
表没食子儿茶素对TLR4/MyD88/NF-κB通路减轻脂多糖诱导BV2细胞炎症反应的影响*
2020-06-18 01:49张雯宋俊科朱晓瑜杨海光许启泰杜冠华
医药导报 2020年6期
张雯,宋俊科,朱晓瑜,杨海光,许启泰,杜冠华
(1.中国医学科学院北京协和医学院药物研究所,天然药物活性物质与功能国家重点实验室,药物靶点研究与新药筛选北京市重点实验室,北京 100050;2.海南绿槟榔科技发展有限公司,定安 571200)
神经炎症在脑血管疾病和神经退行性疾病的病理进程中扮演重要的角色[1-4]。研究表明,抑制炎症反应可以有效减轻卒中造成的脑部组织损伤,减轻脑水肿[5-6]。抑制炎症反应同样是预防和治疗帕金森病及阿尔茨海默病的有效手段[7-8]。植物多酚具有抗氧化、抗炎等多种药理学作用。多酚类化合物被认为是潜在的神经保护剂,能够调节信号传导、细胞凋亡、氧化还原和细胞分化等。多酚类化合物的摄入能够对抗神经元损伤,减缓疾病的进程。对多酚类化合物的研究,有助于揭示天然小分子化合物对机体的影响和作用,同时有助于新型化合物发现,指导新药研发。以往的研究中,对表没食子儿茶素没食子酸酯(epigallocatechingallate,EGCG)的研究较多,但对表没食子儿茶素(epigallocatechin,EGC)研究较少[9-10]。为了明确EGC是否具有神经细胞改善作用,笔者应用BV2小胶质细胞为研究对象,使用脂多糖(LPS)诱导BV2细胞炎症反应,研究EGC对LPS诱导BV2细胞炎症反应的抑制作用及机制。
1 材料与方法
1.1试剂 EGC购自上海源叶生物科技有限公司;达尔伯克改良伊格尔培养基(DMEM),胎牛血清,蛋白Marker均购自Thermo Fisher Scientific;多聚赖氨酸、LPS、Hoechst 33258、MTT均购自Sigma;一氧化氮(NO)检测试剂盒、细胞核蛋白与细胞浆蛋白抽提试剂盒,均购自碧云天生物技术有限公司;肿瘤坏死因子-α(TNF-α)、白细胞介素(IL)-1β和IL-6酶联免疫吸附测定(ELISA)检测试剂盒购自武汉华美生物工程有限公司;Toll样受体4(Toll-like receptor 4,TLR4)、髓样分化因子(myeloid differentiation factor 88,MyD88)、核因子κB(nuclear factor kappa-B,NF-κB)p65、β-actin、Lamin B1和Anti-rabbit IgG (H+L),F(ab')2Fragment (Alexa Fluor®488 Conjugate)抗体均购自Cell Signaling Technology;BCA蛋白定量试剂盒购自普利莱;Goat Anti-Rabbit IgG,HRP Conjugated和Goat Anti-Mouse IgG,HRP Conjugated均购自康为世纪生物科技有限公司。
1.2仪器 细胞培养瓶、96孔板均购自美国Costar;SpectraMax M5酶标仪购自美国Molecular Devices;Thermo Scientific ArrayScan Infinity高内涵分析仪购自Thermo Scientific;蛋白电泳仪及转膜仪购自美国Bio-Rad;Molecular Imager ChemiDoc XRS+ System成像仪购自美国Bio-Rad公司。
