
气候变化情景下四川省猕猴桃溃疡病菌潜在地理分布模拟
2020-06-08 09:41王茹琳刘原李庆
植物保护 2020年2期
关键词:气候变化
王茹琳 刘原 李庆


摘要 近年来,猕猴桃溃疡病在四川各猕猴桃主产区严重发生,造成严重经济损失。本研究采用MaxEnt模型分析四川省猕猴桃溃疡病菌潜在分布,并预测2030年代、2050年代、2070年代和2080年代的RCP2.6、RCP4.5和RCP8.5等3种气候变化情景下适生区变化。预测结果运用ROC曲线评价模拟准确性。结果表明:所建立13个模型的训练数据和测试数据AUC (areas under curve)值均高于0.9,达到极高的精度。当前气候条件下,猕猴桃溃疡病菌在四川的高适生区主要位于成都市、德阳市、绵阳市、广元市、巴中市、达州市和雅安市,中适生区在四川21地市州均有分布。2030年代-2080年代,气候变化情景下,与当前情景相比,高适生区和低适生区区域均显著增加,中适生区区域先增加后减少,不同适生区几何中心位置和迁移规律均有所不同但总体上均向北移动。
关键词 气候变化; 潜在地理分布; MaxEnt模型; 猕猴桃溃疡病菌
中图分类号: S 431.2 文献标识码: A DOI: 10.16688/j.zwbh.2019055
Abstract In recent years, the kiwifruit canker disease has occurred seriously in the main kiwifruit producing areas of Sichuan, and caused serious economic losses. In this study, MaxEnt (the maximum entropy model) was applied to analyze the potential geographic distribution of Pseudomonas syringae pv.actinidiae (Psa) in Sichuan. The future distribution of Psa were also predicted for 2030s, 2050s, 2070s and 2080s under the climate change scenarios of RCP2.6, RCP4.5 and RCP8.5. The results showed that AUC (areas under curve) values of training data and test data of all models were higher than 0.9, which indicated a better forecast. Under current climate situation, the highly suitable areas for Psa were in Chengdu, Deyang, Mianyang, Guangyuan, Bazhong, Dazhou, and Yaan, whereas the moderately suitable areas covered 21 cities of Sichuan. Under climate change scenarios, the areas of highly and least suitable areas increased significantly compared with the current scenario, while the areas of the moderately suitable areas decreased significantly. The geometric center location and migration rule of different suitable areas were different, but they all moved northward in general.
