
北方储藏稻谷真菌多样性分析
2020-06-13 04:02祁智慧张海洋王鹏杰
中国粮油学报 2020年5期
祁智慧 张海洋 田 琳 王鹏杰 唐 芳 丁 轲
(北京农学院1,北京 102206) (国家粮食和物资储备局科学研究院2,北京 100037 )
我国稻谷的产量约占粮食总产量的三分之一[1],作为主要粮食作物,稻谷的储藏安全与人们的生活息息相关[1]。北方地区因其独特的生态气候条件,是我国优质粳稻的主产区和主要储存区。稻谷储藏时粮堆是一个复杂的生态系统,而北方地区粮库冬夏两季的环境温差极大,会导致粮堆内外温差加大,局部粮堆易出现霉变发热的问题[2],影响稻谷的储存安全。因此,针对北方粮仓储藏稻谷研究真菌的多样性,分析储藏环境对真菌分布的影响,通过监测分析真菌变化情况,用以指导北方稻谷储藏。
储藏稻谷的真菌种类数量与其产区地理位置、收获季节、气候条件或储藏方式有关[3]。早在20世纪80年代初,研究者曾对我国主要粮食,包括小麦、稻谷、玉米等的微生物多样性做了初步的调查[4-6],后来也有学者针对个别省份的稻谷真菌区系或多样性进行研究[7-8]。随着研究的不断深入,研究重点逐渐转向储藏条件对真菌的影响。田国军等[9]发现在不同储藏条件下由于上层稻谷受环境影响较大,其霉菌总数波动大于中、下层稻谷。周建新等[10]证实了稻谷储藏过程中的微生物区系演替与储藏温度和水分有关,并指出稻谷发生霉变的优势菌大多是曲霉,如白曲霉、米曲霉和黄曲霉。这些真菌区系及多样性的研究中,真菌的分离鉴定多以形态特征鉴定为主,部分菌种的鉴定可能存在不准确或不确定性,有必要采用现代分子生物学技术辅以形态学对稻谷储藏真菌多样性进行深入研究。
本研究选择北方地区有代表性的高大平房仓储藏的稻谷为研究对象,通过传统的真菌形态特征鉴定与现代分子生物学技术相结合将储藏稻谷中分离出的真菌基本鉴定到种。基于准确的真菌鉴定结果,结合实际仓储储藏条件的变化,对真菌多样性、真菌类型以及储藏条件进行分析,试图找出指示北方稻谷储藏条件变化的主要优势菌群及仓房重点关注位置,为北方仓储过程监控提供参考。
1 材料与方法
1.1 样品采集
参考2017年中国农业年鉴,依据粮食产量在北方地区采集稻谷样品。东北地区选择黑龙江、吉林、辽宁,华北地区选择北京,西北地区选择宁夏。每个省选择1个稻谷产量较高地区的粮库,抽取1~2个具有代表性的高大平方仓,选择储存一年的稻谷仓进行取样。
每仓5点3层取样,每点取200 g样品,取样点为粮仓的四个角加中间。3层的垂直分布为:上层(距离粮堆表面0.5 m左右)、中层(距粮面2~3 m左右)、底层(距离粮仓地面0.5 m左右),取样后将上、中、下层3层样品按层分别混合。
1.2 方法
1.2.1 菌落计数、分离及纯化
采用高盐察氏(High-salt CA)和察氏(CA)2种培养基同时对样品进行平板菌落计数和纯化,具体操作步骤参考GB 4789.15—2016《食品微生物学检验霉菌和酵母计数》,每份样品3个平行。计数时将不同表观形态特征的菌落分别标号,不同编号的菌株单独进行计数,挑取单菌落边缘菌丝转接3~4次,纯化不同编号真菌获得纯种菌株。样品前处理过程为称取25 g稻谷样品至盛有225 mL无菌水的锥形瓶中,在振荡器上振荡30 min,充分洗涤后,制备10倍系列稀释的样品匀液,选择2个适宜的稀释梯度进行接种。
