
小麦抗麦红吸浆虫候选基因TraesCS4A01G437800 的四引物ARMS-PCR 标记开发
2020-06-09 09:02郝志明张力菁温树敏刘桂茹王睿辉
河北农业大学学报 2020年2期
郝志明,章 悦,张力菁,温树敏,刘桂茹,王睿辉
(河北农业大学 农学院/华北作物种质资源研究与利用教育部重点实验室,河北 保定 071001)
麦红吸浆虫 (Sitodiplosis mosellana Géhin) 是严重影响我国小麦生产的重要害虫,以幼虫吸食灌浆期小麦籽粒汁液造成为害,使受害籽粒空瘪,导致小麦产量和品质下降。一般情况下,受害麦田减产幅度在10%~60%之间,严重为害时甚至颗粒无收[1-2]。在包括加拿大、美国、英国、德国在内的小麦主产国,均有麦红吸浆虫造成严重损失的报 道[3-7]。在我国,麦红吸浆虫曾于20 世纪50 年代和80 年代两度严重爆发,造成小麦大规模损失[8-9]。21 世纪以来,麦红吸浆虫危害仍为严重。近年来,我国小麦吸浆虫年均发生面积在2.0×106hm2左右 (http://www.agri.gov.cn/),已成为影响华北、黄淮和西北麦区小麦生产的重要虫害 (https://www.natesc.org.cn/)。生产上,可通过喷洒农药、轮作倒茬或种植抗虫小麦品种等方法对吸浆虫进行防治[4,10-11],其中选用抗虫品种是最为经济、安全、有效的控制措施[12]。虽然20 世纪选育的抗吸浆虫小麦品种如‘西农6028’‘南大2419’等对防治虫害发挥了作用,但目前抗虫品种匮乏的现状仍未得到显著改变[8]。同时,由于吸浆虫危害的毁灭性,小麦对吸浆虫抗性遗传的复杂性以及人工抗虫鉴定的不稳定性,使得小麦抗虫育种进展缓慢[13-14]。而分子标记辅助选择 (Marker-assisted selection, MAS) 技术在小麦等主要作物育种中的应用,能够显著提高针对目标性状的育种效率,同时有利于实现目标性状与其他优良性状的同步改良[15-16]。因此,开发与抗吸浆虫基因/QTL 紧密连锁甚至共分离的分子标记,不仅能够提高抗虫育种效率,还有利于实现抗虫性与其他优良品质性状的聚合。
Sm1 是最早发现并被定位的小麦抗麦红吸浆虫基因[3,17],与该基因连锁的标记Xbarc35 和XWM1已被应用于小麦MAS 育种中[18];后来Kassa 等人又找到7 个与抗虫性紧密连锁的竞争性等位基因特异性PCR (KASP, Kompetitive Allele-Specific PCR) 标记,可用于鉴定抗/感虫单倍型[19]。而中国小麦品种可能携带了与Sm1 不同的抗性基因。来自国外抗虫基因Sm1 的连锁标记虽已被应用于当地小麦抗虫品种的培育中,但这些标记在我国小麦品种中要么不具多态性,要么检测结果与抗虫性不相符,并不适用于我国小麦品种的抗虫性鉴定[20]。有鉴于此,开发我国小麦种质中与抗虫位点紧密连锁甚至共分离的分子标记,不但有助于准确鉴定我国小麦种质资源的抗虫性,也能更好地满足国内抗虫分子标记辅助育种的需要。