Key words climate change; potential geographical distribution; MaxEnt; Pseudomonas syringae pv. actinidiae
四川是紅心猕猴桃的发源地,主栽品种有‘红阳‘ 东红 ‘红什2号和‘金红50等,其中以‘红阳猕猴桃栽培面积最大,种植面积超过3.33万hm2,占全省栽培面积的68.53%。猕猴桃种植作为四川省扶贫攻坚的主要产业,近年来在全省大范围推广,种植面积和产量分别占全国22%和16.6%,均居全国第二位[1]。由于种植品种以红心猕猴桃为主,且在地区间引种、购苗、采粉及果品销售活动中缺乏必要的检疫措施,猕猴桃溃疡病在猕猴桃主栽区迅速蔓延,发生面积逐年扩大,在成都、广元、德阳、崇州、雅安等市均有发生,给种植户造成了巨大经济损失。据统计,2017年,猕猴桃溃疡病在四川发病面积达1.04万hm2,约为全省猕猴桃种植面积的26%,对产业发展造成极大威胁[2-3]。
区域或全球大尺度背景下,物种地理分布主要决定于气候条件,气候变化对病虫害发生、消长、蔓延和流行的影响主要表现为两点:首先世界性气候改变可能会引起新病虫害出现,改变病虫害的主次位置,增大或减少病虫害地理分布范围[4]。其次气候变化改变寄主植物的分布范围,进而引起病虫害生境的变化[5]。利用当前气象数据评价病虫害的分布及危害,并依据可能出现的气候事实,借助多种气候变化模式,预测未来病虫害的风险等级,具有重要指导意义。生态系统中每一个物种都有其特定的小生境,生态位模型可根据特定小生境所对应的环境因子,分析物种在不同时间和空间存在的可能性,获得物种潜在地理分布[6]。最大熵模型(MaxEnt)是目前被国内外学者广泛应用且具有较高评价的一种数学模型,该模型在植保方面应用主要包括分析重大病虫害气候适宜性、预测检疫性病虫害入侵可能性和模拟气候变化对病虫害分布区影响等[7-10]。
猕猴桃溃疡病危害部位包括主干、枝条、花和叶片,其病原菌为丁香假单胞杆菌猕猴桃致病变种Pseudomonas syringae pv.actinidiae, Psa,依靠农事操作、苗木、花粉、风雨和昆虫等方式传播[11-12]。研究表明,猕猴桃溃疡病的发生和危害与气候关系密切,气候适宜时蔓延扩散极快[13-14]。室内培养发现,猕猴桃溃疡病菌生长适宜温度范围为5~30℃,最适温度为25℃,最高温度为35℃,致死温度为55℃[15-16]。猕猴桃溃疡病发病时间、发病程度、流行速度和停止蔓延时间与温度关系密切。国内学者在不同地区调查了猕猴桃溃疡病田间发病条件,结果表明该病菌5℃时开始繁殖,田间最适发病温度范围为12~16℃,在此温度范围内,病害扩展迅速。20~25℃时,阴雨天发病概率大,高于25℃时,田间病斑停止扩展[15, 17]。猕猴桃溃疡病病原菌是低温型病菌,低温是导致溃疡病发病的主要原因。同时低温冻害易导致树体受冻伤,猕猴桃溃疡病菌易从伤口侵染树体[18]。四川盆地气温东高西低,南高北低,冬季平均温度4~8℃之间,夏季平均温度在24~28℃之间,非常适宜猕猴桃溃疡病菌生存[19]。影响猕猴桃溃疡病发生的重要气象因子还包括相对湿度和降水量。细菌菌落生长要求相对湿度在80%以上。11月至次年1月是四川省猕猴桃的休眠期,相对湿度偏高不利于猕猴桃深度休眠,有利于猕猴桃溃疡病病菌的侵染;同时相对湿度偏高也有利于猕猴桃溃疡病病菌的传播[15, 17-18]。
本文通过实地调查和查阅文献等方式获得猕猴桃溃疡病菌分布信息和未来气候变化数据,利用MaxEnt模型模拟并预测气候变化情景下四川省猕猴桃溃疡病菌的适生分布,通过GIS软件制图并计算适生区面积和质心位移的变化,旨在为四川省猕猴桃溃疡病风险分析、预测预报和有效防控提供一定的理论依据。
1 材料与方法
1.1 气候数据的来源与处理