1.2.2 真菌的表观及显微形态特征鉴定
表观形态特征鉴定采用三点接种法(或单点接种法)在高盐察氏培养(或察氏培养基)上接种,并置于(28±1)℃恒温培养箱中培养7 d,观察菌落特征并拍照,将相同特征的真菌归为一类进行保藏。微观形态特征鉴定采用斜插盖玻片培养的方法,使菌丝沿着盖玻片一侧往上生长,培养5 d后取出盖玻片,滴加乳酸—甘油固定液,在10×63倍的显微镜下观察拍照。菌种表观及显微形态特征的鉴定参照 GB/T 4789.16—2003《食品卫生微生物学检验常见产毒霉菌的鉴定》、GB/T 26628.1—2011《粮油检验储粮真菌标准图谱》、《真菌鉴定手册》和《中国真菌志》等资料。
1.2.3 真菌的分子生物学鉴定
采用液氮研磨和DNA提取试剂盒提取真菌DNA, 提取DNA后进行PCR扩增,扩增所用引物为 ITS1(5′-CTTGGTCATTTAGAGGAAGTAA-3′)和ITS4(5′-TCCTCCGCTTATTGATATGC-3′)以及bT2a (5′-GGTAACCAAATCGGTGCTGCTTTC-3′)和bT2b(5′-ACCCTCAGTGTAGTGACCCTTGGC-3′)[11,12]。用PCR提纯试剂盒对 PCR 产物进行纯化,通过琼脂糖凝胶电泳检测有正确条带后进行测序。测序成功后用 BioEdit 软件去除多余序列,保留波峰较整齐部分的序列,进行拼接后得到ITS和β-微管蛋白基因序列,将得到的序列分别与GeneBank数据库中的序列进行BLAST同源序列比对,确定分离菌株的菌种信息。
2 结果与分析
2.1 菌种鉴定、系统发育树构建及真菌分类
对高盐察氏和察氏培养基分离出的所有表观形态特征略有差异的322株分离菌株进行分子生物学鉴定。结合ITS和β-微管蛋白基因序列比对的同源序列结果以及真菌的形态特征,将分离出的菌株初步确定种属名称。基于ITS序列,采用最大似然法(Maximum likelihood)将所有分离菌种构建系统发育树,对分离菌种的种属进一步确认,并对所有真菌进行分类分析。
由图1可知,北方地区储藏期稻谷共分离出真菌58种,分布在2门,4纲,6目,13科,16属。属水平分别为链格孢属Alternaria、弯孢属Curvularia、毛霉属Mucor、横梗霉属Lichtheimia、共头霉属Syncephalastrum、根毛霉属Rhizomucor、附球菌属Epicoccum、亚隔孢壳属Didymella、枝孢属Cladosporium、Gibellulopsis属、帚枝霉属Sarocladium、 木霉属Trichoderma、镰刀菌属Fusarium、 蓝状菌属Talaromyces、青霉属Penicillium和曲霉属Aspergillus,其中曲霉属真菌种类最多为24种,青霉属和镰刀菌属次之,分别为9种和7种。分离获得的菌种以散囊菌目Eurotiales成员为最多,共计34种,分布在蓝状菌属Talaromyces、青霉属Penicillium和曲霉属Aspergillus。非散囊菌目的其他菌种在系统树上排布较为分散,分属于5目,11科, 13属。
