
铅胁迫对垂柳生长和光合生理特性的影响
2020-06-10 02:13薛文秀尚晓硕邹金华
天津师范大学学报(自然科学版) 2020年3期
薛文秀,尚晓硕,蒋 艺,邹金华
(1.天津师范大学生命科学学院,天津300387;2.天津师范大学 天津市动植物抗性重点实验室,天津300387)
土壤污染中,铅是传播最广且污染最为严重的重金属污染物之一.当铅在土壤或水中积累到一定程度时,就会通过食物链进入到人体内,影响人体健康.关于铅胁迫对植物产生毒害作用的研究已有一些报道:如Hou 等[1]通过测量叶绿素荧光、抗氧化酶活性、有机酸含量和Pb 含量等指标探究了金丝草对铅的响应机制;王颖等[2]通过研究不同浓度的铅对水稻分蘖情况、植株高度、生物量分配、不同部位铅含量的影响,分析了铅对水稻生长的胁迫作用.
植物修复是使用绿色植物提取和去除土壤中重金属污染物的技术,具有高效和低成本的特点,适用于大面积重金属污染土壤的净化.目前已知的超富集草本植物有虉草(Phalaris arundinacea)、芦苇(Phragmites australis)、香蒲(Typha orientalis)等[3]. 草本植物由于生长矮小、生物量低,在植物修复中往往只作为科学研究的对象.速生木本植物由于具有生物量大、根系较深而发达、易繁殖、不与食物链相连等特性在植物修复中备受关注.如欧美国家广泛种植短轮伐期柳树人工林用于废水更新和土壤修复,同时其生物量转化为生物质能源,兼具生态效益和经济效益[4].
垂柳(Salix babylonica)是我国重要的绿化树种,具有传播广、适应性强、生长快等特点,是一种铅高富集和高耐受性植物[5]. 本研究用不同浓度的Pb(NO3)2溶液处理垂柳水培枝条,测试不同程度的铅胁迫下垂柳植株生长、光合色素合成以及光合生理特性的变化,以此探究垂柳在生长和光合作用方面对铅胁迫做出的响应.
1 材料和方法
1.1 实验材料
在天津师范大学校园内选取长势较好的垂柳,剪取长度约为15 cm 的枝条.
1.1.1 试剂
硝酸铅(Pb(NO3)2)、丙酮(CH3COCH3)、Hoagland营养液(5 mmol/L Ca(NO3)2、5 mmol/L KNO3、1 mmol/L KH2PO4、50 μmol/L H3BO3、1 mmol/L MgSO4、4.5 μmol/L MnCl2、3.8 μmol/L ZnSO4、0.3 μmol/L CuSO4、0.1 μmol/L(NH4)6Mo7O24、10 μmol/L Fe-EDTA).
1.1.2 仪器
Lambda35 紫外可见分光光度计, 美国Perkin Elmer 公司;DUAL-PAM/F 便携式PAM 荧光仪,德国WALZ 公司.
1.2 方法
1.2.1 植物培养与处理
选取直径相近的垂柳枝条,将其剪成长度约为15 cm 的短枝并分成5 组,每组24 枝,把短枝固定在纸板上,分别放置于5个相同容量的盛有自来水的盆中进行生根培养,每隔1 周更换1 次自来水.水培2 周后,再使用Hoagland 营养液培养1 周. 之后,在5个盆中加入同等体积的蒸馏水,再分别加入不同量的Pb(NO3)2,使其浓度分别为0、1、10、50、100 μmol/L.用气泵通气,每天观察垂柳生长情况,定期换水.
1.2.2 垂柳插条根长和株高的测量
Pb(NO3)2溶液处理垂柳插条28 d 后,随机测量5组插条的根长和新生枝条的株高,分别记录15个数据.
1.2.3 叶绿素的提取和测定
Pb(NO3)2溶液处理28 d 后,在5个处理组中分别取0.2 g 叶片,冲洗干净,放进研钵中,加入5 mL 80%的丙酮溶液在避光条件下研磨,研磨充分后倒入锥形瓶中并封紧,避光过夜.次日早上,在避光条件下,将锥形瓶中的液体倒入15 mL 离心管中,在4 ℃、4000 r/min的条件下离心20 min.离心后将上清液转入到平底试管中并使用80%的丙酮溶液定容至10 mL.摇匀后倒入比色皿中,用分光光度计分别测量波长为663、646、470 nm 时的吸光度值,将数值带入公式中分别计算叶绿素a(Ca)、叶绿素b(Cb)和类胡萝卜素(Cc)的含量,单位均为mg/g[6].