1.3细胞培养 用含10%胎牛血清的DMEM培养液,在37 ℃,5%二氧化碳(CO2)饱和湿度条件下培养BV2小胶质细胞。
1.4细胞模型建立及分组 选取对数生长期BV2细胞,接种于多聚赖氨酸包被的96孔板。建立LPS刺激细胞模型,LPS终浓度为1 μg·mL-1。实验分为正常对照组,模型对照组(1 μg·mL-1LPS),EGC小剂量组(1 μg·mL-1LPS + 5 μmol·L-1EGC),EGC中剂量组(1 μg·mL-1LPS + 10 μmol·L-1EGC)和EGC大剂量组(1 μg·mL-1LPS + 20 μmol·L-1EGC)。
1.5细胞活力检测 实验终点,吸去培养液,每孔加入0.5 mg·mL-1MTT溶液100 μL,37 ℃培养4 h后弃上清液,每孔加入DMSO100 μL,振荡5 min,用酶标仪在波长490 nm处测定吸光度(A值),计算细胞存活率。
1.6NO分泌的检测 实验终点,每孔取上清液50 μL,再加入Griess Reagent I 50 μL,混匀,Griess Reagent II 50 μL,混匀,在波长540 nm测定A值,计算NO浓度。
1.7炎症因子的检测 实验终点,吸去培养液,磷酸盐缓冲液(PBS)洗涤细胞后细胞转移到离心管中,用ELISA检测试剂盒测定TNF-α、IL-1β和IL-6的含量,具体操作按照说明书进行。
1.8Western blotting法检测EGC对炎症相关通路的影响 实验终点,吸去培养液,PBS洗涤3次,加入蛋白裂解液,冰上裂解细胞,提取胞浆和胞核蛋白,之后进行十二烷基磺酸钠-聚丙烯酰胺(SDS-PAGE)凝胶电泳。然后将SDS-PAGE凝胶转印至聚偏氟乙烯(PVDF)膜上,5% BSA室温封闭2 h后加入稀释的一抗,4 ℃共孵育过夜。TBST洗涤3次,再与二抗稀释液室温共孵育2 h。TBST洗涤3次后显影。
1.9免疫荧光法检测EGC对NF-κB p65核转位的影响 实验终点,弃去原培养液,PBS洗1次后用4%多聚甲醛固定15 min。之后PBS洗1次,加入0.3% Triton-100孵育10 min。再用PBS洗1次,加入5% BSA溶液封闭1 h,然后加入稀释的一抗4 ℃共孵育过夜。PBS洗3次,每次10 min,加入荧光二抗稀释液,室温孵育1.5 h。PBS洗3次,每次10 min,加入Hoechst,37 ℃避光孵育30 min,PBS洗1次,置于高内涵细胞分析系统进行检测。
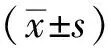
2 结果
2.1LPS和EGC对BV2小胶质细胞活力影响 结果见图1。实验结果表明,与正常对照组比较,LPS作用于BV2细胞24 h,细胞存活率无显著变化,说明1 μg·mL-1LPS对BV2细胞不存在细胞毒作用。EGC单独作用或与LPS共同孵育BV2细胞24 h后,与对照组比较,细胞活力差异无统计学意义。以上结果表明,在实验条件下,EGC(1~100 μmol·L-1)与LPS(1 μg·mL-1)对BV2细胞无明显毒性作用。