本文模拟当前(1950年-2000年)和21世纪4个不同年代2030s(2021年-2040年)、2050s(2041年-2060年)、2070s(2061年-2080年)和2080s(2071年-2090年)5个不同时期四川省猕猴桃溃疡病菌的分布情况。当前和未来的气候数据分别下载自WorldClim和Climate Change, Agriculture and Food Security, CCAFS数据库,分辨率为2.5 arc-minutes。未来气候数据选取的是IPCC第五次评估报告发布的RCP2.6、RCP4.5和RCP8.5情景。氣候数据选择的是预测物种地理分布最常用的19个生物气候变量,本研究通过对比各变量建模的贡献百分比筛选建模变量,比较所选变量的相关系数,消除共线性的影响,最终保留了7个气候变量进行建模[20](表1)。
1.2 猕猴桃溃疡病菌分布位点的搜集与确定
猕猴桃溃疡病在意大利、新西兰、中国和欧洲等主要猕猴桃种植区域均有发生,为模拟该病原的准确分布,除该物种的本地分布数据,还应查询其全球分布数据。本研究采取以下方式搜集分布数据:四川省的分布数据为本课题组2015年-2018年对全省猕猴桃溃疡病发生情况系统调查获得,使用GPS精确记录发病地的地理分布信息。国内其他省份及全球的分布数据通过查询物种分布数据库和检索公开发表的猕猴桃溃疡病菌相关文献获得[21-34]。获得数据中有具体经纬度的则直接应用,无经纬度的则查询全球地理信息集成数据库GeoName获得相关坐标信息。为避免同一网格内出现多个分布点而导致的空间自相关性,对分布数据进行了有效筛选[35]。利用ArcGIS的统计分析功能,计算网格中心与分布点之间的距离,保留1条距离中心最近分布记录,最终获得148个分布点。
1.3 模型的构建和适生等级划分
首先将1.1和1.2的数据导入软件中,分布点中选择75%作为训练数据,25%作为测试数据,用于验证模型。其次选择刀切法(jackknife)衡量环境变量的重要性。其余参数均选择模型的默认值,重复运行10次进行建模并设定输出路径。MaxEnt模型模拟结果为目标物种在每个栅格中出现的可能性,即P值,系统默认等级为10级,根据猕猴桃溃疡病菌在四川的实际发生情况并结合相关文献,利用ArcGIS的“Reclassify”功能对连续分布概率(P)进行重分类,划分方法为:白色代表不适生区,P<0.2;黄色代表低适生区,0.2≤P<0.4;橙色代表中适生区,0.4≤P<0.8;红色代表高适生区,P≥0.8[36-37]。
1.4 模型模拟结果评价
ROC(receiver operating characteristic,ROC)曲线其曲线下面积(area under curve,AUC)值的大小是评估生态位模型准确性的有效方法[38]。评估标准定义为:AUC取值范围为[0.5,0.6)时,定义模型模拟结果为“失败”;AUC取值范围为[0.6,0.7)时,定义模型模拟结果为“较差”;AUC取值范围为[0.7,0.8)时,定义模型模拟结果为“一般”;AUC取值范围为[0.8,0.9)时,定义模型模拟结果为“好”;AUC取值范围为[0.9,1.0]时,定义模型模拟结果为“很好”[39]。
2 结果与分析
2.1 模拟准确性检验
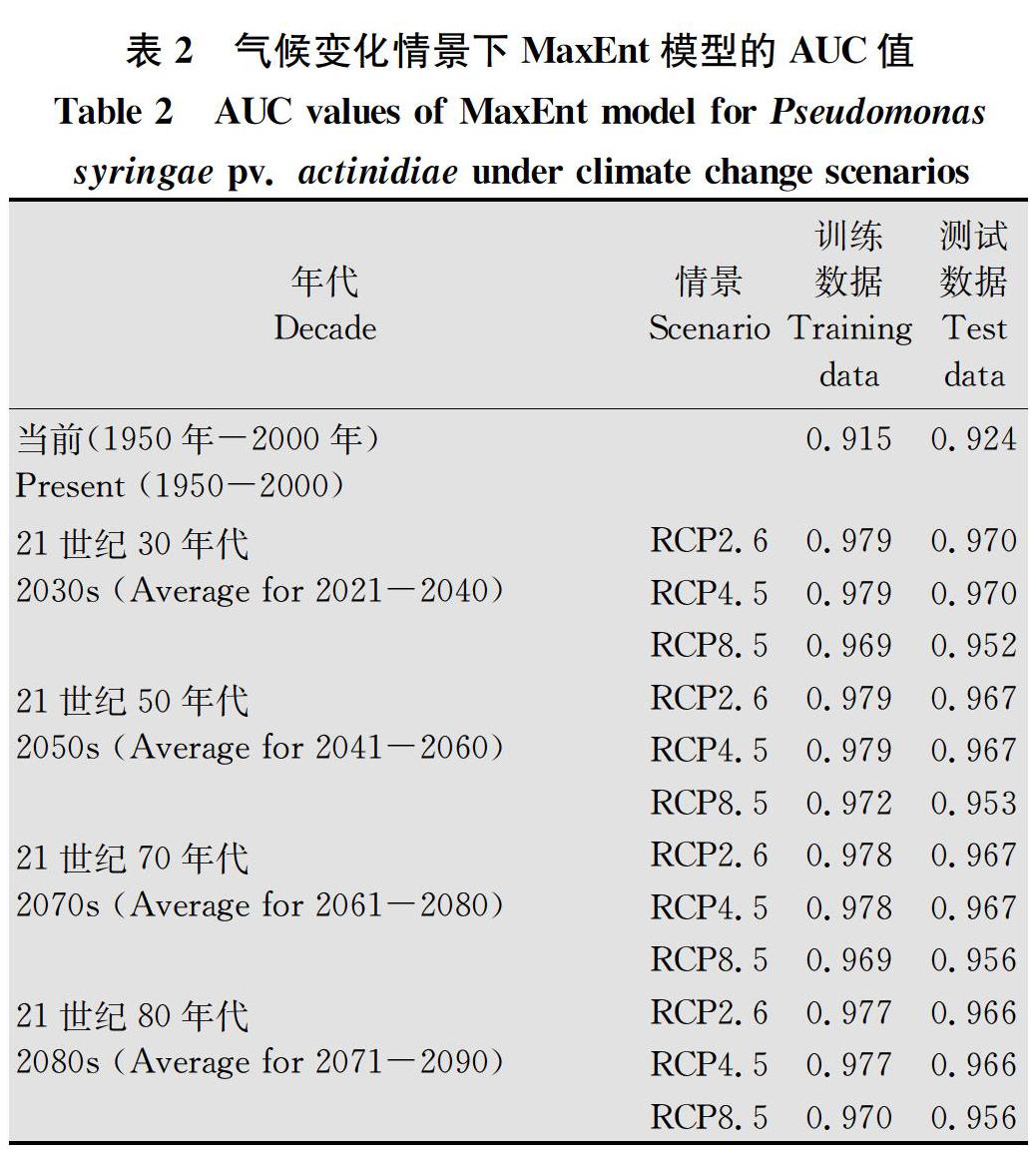
表2是当前及未来气候变化情景下MaxEnt模型训练数据和测试数据的AUC值,结果表明当前气候条件下训练数据和测试数据AUC值分别为0.915和0.924。RCP2.6、RCP4.5和RCP8.5情景下训练数据AUC值分别介于0.977~0.979、0.977~0.979和0.969~0.970,测试数据AUC值则分别介于0.966~0.970、0.966~0.970和0.952~0.956,表明构建的所有模型的模拟结果为“很好”,可用于后续分析。
2.2 当前情景下四川省猕猴桃溃疡病菌地理分布