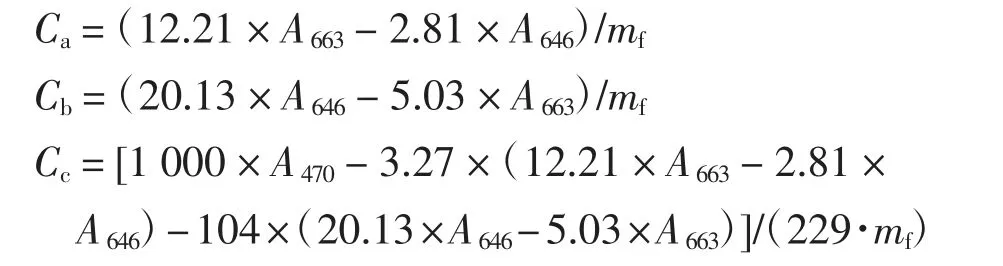
式中:A663、A646、A646分别为波长663、646、470 nm时测量的吸光度值;mf为样品的鲜重(g).
1.2.4 叶绿素荧光参数的测定
Pb(NO3)2溶液处理28 d 后,将5个处理组的垂柳枝条分别黑暗处理30 min,之后选取生长良好的叶片,使用便携式PAM 荧光仪测量叶绿素荧光参数,包括初始荧光(F0)、最大荧光(Fm)、PSⅡ最大光化学量子效率(Fv/Fm)、光化学淬灭系数(qP)、非光化学淬灭系数(qN 或NPQ)、PSⅡ光化学能量转化的有效量子产量(Y(Ⅱ))[7].根据数据计算可变荧光(Fv=Fm-F0)和PSⅡ潜在活性(Fv/F0).
F0是PSⅡ反应中心处于完全开放时的荧光产量,反映了PSⅡ光反应中心的活性.Fv/Fm是PSⅡ最大光化学量子产量,反映PSⅡ反应中心的内禀光能转换效率,胁迫条件下该参数明显下降[8].qP 反映PSⅡ原初电子受体质体醌A(QA)的还原状态[9].qN 或NPQ 是指热耗散引起的荧光猝灭,qN 为NPQ 的总系数,它是植物在光照过强时的一种自我保护机制,反映植物耗散过剩光能为热的能力,NPQ 表示光适应下的非光化学猝灭,也是植物的一种自我保护机制,它反映的是PSⅡ天线色素吸收的光能不能用于光合电子传递而以热的形式耗散的部分.
1.3 数据分析
用Excel 进行数据统计分析,用SigmaPlot 13.0 和Photoshop CS6 作图,采用SPSS 17.0 对数据进行差异统计学检验.
2 结果与分析
2.1 Pb 胁迫对垂柳根长和株高的影响
不同浓度的Pb(NO3)2溶液处理28 d 后垂柳的形态如图1 所示,株高和根长情况如图2 所示.

图1 不同处理组中垂柳的形态Fig.1 Morphology of S.babylonica with different treatments

图2 不同处理组中垂柳的株高和根长Fig.2 Shoot height and root length of S.babylonica with different treatments
由图1 可以看出,1 μmol/L Pb(NO3)2处理组的垂柳形态与对照组相近;10 μmol/L 处理组中株高明显降低,根长和侧根数量下降,但地上分枝数并未减少且叶片无异常情况;50 μmol/L 和100 μmol/L 处理组中垂柳株高进一步降低,根的茂盛程度下降,分枝数减少,部分叶片变黄甚至脱落.由图2 可以看出,与对照组相比,1 μmol/L 的Pb(NO3)2处理组中,垂柳的株高和根长均增加(P <0.05);10、50 和100 μmol/L 处理组中垂柳的株高和根长均显著降低,且溶液浓度越大,垂柳的株高和根长越小.
2.2 Pb 胁迫对垂柳叶片光合色素含量的影响
不同浓度的Pb(NO3)2溶液处理后垂柳叶片的光合色素含量变化如图3 所示.