在前期研究中,通过对构建的2 个重组近交系 (RIL) 群体进行连锁分析,将我国小麦品种中的抗虫主效QTL 位点 (QSm.hbau-4A) 定位于4A 染色体上[13,20-21];并利用混池转录组测序 (BSR-Seq)[22]方法结合 QTL-Seq[23]的分析思路,对抗/感虫小麦样本 (亲本、混池及株系) 进行了转录组数据分析,结合基因功能注释、qRT-PCR 及基因序列分析,在4AL 抗虫候选区间中挖掘到6 个与抗虫性紧密相关的基因[24]。其中2 个基因 (TraesCS4A01G436100, TraesCS4A01G437800) 不仅富集于与抗虫性相关的异黄酮生物合成代谢通路 (Isoflavonoid biosynthesis pathway) 中,而且在抗、感近等基因系间的表达水平呈现显著差异,因此为重点关注的基因。但由于基因TraesCS4A01G436100 在抗虫亲本‘冀麦24’中的序列上存在一个无义突变,导致编码蛋白结构不完整,因此推测基因TraesCS4A01G437800 为抗虫候选基因的可能性更高[24]。后来,利用上述6个抗虫相关基因序列中存在的插入缺失 (InDel) 和单核苷酸多态性位点 (SNP) 成功开发了8 个标记,其 中6 个KASP 标 记 (K3-1-1、K3-7-1、K3-7-3、K3-16-1、K10-10-6 和K10-13-x) 和1 个EST 标 记 (E1-2) 可较好地检测供试品种在QSm.hbau-4A 位点上的基因型[25]。虽然KASP 技术可以很好地将基因序列中存在的差异位点 (SNP/InDel) 转化为可分型的标记,并可以简便、快速地检测标记在供试材料中的分型,但开发成本较高[26],对于需要进行大量分型检测的MAS 而言,开发和使用成本较低且通量高的分子标记更具重要意义。四引物扩增受阻突变体系PCR (四引物ARMS-PCR, Tetra-primer amplification refractory mutation system PCR) 标记体系是一种在普通PCR 基础上发展起来的单核苷酸多态性分型技术,与KASP 标记在原理及开发上具有相似性,即这2 种标记都是基于核苷酸序列中的差异位点,以突变位点作为引物的3’末端,并以相应3’末端碱基所对应的扩增目的片段的有无来确定分型结果[27]。四引物ARMS-PCR 标记解决了传统方法分型通量低的问题,具有简便、快速和费用低廉等特点,但对扩增条件要求更为严格,目前较为广泛地应用于医学[28-29]和水稻[30-32]研究中,在小麦中的应用相对较少[33]。