由图1可以看出,猕猴桃溃疡病菌高适生区位于成都市、德阳市、绵阳市、广元市、巴中市、达州市、雅安市、乐山市、泸州市、宜宾市和广安市,面积达8.4万km2。中适生区在四川21地市(州)均有分布,面积为13.32万km2。低适生区位于甘孜州、阿坝州、凉山州、攀枝花市和达州市,面积为1.18万km2。将猕猴桃溃疡病菌的适生分布图与四川省行政区划图叠加,利用ArcGIS的栅格统计功能计算该病菌在不同地市(州)的适生面积。结果显示,广元市、绵阳市、巴中市、达州市和成都市猕猴桃溃疡病菌的适生面积较大,分别为14 965.28、11 753.47、10 798.61、9 565.97、8 854.17 km2,分别占高适生区总面积的17.81%、13.99%、12.85%、11.38%、10.54%;在中适生区中,凉山州所占面积为38 107.64 km2,占中适生区比例达28.67%(表3)。
2.3 气候变化情景下四川省猕猴桃溃疡病菌地理分布预测 图2是模拟的RCP2.6气候变化情景下,21世纪30年代、50年代、70年代和80年代猕猴桃溃疡病菌在四川的适生分布图。由图2和表4可见,与当前情景相比,高适生区和低适生区区域均显著增加,中适生区区域显著减少。未来高适生区面积由当前的8.41万km2增加到15.91万km2 (2030s)、14.26万km2 (2050s)、14.35万km2 (2070s)、14.97万4 km2 (2080s)。未来中适生区面积由当前的13.32万km2减少至9.39万km2 (2030s)、10.77万km2 (2050s)、9.81万km2 (2070s)、9.69万km2 (2080s)。低适生区面积未来增幅最大,由当前的1.18万km2分别增加至7.85万km2 (2030s)、7.96万km2 (2050s)、5.98万km2 (2070s)和5.95万km2 (2080s)。
图3是RCP4.5气候变化情景下,21世纪30年代、50年代、70年代和80年代猕猴桃溃疡病菌在四川的适生分布图。与当前情景相比,高适生区和低适生区区域均显著增加,中适生区区域显著减少。利用ArcGIS的栅格统计功能计算该病菌适生面积,得到各时期适生区猕猴桃溃疡病菌在四川的分布面积(表4)。
RCP8.5气候变化情景下,21世纪30年代、50年代、70年代和80年代猕猴桃溃疡病菌在四川的潜在分布如图4和表4所示。高适生区面积变化趋势为:由当前的8.41万km2增加到10.64万km2 (2030s)、11.18万km2 (2050s)、13.69萬km2 (2070s)、16.11万km2 (2080s)。中适生区面积变化趋势为:首先由当前的13.32万km2增加至14.18万km2 (2030s)、15.01万km2 (2050s),然后减少至12.69万km2 (2070s)、10.89万km2 (2080s)。低适生区面积未来增幅最大,由当前的1.18万km2分别增加至8.29万km2 (2030s)、8.34万km2 (2050s)、6.79万km2 (2070s)和6.77万km2 (2080s)。
2.4 质心迁移轨迹
为理解不同气候变化情景对猕猴桃溃疡病菌分布的影响,本文参考Yue等[40]的方法计算了不同气候变化情景下适生区质心的位置和迁移规律。由表5可以看出,RCP2.6情景下,中适生区质心由当前位置依次沿西北92.69 km(2030s)、西南16.80 km(2050s)、东北30.53 km(2070s)和西北22.15 km(2080s)移动,至2080s总体上向西北方向移动91.96 km;高适生区质心由当前位置依次沿西南46.07 km(2030s)、西南7.74 km(2050s)、东北15.90 km(2070s)和西北30.08 km(2080s)移动,至2080s总体上向西北方向移动59.02 km。
RCP4.5情景下,高适生区质心由当前位置依次沿西南74.95 km(2030s)、西北4.36 km(2050s)、西北24.05 km(2070s)和东北34.60 km(2080s)移动,至2080s向西北方向移动了62.3 km。中适生区质心由当前位置依次沿西北87.12 km(2030s)、东北24.38 km(2050s)、西北28.66 km(2070s)和东北15.47 km(2080s)移动,至2080s总体上向西北方向移动108.77 km。