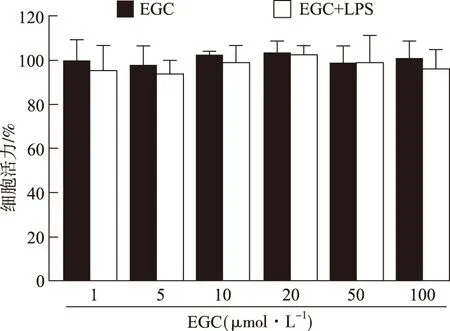
图1 LPS和EGC对BV2细胞活力的影响
Fig.1 Effects of LPS and EGC on the viability of BV2 cells
2.2EGC对LPS刺激BV2细胞NO分泌的影响 结果见图2。未加入刺激时BV2细胞上清液中NO含量较低,与正常对照组比较,给予LPS刺激后,NO含量显著增加(P<0.01)。20 μmol·L-1EGC能够显著抑制LPS诱导的NO产生(P<0.01)。
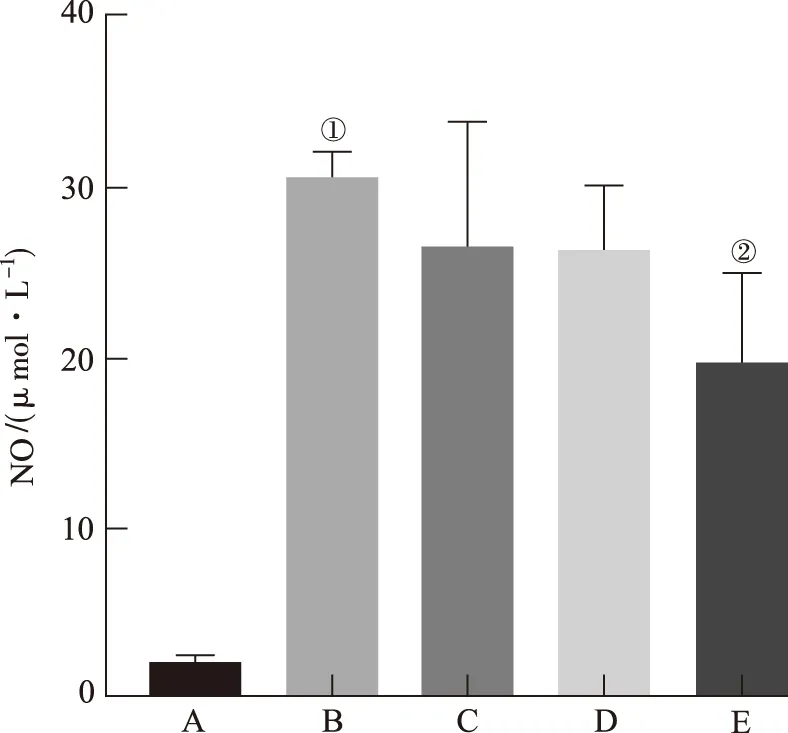
A.正常对照组;B.模型对照组;C.EGC小剂量组;D.EGC中剂量组;E.EGC大剂量组;①与正常对照组比较,P<0.01;②与模型对照组比较,P<0.01。
图2 EGC对LPS刺激BV2细胞NO分泌的影响
A.normal control group;B.model control group;C.low-dose EGC group;D.medium-dose EGC group;E.high-dose EGC group;①compared with normal control group,P<0.01;②compared with model control group,P<0.01.
Fig.2 Effects of EGC on NO production in LPS-stimulated BV2 microglia cells
2.3EGC对LPS刺激BV2细胞分泌TNF-α、IL-1β和IL-6的影响 通过ELISA法检测各组细胞中TNF-α、IL-1β和IL-6的含量。结果见图3。与正常对照组比较,LPS能够刺激BV2细胞分泌TNF-α、IL-1β和IL-6,其含量显著高于正常对照组(均P<0.01)。EGC能够显著抑制LPS引起的BV2细胞中TNF-α、IL-1β和IL-6的分泌,降低TNF-α、IL-1β和IL-6水平。
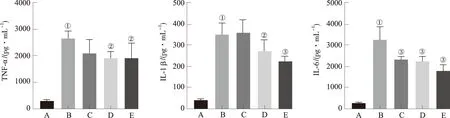
A.正常对照组;B.模型对照组;C.EGC小剂量组;D.EGC中剂量组;E.EGC大剂量组;①与正常对照组比较,P<0.01;②与模型对照组比较,P<0.05;③与模型对照组比较,P<0.01。
图3 EGC对LPS刺激BV2细胞分泌TNF-α、IL-1β和IL-6的影响
A.normal control group;B.model control group;C.low-dose EGC group;D.medium-dose EGC group;E.high-dose EGC group;①compared with normal control group,P<0.01;②compared with model control group,P<0.05;③compared with model control group,P<0.01.