因此, 本研究根据推定抗虫候选基因TraesCS4A01G437800 序列上的SNP 位点,设计并开发四引物ARMS-PCR 标记,并利用抗/感虫RIL株系和不同抗性的小麦品种验证标记在小麦吸浆虫抗性鉴定中的可用性。
1 材料与方法
1.1 植物材料及DNA 提取
用于标记分型检测的植物材料包括抗虫小麦亲本‘冀麦24’、感虫小麦亲本‘6218’,从6218/冀麦24 重组近交系 (RIL) 群体中选择的47 个抗虫RIL 株系和45 个感虫RIL 株系,以及95 个具有不同抗性水平的小麦品种 (表1)。供试样本在培养皿中生长,待生长至两叶一心时剪取新鲜叶片,采用CTAB 法[34]提取小麦基因组DNA。
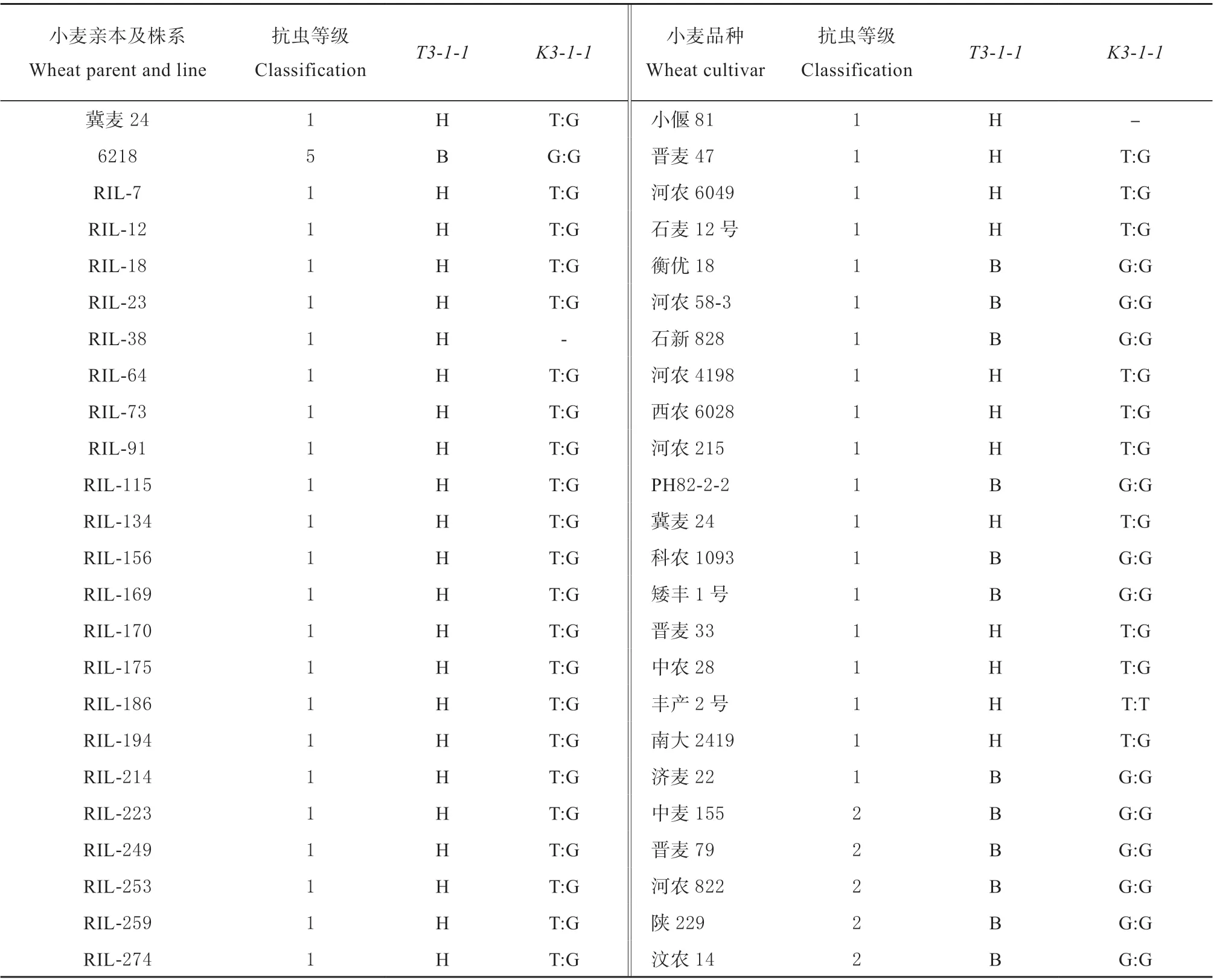
表1 供试小麦材料的表型及基因型Table 1 The phenotypes and genotypes for selected wheat materials

续表:
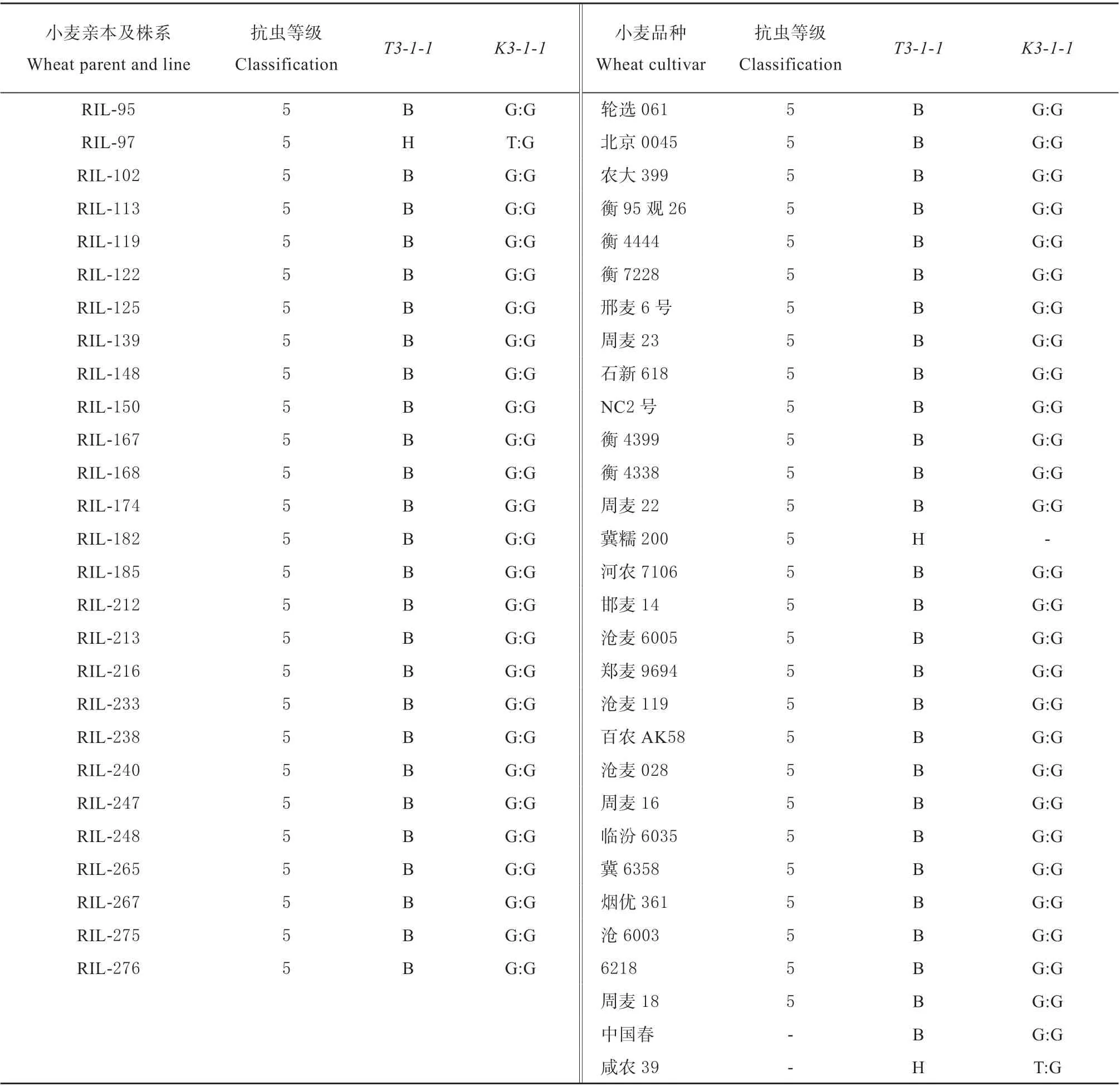
续表:
1.2 供试小麦材料的抗虫性鉴定
利用抗性指数对小麦材料的抗虫性进行评价[13]。 在小麦灌浆至乳熟期时,每重复、每株系取10 ~15个穗子,密封于纸袋中,带回室内对籽粒上的虫子进行计数。根据籽粒上吸浆虫数目,将待鉴定小麦株系的籽粒抗性分为5 级,根据以下公式计算每个株系的估计损失率(L)[2,13]:
L(%) = Σ(xf) / 4Σf × 100%
式中,x 为相应子粒的抗性级别,f 为各级别的籽粒数目。
用每个株系的估计损失率 (L) 除以全部参试株系( 品种) 的平均估计损失率 (ML),计算出抗性指数 (RI) 来确定株系的抗性分级。当同一株系在不同重复间的鉴定结果存在较大差异时,以最重重复数据进行L、RI 值的估算[2]。基于RI 值将供试小麦材料的抗性分为免疫 (RI=0) (Immune, I)、高抗 (0.01 ≤RI ≤0.19) (Highly resistant, HR)、 中抗 (0.20 ≤RI<0.49) (Moderately resistant, MR)、低 抗 (0.50 ≤RI<0.99) (Lowly resistant, LR)、感 虫 (1.00 ≤RI<1.50) (Susceptible, S) 和高感 (RI ≥1.50) (Highly susceptible, HS)。