RCP8.5情景下,高适生区质心由当前位置依次沿西南57.05 km(2030s)、东南8.00 km(2050s)、西南59.13 km(2070s)和东北57.11 km(2080s)移动,至2080s总体上向西北方向移动66.59 km。中适生区质心由当前位置依次沿西北62.51 km(2030s)、西北25.32 km(2050s)、西北17.83 km(2070s)和东北11.77 km(2080s)移动,至2080s总体上向西北方向移动103.13 km。由此可见,未来不同适生区几何中心位置和迁移规律均有所不同。
3 讨论
本研究选取目前应用较为成熟的最大熵模型构建猕猴桃溃疡病菌模型,判断该病菌在四川的生态位需求,预测未来的地理分布。研究表明,MaxEnt模型较其他常用生态位模型(CLIMEX、GARP、BIOCLIM、DOMAIN等)操作简单、运算效率高,且不受样本量大小限制,并有大量模拟植物病菌潜在分布研究[41-43]。对生态位模型预测效果的评估多采用ROC曲线下面积AUC进行,如韩阳阳等[44]利用ROC曲线测定生态位模型预测松材线虫在中国适生区的准确性。Wang等[45]用ROC曲线评价了MaxEnt模型对稻水象甲在全球适生区预测的效果。本文中所有预测模型的AUC值均达到极好水平,说明模型拟合能力出色。选择环境变量过程中应注意避免变量自相关性,本研究比较各变量对建模的百分比贡献率筛选建模变量,并利用Pearson相关系数法消除共线性的影响,获得了7个变量进行建模,提高了预测的准确性。
本文利用MaxEnt模型,结合前期田间调查和前人研究成果对当前气候条件下四川省猕猴桃溃疡病菌潜在分布区进行预测划分适宜等级,利用ArcGIS软件计算猕猴桃溃疡病菌在各等级的适生面积。结果显示:当前气候条件下,成都市、德阳市、绵阳市、广元市、巴中市、达州市和雅安市为该病菌在四川的高适生区。适生区域(中适生区和高适生区)除甘孜州和阿坝州外,其他19个市(州)均有分布。2017年,猕猴桃溃疡病在四川14个市(州)发生,其中雅安市、成都市和广元市发病面积最大,分别占全省发生面积的43%、37%和7%。对比此次预测结果和猕猴桃溃疡病菌在四川的发生现状可知,除目前已知该病菌广泛分布的成都、雅安、广元等地以外,巴中、达州、广安等地也为该病菌适生等级较高的地区。由此推测,四川省猕猴桃溃疡病仍存在继续扩散的可能。上述高适生区中,已发现该病菌的地区必须及时采取相应措施进行防治,阻止其向其他地区扩散蔓延;对于具备该病菌适生的寄主植物和气候条件的潜在适生分布区,应高度重视,加强检验检疫工作,防止该病菌的传入。调查过程中发现,猕猴桃溃疡病虽为全国森林植物检疫对象,但四川省猕猴桃的种植、管理、推广和病虫害防治等工作由农业部门负责,对于该病害的检验检疫存在一定盲区,造成监管存在漏洞,导致带菌苗木和花粉的传播。我们建议农林部门应加强沟通合作,建立行之有效的监管措施;猕猴桃溃疡病自1986年在中国湖南被发现以来,在长期的扩散传播过程中产生了较高的种内遗传多样性[18],且随着全球气候的变暖,当前的低适生区或不适生区有可能变为该病原菌的适生区,因此对于非适生区,仍应保持高度警惕。邵宝林等[46]采用模糊数学法分析了猕猴桃溃疡病菌在中国的适生性,结果表明该病菌在中国最适宜区包括四川省,但适生面积远小于本研究结果,原因主要是所选模型的预测机理不同所造成。