通过系统发育树可以看出,北方地区储藏稻谷分离出的真菌种类较多。部分类群的ITS序列存在着极高的相似性,在种的划分上很多分支自展支持率小于50%,特别是链格孢属Alternaria、曲霉属Aspergillus和青霉属Penicillium真菌,表明ITS序列对真菌种水平上的鉴别能力不足,通过ITS测序仅可以将北方储藏稻谷分离出的大部分真菌鉴定到属,很少部分鉴定到种。因此对稻谷真菌多样性进行深入分析时,应结合形态学特征并补充其他特征引物条码进行分析。本研究对于ITS未确定到种的菌株,补测了β-微管蛋白基因的序列。β-微管蛋白是真核生物细胞内细胞器微管的成分之一,构成细胞内骨架结构,通过β-微管蛋白编码基因序列分析,大部分情况下可进一步将属水平或属内复合群水平的真菌进一步的鉴定到种[13]。综合分析ITS和β-微管蛋白基因序列比对的同源序列结果及真菌形态特征,将322株分离菌株,确认为58种真菌,55种真菌鉴定到种水平,有3株真菌未确定到种,但可以确认分别为附球菌属Epicoccum、镰刀菌属Fusarium和曲霉属Aspergillus的物种。
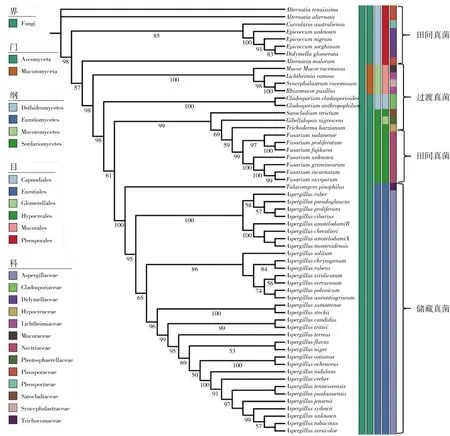
注:分支下面数据表示该分支以bootstrap进行检验,1 000次重复获得50%以上的自展支持率。图1 北方储藏稻谷真菌基于ITS构建的ML系统发育树及真菌分类
不同种类的真菌对于环境的适应性不同,对于仓储稻谷的潜在危害性也不同。针对这一点,国外学者通过系统研究,结合粮食收获和储藏环节,根据真菌生长适宜的水活度范围对粮食真菌进行了划分[14],如图2所示。

图2 基于真菌生长适宜水活度范围的真菌分类[14]
由图2可看出,粮食真菌主要分为3种类型:1)田间真菌,亲水型植物病原真菌,粮食作物生长或收获期侵染粮食籽粒;2)过渡真菌,对水活度要求相对宽泛,以腐生和嗜热真菌为主,收获前后污染谷穗和破碎颗粒或受伤的植株;3)储藏真菌,嗜旱真菌,对水活度要求相对低,主要在粮食储藏期间侵染粮粒[15]。本研究根据Mannaa等[14]和Fleurat-Lessard[15]对粮食真菌3种类型的总结,初步将北方地区储藏稻谷分离到的全部真菌进行划分。因基于ITS序列构建的系统发育树可以表征不同菌株之间亲缘关系的远近[16],这里对文献中未提及的部分真菌物种,根据其在系统发育树上亲缘关系相近的物种进行聚类划分,并对3种类型进行标注,具体分类标注情况见图1。同一类型真菌在系统发育树上基本聚在一起,这是由于粮食真菌对于水分和温度的要求由其特征基因编码决定的[17]。通过本研究将我国北方仓储稻谷中分离出的真菌进行全面准确的鉴定,并进行了分类,为北方稻谷储藏状况分析提供借鉴。
2.2 北方储藏稻谷真菌菌落数及菌种丰度