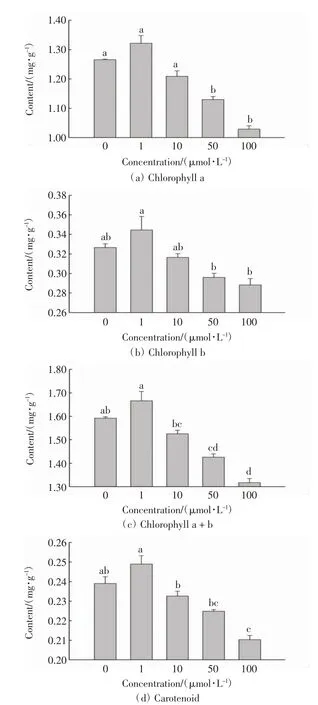
图3 不同处理组中垂柳的光合色素含量Fig.3 Photosynthetic pigment content of S.babylonica with different treatments
由图3 可以看出,与对照组相比,在浓度为1 μmol/L的Pb(NO3)2溶液的处理下,垂柳叶片中叶绿素a、叶绿素b、叶绿素a+b、类胡萝卜素的含量均有所增加,但与对照组的差异不具有统计学意义(P >0.05).随着Pb(NO3)2溶液浓度的进一步增加,垂柳叶片光合色素含量呈下降趋势,10、50 和100 μmol/L 浓度处理下叶绿素a、叶绿素b、叶绿素a+b、类胡萝卜素的含量均低于对照组,尤其是50 μmol/L 和100 μmol/L 处理组中,垂柳叶片的光合色素含量同对照组的差异均具有统计学意义(P <0.05). 由此可见,低浓度的铅胁迫(1 μmol/L)对垂柳叶片的光合色素含量具有轻微促进作用,这可能是植物对重金属胁迫作出的应激反应,即一定范围内的重金属胁迫会刺激植物增加光合色素的合成.铅胁迫超过一定程度时(≥10 μmol/L),垂柳叶片的光合色素生成受到抑制.
2.3 Pb 胁迫对垂柳叶片叶绿素荧光参数的影响
不同浓度的Pb(NO3)2溶液处理下,垂柳叶片的叶绿素荧光参数如表1 所示.
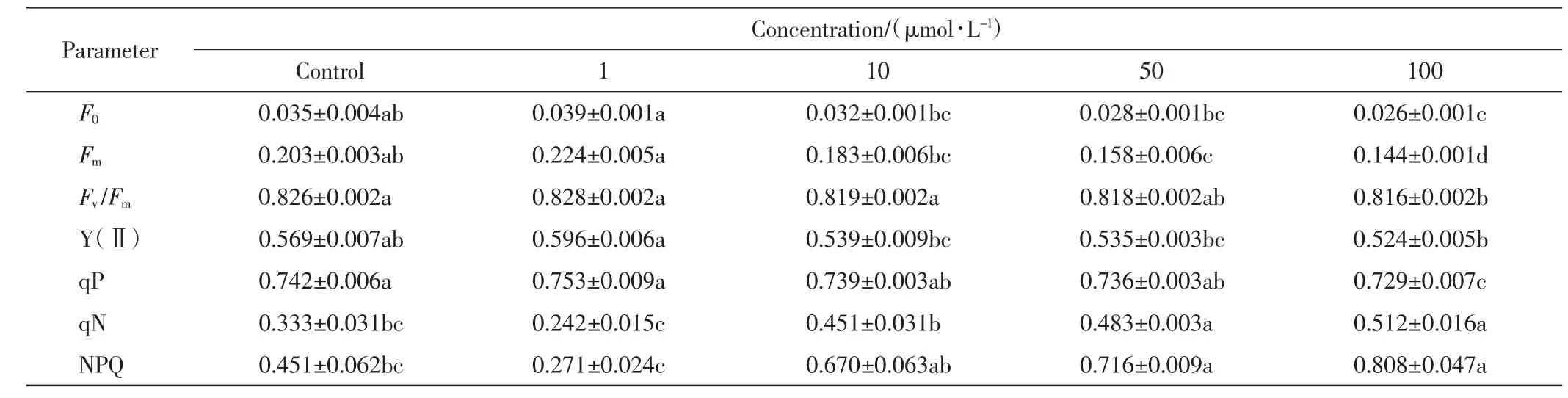
表1 不同处理组中垂柳的叶绿素荧光参数Tab.1 Chlorophyll fluorescence parameters of S.babylonica with different treatments
由表1 可以看出,随着Pb(NO3)2浓度的增加,垂柳叶片的初始荧光F0、最大荧光Fm、PSⅡ最大光化学量子效率Fv/Fm、PSⅡ光化学能量转化的有效量子产量Y(Ⅱ)、光化学猝灭系数qP 均呈现先增加后减小的趋势,Pb(NO3)2溶液的浓度为1 μmol/L 时达到最大值,Pb(NO3)2溶液超过10 μmol/L 时垂柳叶片的叶绿素荧光效率则受到抑制,且随着Pb(NO3)2浓度的增加抑制作用增强.非光化学猝灭系数qN 和NPQ随Pb(NO3)2浓度增加的变化趋势则相反.