四川盆地气候特殊,该区域农业病虫害受冬季温度、降水量和日照时数的影响较大[47]。陈超等[48]分析了1961年-2009年四川省气候资源变化趋势,结果表明亚热带向西移动明显,全省热量资源增加趋势明显,大部分地区日照时数减少,该区域气候的变化利于病虫害的发生发展。为研究未来气候变化情景下四川省猕猴桃溃疡病菌分布情况,本文选取了IPCC第五次评估报告公布的RCP2.6情景、RCP4.5情景和RCP8.5情景,2030年代、2050年代、2070年代和2080年代四个未来时段进行分析,定量描述了适生区的变化。RCP2.6情景下,全球范围内促进能源结构改变,大力推广生物能源,生态环境得到巨大改善,温室气体排放最低,全球平均温度上升最少;RCP8.5情景下温室气体排放量大,缺乏应对气候变暖的有效措施,技术革新率低,导致全球能源需求最高,全球平均温度上升最多,生态环境改善甚微;RCP4.5情景为中等稳定排放情景[45]。研究结果表明与当前情景相对比发现,三种气候情景下高适生区和低适生区区域面积均显著增加,中、高适生区质心总体上均向西北方向移动。说明未来气候变暖将使病菌适生范围扩大,更利于病菌越冬和生长繁殖。因此未来四川盆地具备猕猴桃溃疡病大暴发的气候条件,防控任务依旧艰巨。
猕猴桃为猕猴桃溃疡病菌的唯一寄主,不同品种对猕猴桃溃疡病的抗性存在显著性差异,其分布很大程度上取决于寄主品种的地理分布[50-52]。国内外学者研究表明,美味系列猕猴桃品种对猕猴桃溃疡病抗性高于中华系列。目前四川省猕猴桃主栽品种为以‘红阳为代表的中华系列,占总面积比例达83.5%,且有不断增加趋势。张杰等[53]模拟了中华猕猴桃在中国潜在分布及气候变化下潜在擴散地理范围,结果表明,在四川高适生区面积将由当前的13.74万km2增加至2050s的16.49万km2。本文中模拟的当前猕猴桃溃疡病菌高适生区面积为8.41万km2,2050s的适生面积范围为11.18万km2~14.93万km2,表明随着气候变化,猕猴桃溃疡病菌的适生范围有随寄主不断扩大的可能性,特别是二者重合区域在引扩种此类品种时应更为重视不同抗性品种布局。张丽芳等[54]利用遥感技术从地形、气候、土壤、植被等4个方面分析了苍溪红心猕猴桃在四川的适宜性,结果可靠,为苍溪猕猴桃在四川省内引扩种提供了依据。本文仅从气候适宜性角度预测了该病菌在四川的适生分布,预测结果具有一定的局限性。未来工作中,应利用多种手段研究四川省主栽猕猴桃品种的适生性,综合分析寄主-病原物之间的关系,从而更加准确地预测不同气候变化情景下该病菌的潜在分布,为该病菌的有效监测和猕猴桃的种植规划提供理论依据。
参考文献
[1] 茹婧.四川猕猴桃产业发展前景及新商业模式研究[D].北京:北京交通大学,2018.
[2] 涂美艳,黄昌学,陈栋,等.四川猕猴桃产区溃疡病综合防治月历表[J].四川农业科技,2018(1):31-33.
[3] 马利,尹勇,封传红,等.四川省猕猴桃溃疡病发生现状及绿色防控技术体系的建立[J].中国植保导刊,2017,37(11):80-83.
[4] 张花龙,杨念婉,李有志,等.气候变暖对农业害虫及其天敌的影响[J].植物保护,2015,41(2):5-15.
[5] 李培先,林峻,麦迪·库尔曼,等.气候变化对新疆意大利蝗潜在分布的影响[J].植物保护,2017,43(3):90-96.
[6] 蔡静芸,张明明,粟海军,等.生态位模型在物种生境选择中的应用研究[J].经济动物学报,2014,18(1):47-52.
[7] 唐继洪,程云霞,罗礼智,等.基于Maxent模型的不同气候变化情景下我国草地螟越冬区预测[J].生态学报,2017,37(14):4852-4863.
[8] 刘静远,陈林,宋绍祎,等.基于MAXENT的维氏粒线虫(Anguina wevelli)在我国的潜在分布研究[J].植物保护,2016,42(6):86-89.
[9] 齊国君,陈婷,何自福,等.基于Maxent模型的棉花曲叶病在中国的适生性分析[J].棉花学报,2016,28(5):443-451.
[10] 常志隆,周益林,赵遵田,等.基于MaxEnt模型的小麦印度腥黑穗病在中国的适生性分析[J].植物保护,2010,36(3):110-112.
[11] 刘瑶,朱天辉,樊芳冰,等.四川猕猴桃溃疡病的发生与病原研究[J].湖北农业科学,2013,52(20):4937-4942.
[12] 任茂琼,余敖,李家慧,等.四川省北川县猕猴桃溃疡病发生特点与综合防控技术示范[J].中国植保导刊,2018,38(11):55-57.