1.3 抗虫相关基因序列检测
对推定抗虫候选基因TraesCS4A01G437800[35]进行四引物ARMS-PCR 标记开发。从‘中国春’参考基因组 (IWGSCv1.0, https://wheat-urgi.versailles.inra.fr/) 中提取该基因的参考序列及其侧翼序列,用Primer 5 软件设计测序引物 (表2)。利用高保真酶扩增该基因在小麦亲本 (‘冀麦24’‘6218’) 中的序列。PCR 扩增采用10 μL 反应体系,包括模板DNA 50 ng、正反引物各0.4 μmol/L、1×Pfu PCR MasterMix (天根生化科技(北京)有限公司)。PCR反应条件为:94 ℃预变性3 min;94 ℃变性30 s,55 ℃退火30 s,72 ℃延伸2 min,35 个循环;72 ℃ 终延伸5 min。扩增产物经1 %琼脂糖凝胶电泳分离,对目的条带进行回收,送生工生物工程(上海)股份有限公司进行双向测序。将测序结果与‘中国春’ (IWGSCv1.0) 的基因组序列进行比对。

表2 基因TraesCS4A01G437800 的测序引物Table 2 Sequencing primer for gene TraesCS4A01G437800
1.4 四引物扩增受阻突变体系PCR 标记的设计与开发
根据基因的测序结果,对抗、感亲本间存在的SNP 位点设计并开发四引物ARMS-PCR 标记。根据SNP 侧翼各300 bp 的序列设计多对外引物,并根据SNP 两侧相邻序列设计退火温度、GC 含量较为适宜的内引物,再利用引物设计软件 (Primer 5) 对设计的引物质量进行评判,选择评分最好的引物进行后续试验。对设计好的引物采用Taq DNA 聚合酶进行PCR 扩增。优化的扩增反应体系包括模板DNA 50 ng、外引物各0.16 μmol/L、内引物各0.24 μmol/L、1×Es Taq MasterMix (5 μL)。扩增反应程序为:94 ℃预变性5 min;94 ℃变性30 s,57 ℃退火30 s,72 ℃延伸1 min,35 个循环;72 ℃终延伸7 min。扩增产物在30 %的聚丙烯酰胺凝胶上电泳,银染显色。统计全部样本的基因型 (带型),分别计算该标记在高抗品种、中抗品种、感虫品种和高感品种中的检测有效率 (即检测基因型与某一表型吻合的品种数占该表型品种数的百分比),计算方法如下:
检测有效率(抗虫品种)=检测出抗虫基因型的抗虫品种数/抗虫品种总数。
检测有效率(感虫品种)=检测出感虫基因型的感虫品种数/感虫品种总数。
2 结果与分析
2.1 基因序列分析