粮食储藏期间,侵染粮食的真菌以耐旱的储藏真菌为主,从系统发育树上看,大部分真菌属于曲霉属。曲霉菌属的部分成员(在之前的命名法中属于散囊属Eurotium的真菌)极易污染各种低水活度的生物基质,特别是谷物及其制品[18-20]。《中国真菌志》中介绍散囊属Eurotium真菌都是嗜高渗透压的,在一般的培养基上生长不良或不能生长,在低水活度培养基上才能生长正常[21]。因此,选取高盐察氏培养基分离计数结果对后续储藏稻谷的真菌菌落数、菌种丰度和优势菌群等进行分析。真菌菌落总数和菌种丰度结果见表1。

表1 北方各省储藏稻谷真菌菌落总数及菌种丰度
由表1可知,东北三省和华北地区北京真菌菌落数和菌种丰度相差不大,真菌菌落数基本处于1.0×103~ 2.2×103CFU/g的水平上,菌种丰度在16 ~ 21种。四个省区的稻谷在储藏过程中基本实现了准低温储藏,黑龙江和吉林因其外界自然低温环境优越,仓内主要采用内环流控温或夜间开窗自然通风降温,辽宁和北京主要采用空调控温。这4个省区粮库,度夏期间,整仓平均温度在10 ℃左右,上层温度均控制在20 ℃左右,个别地区出现过25 ℃,但持续时间不长。因此,这些地区储藏稻谷的真菌菌落数基本维持在初始带菌量的水平,真菌丰度也没有明显变化。而西北宁夏粮库稻谷真菌菌落数明显增加,达到104CFU/g水平,表层菌种丰度明显下降。宁夏该粮库夏季高温持续一段时间,但粮库未采取控温措施,追溯粮堆温度数据,整仓平均温度在15 ℃左右,上层曾出现过局部32 ℃ 高温现象。综合分析,宁夏上层稻谷水分和温度偏高,为储藏真菌生长提供了充足条件,致使真菌菌落数增加,高于正常储藏时菌落数一个数量级,并成为优势菌群,使菌种丰度降为3种。
2.3 优势菌群分析
由于储藏条件发生变化,真菌菌落数会明显增加,具体哪些真菌会生长成为优势菌群值得关注。明确优势菌群的种类有助于监控仓储过程中主要危害真菌变化情况,为粮堆霉变防控提供技术指导。
对高盐察氏培养基分离到的菌种进行优势菌群分析,首先确定北方储藏稻谷真菌的优势属,再将优势属中真菌放在一起进行分析,占比小于1%的真菌种类归为其他,优势属和优势种比例关系如图3所示。
由图3可知,用高盐察氏培养基分离时,北方储藏稻谷优势属主要为曲霉属Aspergillus、枝孢属Cladosporium和链格孢属Alternaria,优势种为曲霉属Aspergillus中的阿姆斯特丹曲霉Aspergillusamstelodami和多育曲霉Aspergillusproliferans,枝孢属Cladosporium中的芽枝状枝孢Cladosporiumcladosporioides和Cladosporiumanthropophilum,链格孢属Alternaria中的细极链格孢Alternariatenuissima和链格孢Alternariaalternata。