3 讨论
本研究对垂柳水培枝条施加不同程度的铅胁迫,观察铅胁迫对垂柳生长和光合生理特性的影响.结果发现,低浓度(1 μmol/L)的Pb(NO3)2溶液对垂柳生长具有一定的促进作用,高浓度的Pb(NO3)2溶液(10、50和100 μmol/L)对垂柳生长则具有明显的抑制作用,且随着浓度增加,抑制作用增强.铅胁迫下,垂柳地上和地下部分的生长均受到抑制,但根部受抑制的程度明显高于地上部分. Zou 等[10]研究了不同程度的镉胁迫对旱柳生长的影响,发现根部是镉吸收和积累的主要部位;王晓娟等[11]也认为,植物根部是重金属污染最敏感的部位,这是因为根部与重金属直接接触,使得重金属在根部积累较多,为了避免重金属对植物的危害,植物体产生了多种限制重金属吸收和转移的生理生化机制,因此相比于根部,地上部分所受抑制较小.
光合作用和新陈代谢能否正常进行与光合色素的含量密切相关[12].与对照组相比,在低浓度(1 μmol/L)的铅胁迫下,垂柳的叶绿素a、叶绿素b、叶绿素a+b、类胡萝卜素的含量均增加,而在高浓度(10、50 和100 μmol/L)的铅胁迫下,光合色素含量随铅胁迫程度的增加降低.王娅玲等[13]认为低浓度的铅胁迫可以增强植物体内叶绿体的酶活性,促进叶绿素的合成,从而使叶绿素含量增加;高浓度的铅可能会导致叶绿体酶活性失调,从而加快叶绿素分解. 邹洪梅等[14]也指出,高浓度的金属离子会抑制某些酶的活性,而这些酶与叶绿素生物合成密切相关,导致叶绿素合成受阻,叶绿素含量下降.
叶绿素荧光动力学反映了环境因子对植物光合作用的影响,因此常用于检测环境胁迫对植物的影响以及植物对环境胁迫的响应机制[15].本研究中,在高浓度(10、50 和100 μmol/L)Pb(NO3)2溶液的处理下,随处理浓度的增加,F0、Fm、Fv/Fm、Y(Ⅱ)、qP 的数值逐渐减小,qN、NPQ 的数值则逐渐增大,这与黄轩等[8]的研究结果相似.在高浓度Pb(NO3)2溶液的处理下,Fv/Fm值和Y(Ⅱ)值的降低说明铅胁迫下垂柳叶片的光合机构受到了一定程度的伤害,PSⅡ反应中心的活性被抑制,因而叶片对光能的吸收和转化效率明显下降,暗反应中光合碳同化所需的能量积累被阻碍[16].qP 为光化学淬灭系数,在一定程度上反映了该反应中心的开放度高低,而非光化学猝灭系数qN 是指叶片吸收的光能以热量形式耗散的程度,qN 为NPQ 的总系数,NPQ数值大小反映的是植物对光合机构的保护程度[8].在高浓度Pb(NO3)2溶液的处理下,qP 的下降和qN 的上升说明反应中心的开放度逐渐降低,叶片吸收的光能更多的以热能形式耗散,热耗散可以抵御光抑制带来的破坏,并且保护PSⅡ反应中心免受损伤.相反,在低浓度的Pb(NO3)2溶液(1 μmol/L)处理下,与对照组组相比,垂柳叶片的F0、Fm、Fv/Fm、Y(Ⅱ)、qP 数值均显著增加,qN、NPQ 的数值则有所减小,说明环境中少量的铅会促进植物的光合作用,提高光能的吸收和转化效率,增大反应中心的开放程度.
总的来看,培养基质中铅的含量较低时,垂柳的生长、光合色素生成及光合生理均得到促进,铅含量较高时,垂柳生长和光合色素合成均受到抑制,叶片光合机构受到伤害,PSⅡ反应中心的开放度降低,叶片吸收的光能更多地以热能形式释放出去以减少重金属铅的伤害.
猜你喜欢
娃娃乐园·综合智能(2022年9期)2022-08-16
当代作家(2022年4期)2022-05-13
现代盐化工(2021年3期)2021-08-27
科学大众(2021年9期)2021-07-16
烟台大学学报(自然科学与工程版)(2021年1期)2021-03-19
小天使·一年级语数英综合(2020年3期)2020-12-16
食品与生物技术学报(2020年4期)2020-01-06
小天使·一年级语数英综合(2017年11期)2017-12-05
新课程·下旬(2015年10期)2015-10-21
红蜻蜓·低年级(2014年11期)2015-04-23