对抗虫相关基因TraesCS4A01G437800 的扩增产物进行琼脂糖凝胶电泳检测,发现该基因在抗、感小麦亲本中均能扩增到目的大小的条带。将目的条带的测序结果与参考序列比对,结果显示该基因在两亲本中的测序结果与参考基因序列相符。在两小麦亲本 (‘冀麦24’‘6218’) 间进行序列比对,发现存在1 个使氨基酸发生改变的非同义突变SNP,位于748 bp 处,该位点在‘中国春’和感虫亲本‘6218’中的碱基为“G”,而在抗虫亲本‘冀麦24’中的碱基为“T” (图1)。将这个SNP 位点及其侧翼序列与小麦参考基因组 (IWGSCv1.0) 比对,发现在参考基因组中全部同源序列对应的碱基位点上,都不存在突变型碱基“T”,即该SNP 在抗虫亲本中的突变为基因组特异性突变,因此该特异性SNP 位点可用于标记开发。
2.2 四引物ARMS-PCR 标记T3-1-1 的多态性检测
根据基因TraesCS4A01G437800 上的特异性SNP 位点设计了四引物ARMS-PCR 标记T3-1-1 (表3)。经检测,T3-1-1 在两小麦亲本间均能检测出相应的分型,且具有多态性。该标记在两亲本中,均能扩增出170 bp 的外引物扩增条带;在感虫亲本‘6218’中能扩增出111 bp 的条带 (B 带型),在抗虫亲本‘冀麦24’中能扩增出96 bp 和111 bp 的杂合条带 (H 带型),所扩增出条带片段大小符合预期 (图2)。根据该标记位点侧翼序列与小麦参考基因组序列 (IWGSCv1.0) 的比对结果得知,该序列在参考基因组中存在多个同源序列,且每个同源序列在标记位点上的碱基均为“G”,因此无论在抗、感虫小麦亲本中均能扩增出等位基因型 “G” 的条带 (111 bp);而在抗虫亲本中,标记位点的碱基为“T”,该突变在基因组中任何一个同源序列上均不存在,因此只能在抗虫亲本中扩增出等位基因型 “T” 的条带 (96 bp)。根据这一扩增特性,可以在供试材料中检测出具有抗虫标记位点和不具有抗虫标记位点的小麦品种(株系)。

图 1 基因TraesCS4A01G437800 在两小麦亲本间 (‘6218’与‘冀麦24’) 的序列比对Fig. 1 Sequence comparisons of gene TraesCS4A01G437800 aligned with two wheat parents (‘6218’ and ‘Jimai 24’)
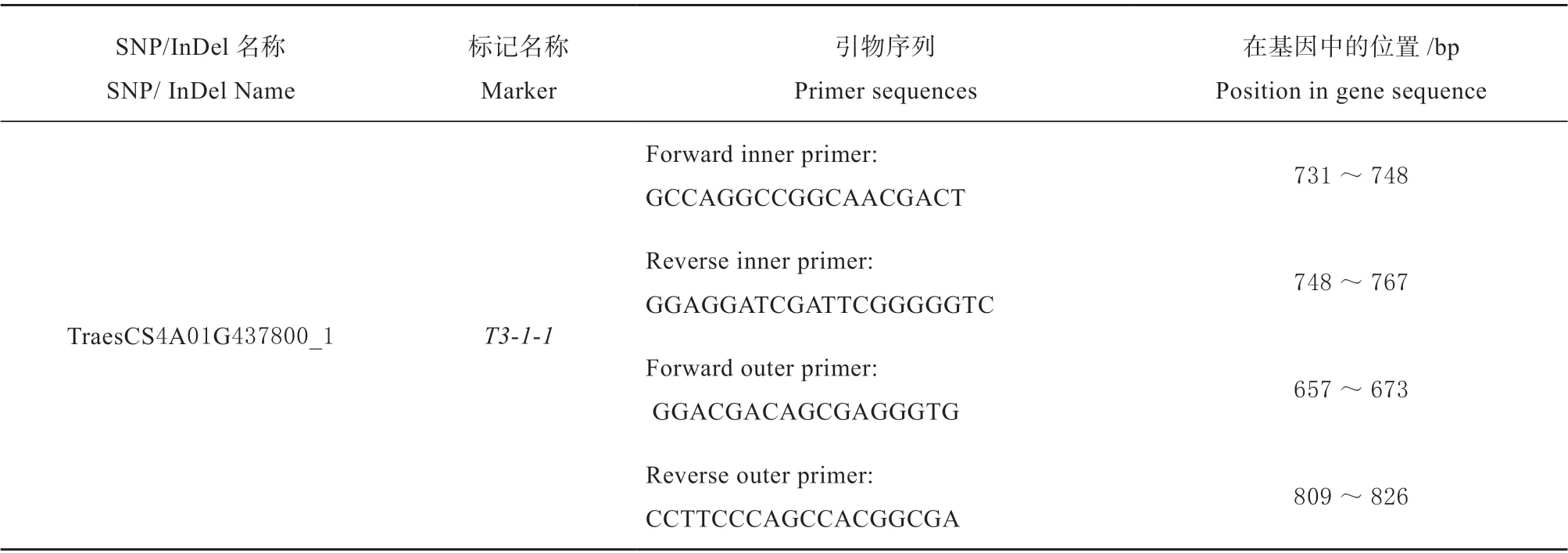
表3 四引物ARMS-PCR 标记T3-1-1 的引物序列Table 3 Primer sequences for tetra-primer ARMS-PCR marker T3-1-1