由2.1分析可以确定,链格孢属Alternaria的细极链格孢Alternariatenuissima和链格孢Alternariaalternata属于田间真菌,在收获前期侵染稻谷,其生长对水活度要求较高。枝孢属Cladosporium的芽枝状枝孢Cladosporiumcladosporioides和Cladosporiumanthropophilum属于过渡真菌,在收获前后污染稻谷,其生长特性与田间真菌相似。这两类真菌通常不会在储藏期间大规模生长繁殖,主要分布在各省高大平方仓粮堆的中层和下层。本研究团队曾对北方新收获入库稻谷的真菌多样性进行调查,结果表明大部分北方稻谷入库时就携带大量的枝孢属真菌,这可能与北方稻谷收获期间环境温度有关。当仓储环境一直处于低温、条件良好时(如粮堆中层和下层),这些田间和过渡真菌仍保持收获入库时的优势比例。曲霉属Aspergillus的阿姆斯特丹曲霉Aspergillusamstelodami和多育曲霉Aspergillusproliferans属于储藏真菌。这2种真菌有嗜高渗透压的特性(耐旱性)[21],能在低水活度的粮食中生长和繁殖,主要出现在宁夏粮库粮堆上层。这类真菌是北方稻谷储存过程中主要监控的危害真菌种类。
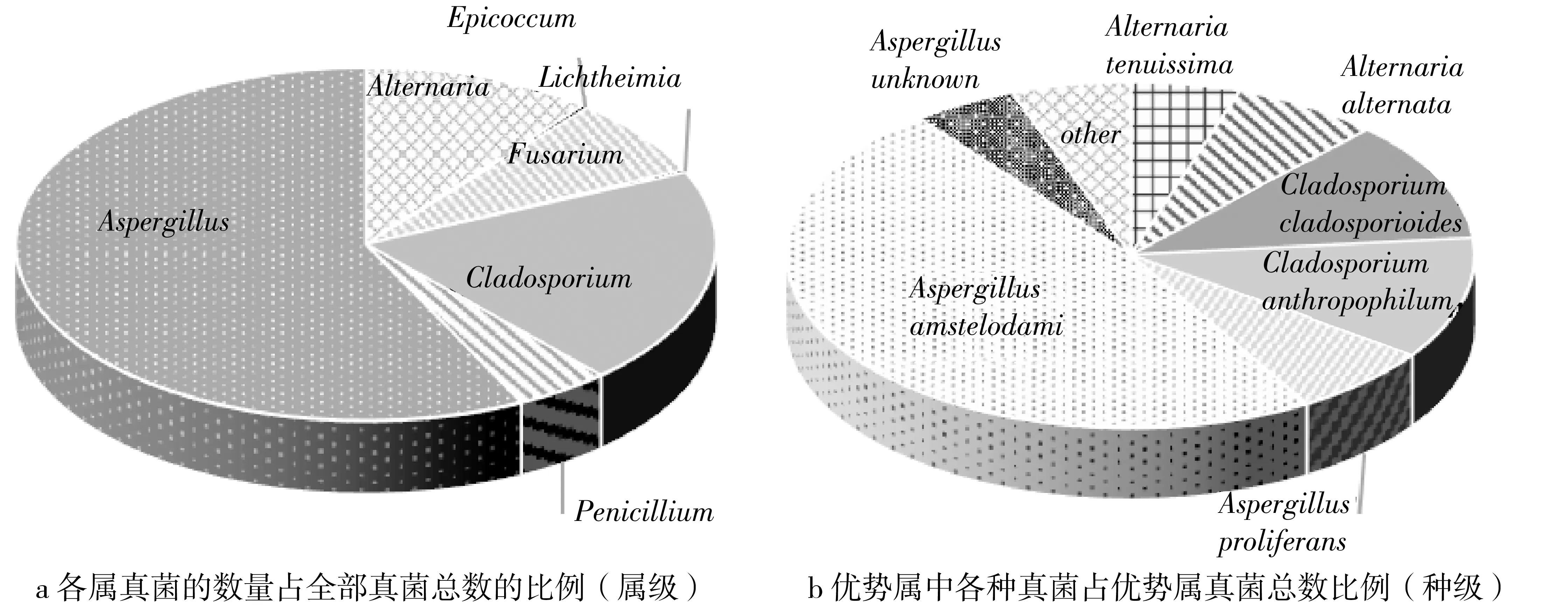
图3 储藏稻谷优势属和优势种分布情况
本研究分离出的优势菌阿姆斯特丹曲霉Aspergillusamstelodami和多育曲霉Aspergillusproliferans与拟灰绿曲霉Aspergilluspseudoglaucus、赤曲霉Aspergillusruber、筛曲霉Aspergilluscibarius、谢瓦曲霉Aspergilluschevalieri、Aspergillusmontevidensis5种真菌的形态特征和ITS序列很相近(见系统发育树),由于阿姆斯特丹曲霉Aspergillusamstelodami、赤曲霉Aspergillusruber和谢瓦曲霉Aspergilluschevalieri归属于灰绿曲霉群[22],故将这7种真菌统一归为灰绿曲霉群及其近缘类群进行分析,这个类群所有真菌均具有嗜高渗透压的特性(耐旱性)[21],属于典型的储藏真菌,其生长需要较低的水活度。由此可知,在稻谷储存过程中,储存条件不佳时,灰绿曲霉群及其近缘类群最先生长繁殖,数量快速增加逐渐取代其他真菌成为优势菌群,使真菌种群丰度下降。
2.4 储藏环境对真菌分布的影响
从表1中真菌数量及丰度数据可以看出粮堆作为一个小生态环境,不同位置温湿度不同,可能会影响到真菌分布情况。本研究根据粮堆环境差异特点,按照方法1.1进行分层取样,对3种类型的真菌在粮堆不同深度的分布情况进行分析,结果如图4所示。

图4 北方储藏稻谷不同深层位置真菌分布情况
由图4可知,北方地区高大平房仓储存稻谷时,粮堆不同深度,不同类型真菌分布存在一定差异。结合北方地区气候特点,冬季干冷,粮库通常会在冬季进行通风降温蓄冷以备度夏。粮堆会保存自然冷心,粮堆中层和下层平均粮温可控制在10 ℃或以下,夏季期间,上层稻谷易受外界环境影响,粮温升高,平均温度可达20~25 ℃,个别区域会出现30 ℃高温,当上层和中层稻谷温差过大时可能出现水分迁移现象导致上层稻谷水分升高[23],温度和水分的上升,易导致储藏真菌的生长。