[13] 王丽,周增强,侯珲,等.我国猕猴桃细菌性溃疡病研究分析及防控[J].中国南方果树,2017(2):178-182.
[14] 潘慧,胡秋舲,张胜菊,等.贵州六盘水市猕猴桃病害调查及病原鉴定[J].植物保护,2018,44(4):125-131.
[15] 龙云川,彭熙,李苇洁,等.贵州猕猴桃溃疡病病菌分离鉴定及其培养特性[J].江苏农业科学,2017,45(20):122-125.
[16] SERIZAWA S, ICHIKAWA T, TAKIKAWA Y, et al. Occurrence of bacterial canker of kiwifruit in Japan: description of symptoms, isolation of the pathogen and screening of bactericides [J]. Japanese Journal of Phytopathology, 1989, 55(4): 427-436.
[17] 高小宁,赵志博,黄其玲,等.猕猴桃细菌性溃疡病研究进展[J].果树学报,2012,29(2):262-268.
[18] 李黎,钟彩虹,李大卫,等.猕猴桃细菌性溃疡病的研究进展[J].华中农业大学学报,2013,32(5):124-133.
[19] 徐晓,肖天贵,麻素红.西南地区气候季节划分及特征分析[J].高原山地气象研究,2010,30(1):35-40.
[20] 张天蛟,刘刚.提高生态位模型时间转移能力的方法研究[J].中国农业大学学报,2017,22(2):98-105.
[21] 张慧琴,李和孟,冯健君,等.浙江省猕猴桃溃疡病发病现状调查及影响因子分析[J].浙江农业学报,2013,25(4):832-835.
[22] 周大祥,殷幼平,王中康,等.利用EMA-qPCR建立快速检测猕猴桃溃疡病菌活菌的方法[J].植物保护,2017,43(3):143-148.
[23] 李瑶,承河元,方书苗,等.猕猴桃细菌性溃疡病流行预测初探[J].应用生态学报,2001,12(3):355-358.
[24] FERRANTE P, TAKIKAWA Y, SCORTICHINI M. Pseudomonas syringae pv. actinidiae strains isolated from past and current epidemics to Actinidia spp. reveal a diverse population structure of the pathogen [J]. European Journal of Plant Pathology, 2015, 142(4): 677-689.
[25] KOH Y J, NOU I S. DNA markers for identification of Pseudomonas syringae pv. actinidiae [J]. Molecules and Cells, 2002, 13(2): 309-314.
[26] FUJIKAWA T, SAWADA H. Genome analysis of the kiwifruit canker pathogen Pseudomonas syringae pv. actinidiae biovar 5 [J/OL]. Scientific Reports, 2016, 6: 21399.
[27] HAN H S, KOH Y J, HUR J W, et al. Identification and characterization of coronatine-producing Pseudomonas syringae pv. actinidiae [J]. Journal of Microbiology and Biotechnologygy, 2003, 13(1): 110-118.
[28] KOH Y J, KIM G H, JUNG J S, et al. Outbreak of bacterial canker on Hort16A (Actinidia chinensis Planchon) caused by Pseudomonas syringae pv. actinidiae in Korea [J]. New Zealand Journal of Crop and Horticultural Science, 2010, 38(4): 275-282.
[29] CUNTY A, CESBRON S, POLIAKOFF F, et al. Origin of the outbreak in France of Pseudomonas syringae pv. actinidiae Biovar 3, the causal agent of bacterial canker of kiwifruit, revealed by a multilocus variable-number tandem-repeat analysis[J]. Applied and Environmental Microbiology, 2015, 81(19): 6773-6789.
[30] RENZI M, MAZZAGLIA A, BALESTRA G M. Widespread distribution of kiwifruit bacterial canker caused by the European Pseudomonas syringae pv. actinidiae genotype in the main production areas of Portugal [J]. Phytopathologia Mediterranea, 2012, 51(2): 402 -409.
[31] BUTLER M I, STOCKWELL P A, BLACK M A, et al. Pseudomonas syringae pv. actinidiae from recent outbreaks of kiwifruit bacterial canker belong to different clones that originated in China [J/OL]. PLoS ONE, 2013, 8(2): e57464.
[32] VANNESTE J L, YU J, CORNISH D A, et al. Identification, virulence, and distribution of two biovars of Pseudomonas syringae pv. actinidiae in New Zealand [J]. Plant Disease, 2013, 97(6): 708-719.
[33] ABELLEIRA A, ARES A, AGUN O, et al. Current situation and characterization of Pseudomonas syringae pv. actinidiae on kiwifruit in Galicia (northwest Spain)[J]. Plant Pathology, 2014, 63(3): 691-699.