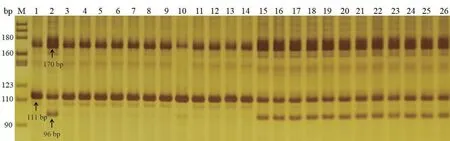
图2 四引物ARMS-PCR 标记T3-1-1 在抗、感虫小麦亲本及部分RIL 系中的扩增结果Fig. 2 Molecular detection of two parents and partial RIL lines by tetra-primer ARMS-PCR markers of T3-1-1
2.3 四引物ARMS-PCR 标记T3-1-1 的有效性检测
检测结果显示,T3-1-1 在小麦抗虫RIL 株系中的检测有效率为87.23 %,在感虫RIL 株系中的检测有效率为95.56 %。在供试95 个小麦品种中,该标记在高抗品种、中抗品种、感虫品种和高感品种中的检测有效率分别为63.16 %、12.50 %、75.00 %和96.43 % (图3);即在34.88 %的抗虫品种 (高抗和中抗) 中检测出抗虫标记位点 (H 带型),在90.% 的感虫品种 (感虫和高感) 中检测出纯合感虫标记位点 (B 带型)。这说明,如果在供试品种中检测到抗虫标记位点,则该品种为抗虫品种 (具有QSm.hbau-4A 抗虫位点) 的可能性约为90 %、为感虫品种 (基因型与表型不相符) 的概率为10 %,因此这个标记可以较好地筛选具有QSm.hbau-4A 位点的抗虫小麦种质。
利用该标记,检测出20 份小麦品种具有抗虫位点 (带型同抗虫亲本‘冀麦24’),其中包括14 个抗虫品种,分别是‘南大2419’ (1932 年引进种质)、‘中农28’ (1932 年引进种质)、‘西农6028’ (1956 年审定,下同)、‘丰产2 号’ (1968)、‘晋麦33’(1985)、‘冀麦23’ (1986)、‘河农215’ (与‘冀麦23’、‘冀麦24’互为姊妹系)、‘晋麦47’ (1998)、‘邯麦9 号’ (2003)、‘石家庄11 号’ (2003) 、‘石麦12 号’ (2004)、‘河农4198’ (2005)、‘小偃81’ (2005)、‘河农6049’ (2008),5 个感虫或中间型 (低抗) 品种 (‘洛麦21’‘石家庄8 号’‘藁优9908’‘河农826’‘冀糯200’),1 个无表型鉴定结果的品种 (‘咸农39’) (表1)。可以看出,在具有抗虫位点的14 个抗虫品种中,多为生产上已停止推广的老品种,比较新的品种审定时间也在10年以上。因此,利用已有的抗虫小麦品种和新开发的分子标记鉴定和创新小麦抗虫种质资源尤为重要。
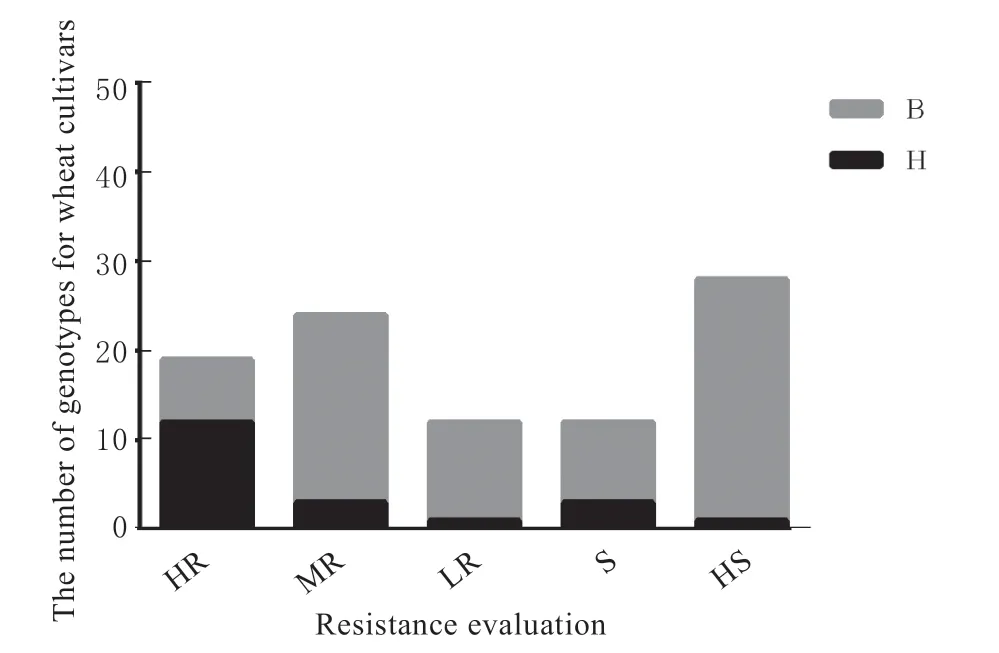