因此,在粮堆上层储藏真菌占绝对优势,田间和过渡真菌的数量由于真菌之间的竞争抑制作用而降低。对于贴近粮仓墙面0.5 m左右的粮堆,当墙体保温隔热效果不佳时,受环境温度影响明显,也可能出现这种情况。中层和下层稻谷因粮食导热性差,受外界环境影响相对较小,会保持良好的低温环境,田间真菌和过渡真菌占主要优势,基本维持在初始水平,大概范围为1.0×103~3.3×103CFU/g,属于正常环境真菌的检出水平。
北方地区粮仓内,当储藏真菌成为优势菌群后,田间和过渡真菌数量会明显下降,这一不同类型真菌的生长和演替,主要受储存条件变化的影响。北方储藏稻谷主要优势菌为灰绿曲霉群及其近缘菌群,这类真菌生长一般不会导致粮堆发热,但若不加以控制,可能会出现发热现象,因此灰绿曲霉群及其近缘菌群可以作为粮堆发热前预测预警的指示菌群[24]。
3 结论
北方地区储藏期稻谷分离出的真菌种类较多,共分离纯化出322株分离菌株,经鉴定为58种真菌,分布于2门,4纲,6目,13科,16属。本研究综合分析ITS和β-微管蛋白基因序列以及真菌形态特征,将我国北方储藏稻谷分离出真菌基本鉴定到种。通过系统发育树分析,结合国外文献对粮食中真菌类型的划分,首次将我国北方仓储稻谷中分离出的真菌进行了分类,为北方稻谷储藏状况分析提供理论支撑。
通过对北方储藏稻谷优势菌群和真菌类型的分析,将阿姆斯特丹曲霉Aspergillusamstelodami和多育曲霉Aspergillusproliferans与其他5种近缘曲霉划为灰绿曲霉群及其近缘类群,这类真菌对于仓储环境的适应性较强,储存温湿度增加时,首先萌发生长,数量增加,成为优势菌群。高大平房内,粮堆上层或邻近墙壁等位置受环境温度的影响,容易出现高温和水分迁移的情况,为储藏真菌生长提供条件,使优势菌群由初始的田间和过渡真菌演替为储藏真菌,若不加以处理可能导致粮堆发热,引起储存稻谷品质的劣变,严重影响储粮安全。综上所述,我国北方地区储藏期稻谷分离出的真菌种类繁多,真菌菌落总数范围在103~ 104CFU/g之间。优势菌阿姆斯特丹曲霉Aspergillusamstelodami和多育曲霉Aspergillusproliferans被划分为灰绿曲霉群及其近缘类群,可作为北方粮堆异常粮情早期监测预警的主要指示菌群,高大平方仓中粮堆上层和邻近墙壁等位置是北方仓储过程中主要关注的区域。
猜你喜欢
科教新报(2021年39期)2021-11-04
河南工业大学学报(自然科学版)(2021年2期)2021-06-01
河南工业大学学报(自然科学版)(2020年6期)2021-01-29
今日农业(2020年15期)2020-12-15
农业工程学报(2020年8期)2020-06-04
河南工业大学学报(自然科学版)(2020年2期)2020-05-22
农村百事通(2018年24期)2018-02-22
农村农业农民·B版(2017年2期)2017-03-11
科学启蒙(2016年11期)2016-12-09
四川党的建设(2015年9期)2015-09-15