Fig.3 Effects of EGC on the production of TNF-α,IL-1β and IL-6 in LPS-stimulated BV2 microglia cells
2.4EGC对LPS刺激BV2细胞中炎症通路的影响 Western blotting和免疫荧光法对LPS刺激的BV2细胞后TLR4/MyD88信号通路的变化进行研究。结果见图4。与正常对照组比较,LPS刺激后,BV2细胞中TLR4和MyD88的表达水平明显上升(均P<0.01)。与模型对照组比较,EGC能明显降低TLR4和MyD88的蛋白水平,抑制TLR4/MyD88信号通路的激活。
2.5EGC抑制LPS刺激BV2细胞中NF-κB核转位 进一步采用Western blotting和免疫荧光法,对BV2细胞中NF-κB p65的表达进行定位。结果见图5,LPS刺激后细胞中细胞核内NF-κB p65浓度显著升高(P<0.01),EGC可显著抑制LPS介导NF-κB p65核转位。同时,EGC也能显著抑制p-IκBα的升高,抑制炎症反应。
3 讨论
笔者在本文通过建立LPS刺激BV2小胶质细胞炎症反应模型,考察EGC对BV2小胶质细胞炎症反应的抑制作用。结果表明,EGC能够显著抑制炎症反应,并且其抗炎作用的发挥,可能与抑制TLR4/MyD88/NF-κB信号通路的激活,抑制NF-κB核转位有关。
TLRs是天然免疫系统中的关键因子。TLRs被激活后,可以通过MyD88依赖性信号通路传递炎症反应信息,介导炎症因子的表达和释放,产生炎性损伤[11-13]。NF-κB是重要的炎症调节因子,作为转录因子家族成员,能够介导TNF-α、IL-1β和IL-6等多种炎症介质和生长因子的转录与表达[14-16]。被NF-κB诱导表达的炎症因子进一步反馈活化NF-κB,诱导NF-κB的持续性活化,从而加重炎症损伤。本研究结果发现,1 μg·mL-1LPS和1~100 μmol·L-1EGC对BV2细胞无明显毒性作用。而LPS刺激后,BV2小胶质细胞NO释放量显著上升,20 μmol·L-1EGC能够显著降低LPS诱导的NO释放。LPS刺激后TNF-α、IL-1β和IL-6的分泌均显著高于正常对照组,10和20 μmol·L-1EGC能够不同程度抑制TNF-α、IL-1β和IL-6炎症因子的分泌。Western blotting和免疫荧光检测结果表明,LPS刺激BV2小胶质细胞后,TLR4/MyD88/NF-κB信号通路被激活,TLR4和MyD88的蛋白表达明显升高,而EGC能显著降低TLR4和MyD88的表达水平,抑制LPS诱导的NF-κB的核转位,抑制TLR4/MyD88/NF-κB信号通路的激活。
综上所述,EGC能够通过抑制TLR4/MyD88/NF-κB信号通路,抑制NF-κB核转位,进而减少TNF-α、IL-1β和IL-6等炎症因子的分泌,降低NO释放,从而有效抑制LPS所致BV2小胶质细胞炎症损伤。
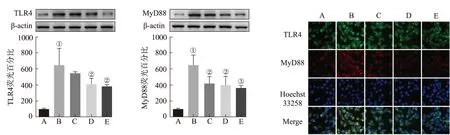
A.正常对照组;B.模型对照组;C.EGC小剂量组;D.EGC中剂量组;E.EGC大剂量组;①与正常对照组比较,P<0.01;②与模型对照组比较,P<0.05;③与模型对照组比较,P<0.01。
图4 EGC对LPS刺激BV2细胞TLR4/MyD88信号通路的影响
A.normal control group;B.model control group;C.low-dose EGC group;D.medium-dose EGC group;E.high-dose EGC group;①compared with normal control group,P<0.01;②compared with model control group,P<0.05;③compared with model control group,P<0.01.
Fig.4 Effects of EGC on TLR4/MyD88 signaling pathway in LPS-stimulated BV2 microglia cells

A.正常对照组;B.模型对照组;C.EGC小剂量组;D.EGC中剂量组;E.EGC大剂量组;①与正常对照组比较,P<0.01;②与模型对照组比较,P<0.05;③与模型对照组比较,P<0.01。
图5 EGC对LPS刺激的BV2小胶质细胞p-NF-κB和p-IκBα表达的影响
A.normal control group;B.model control group;C.low-dose EGC group;D.medium-dose EGC group;E.high-dose EGC group;①compared with normal control group,P<0.01;②compared with model control group,P<0.05;③compared with model control group,P<0.01.
Fig.5 Effects of EGC on p-NF-κB and p-IκBα levels in LPS-stimulated BV2 microglia cells
猜你喜欢
神经损伤与功能重建(2020年10期)2020-12-23
神经损伤与功能重建(2020年11期)2020-12-01
江西医药(2020年4期)2020-04-28
神经损伤与功能重建(2018年2期)2018-02-01
中成药(2017年9期)2017-12-19
中成药(2017年5期)2017-06-13
标记免疫分析与临床(2016年9期)2016-11-21
中国病理生理杂志(2015年8期)2015-12-21
中国生化药物杂志(2015年4期)2015-07-07
医学研究杂志(2015年3期)2015-06-10