图3 标记T3-1-1 在供试小麦品种中不同抗性级别下各基因型所占个数Fig. 3 The number of genotypes for T3-1-1 in tested wheat cultivars for each resistance evaluation
3 讨论
3.1 四引物ARMS-PCR 标记与KASP 标记检测结果的比较
在前期研究中,基于基因TraesCS4A01G437800 上的这个SNP 位点已成功开发1 个KASP 标记K3-1-1,通过检测发现K3-1-1 同样可在抗虫亲本中检测出杂合分型T:G (同H 带型),在感虫亲本中检测出纯合分型G:G (同B 带型)。该标记在抗虫RIL 株系中的检测有效率为86.67%,在感虫RIL 株系中的检测有效率为 95.56%;在供试高抗小麦品种、中抗品种、感虫品种和高感品种中,该标记的检测有效率分别为61.11%、13.64%、75.00%和100.00%[25]。可以看出,本研究基于该SNP 位点开发的四引物ARMSPCR 标记T3-1-1 与KASP 标记K3-1-1 在相应供试材料间的检测有效率基本一致。根据表1 可知,除6份分型数据缺失的小麦材料 (RIL-38、RIL-218、‘小偃81’‘白硬冬2 号’‘烟农23’和‘冀糯200’) 外,标记K3-1-1 与T3-1-1 仅在1 个品种中的检测结果不同,即T3-1-1 对‘丰产2 号’检测出杂合型 (H 带型),而K3-1-1 对该品种检测出纯合抗虫分型T:T。由于理论上K3-1-1 和T3-1-1 对供试材料的检测结果仅可能为杂合型或纯合感虫基因型,不会检测出纯合抗虫基因型,因此推测K3-1-1 对‘丰产2 号’检测出纯合抗虫分型可能是由于该品种在标记位点两侧的序列与其同源序列差异较大所造成的,有待后续检验。除在‘丰产2 号’中的检测结果不同外,K3-1-1与T3-1-1对其他全部材料的基因型检测结果均相同,表明四引物ARMS-PCR 标记技术是开发小麦分子标记的可行方法,所开发的标记可准确检测供试小麦材料的基因型。因此,本研究开发的四引物ARMSPCR 标记可对小麦种质资源进行抗虫等位基因型选择,并为在小麦中开发分子标记的方法提供了一个重要参考。
3.2 四引物ARMS-PCR 标记T3-1-1 的应用及展望