[34] PRENCIPE S, NARI L, VITTONE G, et al. Effect of bacterial canker caused by Pseudomonas syringae pv. actinidiae on postharvest quality and rots of kiwifruit ‘Hayward [J]. Postharvest Biology and Technology, 2016, 113: 119-124.
[35] 張东方,张琴,郭杰,等.基于MaxEnt模型的当归全球生态适宜区和生态特征研究[J].生态学报,2017,37(15):5111-5120.
[36] 叶永昌, 周广胜, 殷晓洁. 1961-2010年内蒙古草原植被分布和生产力变化——基于MaxEnt模型和综合模型的模拟分析[J]. 生态学报, 2016, 36(15): 4718-4728.
[37] 孙颖, 秦大河, 刘洪滨. IPCC第五次评估报告不确定性处理方法的介绍[J]. 气候变化研究进展, 2012, 8(2): 150-153.
[38] 王雷宏, 杨俊仙, 徐小牛. 基于MaxEnt分析金钱松适生的生物气候特征[J]. 林业科学, 2015, 51(1): 127-131.
[39] 王茹琳, 李庆, 封传红, 等. 基于MaxEnt的西藏飞蝗在中国的适生区预测[J]. 生态学报, 2017, 37(24): 8556-8566.
[40] YUE T X, FAN Z M, CHEN C F, et al. Surface modelling of global terrestrial ecosystems under three climate change scenarios [J]. Ecological Modelling, 2010, 222(14): 2342-2461.
[41] ELITH J, GRAHAM C, ANDERSON R, et al. Novel methods improve prediction of species distributions from occurrence data [J]. Ecography, 2006, 29(2): 129-151.
[42] PETITPIERRE B, KUEFFER C, BROENNIMANN O, et al. Climatic niche shifts are rare among terrestrial plant invaders[J]. Science, 2012, 335(6074): 1344-1348.
[43] 张海涛,罗渡,牟希东,等.应用多个生态位模型预测福寿螺在中国的潜在适生区[J].应用生态学报,2016,27(4):1277-1284.
[44] 韩阳阳,王焱,项杨,等.基于Maxent生态位模型的松材线虫在中国的适生区预测分析[J].南京林业大学学报(自然科学版),2015,39(1):6-10.
[45] WANG Cong, HAWTHORNE D, QIN Yujia, et al. Impact of climate and host availability on future distribution of Colorado potato beetle [J/OL]. Scientific Reports, 2017, 7(1): 4489.
[46] 邵宝林,王成华,张婧,等.猕猴桃溃疡病菌在中国的适生性分析[J].植物保护,2016,42(2):146-150.
[47] 孙华,何茂萍,胡明成.全球变化背景下气候变暖对中国农业生产的影响[J].中国农业资源与区划,2015,36(7):51-57.
[48] 陈超,庞艳梅,潘学标,等.气候变化背景下四川省气候资源变化趋势分析[J].资源科学,2011,33(7):1310-1316.
[49] 沈永平,王国亚.IPCC第一工作组第五次评估报告对全球气候变化认知的最新科学要点[J].冰川冻土,2013,35(5):1068-1076.
[50] 李淼,檀根甲,李瑶,等.猕猴桃品种酚类物质及可溶性蛋白含量与抗溃疡病的关系[J].植物保护,2009,35(1):37-41.
[51] 石志军,张慧琴,肖金平,等.不同猕猴桃品种对溃疡病抗性的评价[J].浙江农业学报,2014,26(3):752-759.
[52] 张慧琴,毛雪琴,肖金平,等.猕猴桃溃疡病病原菌分子鉴定与抗性材料初选[J].核农学报,2014,28(7):1181-1187.
[53] 张杰,敖子强,吴永明,等.中华猕猴桃(Actinidia chinensis)在中国的适生性及其潜在地理分布模拟预测[J].热带地理,2017,37(2):218-225.
[54] 张丽芳,张鑫,杨存建,等.四川省引扩种苍溪红心猕猴桃的适宜性研究[J].信阳师范学院学报(自然科学版),2018,31(3):70-76.
(责任编辑: 田 喆)
猜你喜欢
财会月刊·上半月(2022年5期)2022-05-17
睿士(2021年9期)2021-09-15
疯狂英语·新阅版(2021年1期)2021-03-03
环球时报(2021-01-28)2021-01-28
环球时报(2019-12-02)2019-12-02
环球时报(2019-09-24)2019-09-24
商界(2019年12期)2019-01-03
健康管理(2015年12期)2016-01-14
环球时报(2015-12-16)2015-12-16
