT3蛋白的氨基酸序列进行Clustal W多序列比对,结果表明,CsDGAT3-1、CsDGAT3-2、CsDGAT3-3和花生AhDGAT3的序列相似性均为35%,CsDGAT3-1与CsDGAT3-2、CsDGAT3-3序列相似性分别为95%和93%;3个CsDGAT3与花生AhDGAT3-1的氨基酸序列的进一步分析结果如图2所示,共鉴定出磷酸化位点(Ⅰ区)、潜在的DGAT基序(Ⅱ区)以及假定的硫解酶酰基酶(Ⅲ区)和酪氨酸激酶磷酸化保守位点(Ⅳ区)几个保守基序。亚麻荠CsDGAT3-1、CsDGAT3-2和CsDGAT3-3所存在的多个功能位点可能预示着其活性受其他因素的调控或相互作用。
采用邻接法(NJ)对各物种DGAT3蛋白构建系统发育树,结果如图3所示,亚麻荠3个CsDGAT3蛋白均聚在一起,且与荠菜CrDGAT3最接近,并且十字花科家族的DGAT3组成了一个紧密的群体,包括拟南芥、荠菜、山萮菜属、油菜,表明亚麻荠CsDGAT3可能与其他十字花科植物具有共同的进化祖先。进一步说明,从亚麻荠中鉴定出的CsDGAT3基因可能编码TAG的合成。
2.5 亚麻荠CsDGAT3基因的表达谱分析
为了探究CsDGAT3基因在亚麻荠种子油脂积累中的作用,对亚麻荠CsDGAT3在根、茎、叶、花以及种子发育的3个阶段(早、中、晚)和萌发种子(种子萌发5 d的幼苗)中的表达谱进行检测,结果如图4所示,CsDGAT3-1、CsDGAT3-2和CsDGAT3-3基因在亚麻荠各个组织中均有表达,但略有差异,CsDGAT3-1在根中的表达高于CsDGAT3-2和Cs-DGAT3-3;而CsDGAT3-2在花和萌发种子中的表达高于CsDGAT3-1和CsDGAT3-3。进一步推测,萌发种子中CsDGAT3-2的高表达水平可能与拟南芥AtDGAT3基因一样。AtDGAT3在种子发芽过程中利用已合成的TAG来提供能量。CsDGAT3-3在发育中的种子中表达水平最高,进一步推测,CsDGAT3-3可能在种子油的生物合成中起重要作用。
3 结论与讨论
种子油的含油量和脂肪酸组成是大豆、芸薹属和亚麻荠属等油料作物最重要的2个农艺性状,为了有效地提高这2个性状的遗传改良效果,迫切需要确定影响种子油生物合成和积累的关键基因。越来越多的研究表明,DGAT在真核生物TAG生物合成的最后一步和限速步骤中起着至关重要的作用[7,21-23]。与研究充分的DGAT1和DGAT2基因家族成员相比,DGAT3基因仅在几种植物中被鉴定,但其功能尚未验证[11,18-20]。
本研究通过对比花生AhDGAT3序列,从亚麻荠数据库中鉴定获得3个亚麻荠DGAT3蛋白,分别命名为CsDGAT3-1、CsDGAT3-2和CsDGAT3-3。这与亚麻荠基因组是高度未分化的六倍体基因组相吻合[24-25]。据文献报道,不管是单倍体的绿藻如团藻(Volvox carteri)和绿色鞭毛藻(Ostreococcus lucimarinus)还是主要的二倍体商业作物如大豆、玉米和水稻,其基因组中都存在1个或2个DGAT3基因[26]。DGAT3家族中不同成员的存在可为行使不同的功能提供可能。
理化性质分析表明,CsDGAT3-1、CsDGAT3-2和CsDGAT3-3蛋白均定位于细胞质中,为水溶性蛋白。最早在花生中鉴定出能在细胞质中合成TAG的可溶性DGAT酶[18],而且拟南芥AtDGAT3蛋白和花生的3种AhDGAT3蛋白均被鉴定为水溶性且定位于细胞质中,这与亚麻荠CsDGAT3蛋白的亚细胞定位相吻合。此特性使得在基因工程中利用DGAT3来提高油料作物的产油量更加便利。
本研究系统发育分析表明,亚麻荠DGAT3蛋白与荠菜CrDGAT3蛋白具有高度同源性,这可能与2个物种同属于十字花科且遗传封闭有关。多序列比对结果显示,3种亚麻荠CsDGAT3蛋白均具有典型的DGAT保守区和一个酰基转移酶结合位点;此外,还鉴定出了其他的几个保守的基序,包括磷酸化位点、假定的硫解酶酰基酶和酪氨酸激酶磷酸化保守位点[21]。这些保守的基序可能参与了DGAT3酶的翻译后修饰,但这些保守的基序对DGAT3活性的重要性仍有待通过定点诱变或其他方法进行验证。
通过分析基因的表达模式可以为基因的功能提供线索。本研究分析了3个CsDGAT3基因在亚麻荠各种组织中的表达谱,并发现了这3个基因在转录水平上的不同表达模式。总体而言,CsDGAT3-1主要在根中表达,CsDGAT3-2主要在花和萌发种子中发挥作用,表明这2个CsDGAT3基因在这些组织中的功能可能不同;但是,CsDGAT3-3在发育中的种子中表达水平高,这与种子油脂的快速积累相协调,表明CsDGAT3-3可能对亚麻荠种子中的TAG合成有较大贡献。YUAN等[12,20]研究发现,Cs-DGAT1、CsDGAT2和CsPDAT1家族中基因的3个拷贝和CsPDAT2家族中的2个同工型基因在各种组织中都有不同的表达模式,并且不同的基因参与了亚麻荠幼苗的不同胁迫反应。总而言之,在这些与TAG装配相关的酶基因家族中观察到的不同拷贝的不同表达模式可能在这些生物的发育中起着关键作用,表明存在多个拷贝,不仅不冗余而且还可增加功能多样性,进一步表明,这可能与真核生物在进化的过程中脂质代谢途径复杂性的增加相吻合。
本研究鉴定获得了亚麻荠的CsDGAT3基因,并解析了其分子特征和表达谱;3种CsDGAT3被预测为可溶性酶蛋白,以含有[2Fe-2S]结构域的同源二聚体形式存在。表达谱分析表明,3种Cs-DGAT3在不同组织中有不同的表达模式,表现为CsDGAT3-1在根中高表达、CsDGAT3-2主要在花和萌发种子中发挥作用,CsDGAT3-3在发育中的种子中高表达。本研究结果可为全面解析亚麻荠合成调控机制以及后续应用基因工程策略提高亚麻荠油脂的富集等方面提供重要的科学依据。
猜你喜欢
军事文摘(2022年16期)2022-08-24
湖北农业科学(2022年11期)2022-07-18
今日农业(2021年11期)2021-08-13
中国生殖健康(2020年4期)2020-12-09
实用肿瘤学杂志(2020年4期)2020-12-08
中西医结合肝病杂志(2020年2期)2020-10-27
散文诗世界(2017年3期)2017-11-13
家庭百事通·健康一点通(2016年7期)2016-08-04
纺织服装流行趋势展望(2016年6期)2016-05-04
纺织服装流行趋势展望(2016年2期)2016-05-04