本研究根据前期BSR-Seq 分析结果,结合qRTPCR、KEGG Pathway、基因功能注释及基因序列分析初步确定的抗虫候选基因TraesCS4A01G437800上的SNP 位点,利用四引物ARMS-PCR 技术开发了1 个抗虫相关标记T3-1-1。在供试小麦材料中进行基因型检测发现,该标记在感虫小麦品种中的检测有效率高 (90%),而在抗虫小麦品种中的检测有效率较低 (34.88 %),表明在一些小麦品种的基因组中可能存在不同于QSm.hbau-4A 的抗虫基因/QTL。由于T3-1-1 在感虫小麦品种中的检测有效率高达90%,因此在供试感虫品种中检测出抗虫基因型的可能性低 (即检测错误率为10%);反之,只要在供试材料中检测出抗虫基因型,则该品种实际为抗虫品种的可能性在90%左右。因此该标记可作为专门筛选抗虫小麦种质的标记。通过图3 可以看出,大多数高抗小麦品种 (63.16%) 都具有抗虫等位位点,而其他表型的品种很少具有该抗虫等位位点,因此该标记可以很好地检测供试小麦品种是否具有QSm.hbau-4A 抗虫位点,对我们筛选具有QSm.hbau-4A 位点抗虫种质以及鉴定早代育种材料的抗虫性具有重要帮助。本研究中,有14 个抗虫品种被检测到携带了标记T3-1-1 的抗虫等位基因 (表1),这些品种多为生产上已经停止推广的老品种,比较近的品种其审定时间也在10 年以上,因此有效利用老品种创新小麦抗虫资源的任务十分迫切。
本研究基于混池转录组测序并结合多种方法, 快速准确地实现了潜在功能标记的开发。虽然目前尚未实现抗虫主效QTL 的图位克隆,但所获得的抗虫相关标记T3-1-1 在作图群体和不同抗性小麦品种中被证明是有效的, 不仅能用于鉴定小麦种质资源抗虫性, 还可对小麦抗吸浆虫主效QTL 的精细定位和图位克隆提供重要帮助。
4 结论
本研究基于抗虫候选基因TraesCS4A01G437800序列上的SNP 位点成功设计并开发了1 个四引物ARMS-PCR 标记,该标记在抗、感RIL 株系间的检测有效率在90%左右,在抗虫小麦品种中的检测有效率为34.88%,而在感虫品种中的检测有效率可达90%,因此对于鉴定出抗虫基因型的小麦品种来说,其表型符合基因型 (为抗虫品种) 的可能性为90%左右,可用于抗虫小麦种质资源筛选与抗虫性MAS育种。此外,具有抗虫位点的14 个抗虫小麦品种多为生产上已停止推广的老品种,因此鉴定并创新小麦抗虫种质资源尤为重要。
猜你喜欢
分子催化(2022年1期)2022-11-02
西南农业学报(2022年5期)2022-06-06
昆明医科大学学报(2021年3期)2021-07-22
中国糖料(2021年3期)2021-07-13
烟草科技(2021年6期)2021-06-24
林业科技(2020年3期)2021-01-21
热带农业科技(2019年1期)2019-01-14
中国果业信息(2019年11期)2019-01-05
电脑知识与技术(2018年19期)2018-11-01
江苏农业科学(2016年10期)2017-02-05
