
巴东木莲幼苗对干旱胁迫的生长和生理响应
2020-06-03 04:37王毅敏谢春培王玉兵李薇高晗雷华
湖北林业科技 2020年2期
王毅敏 谢春培 王玉兵 李薇 高晗 雷华

摘 要:采用盆栽人工模拟控水法干旱胁迫,探讨巴东木莲Manglietia patungensis幼苗的生长和生理特性对干旱胁迫的响应情况,随着干旱胁迫的加剧,巴东木莲幼苗苗高、地径和生物量逐渐降低,可溶性蛋白含量逐渐升高,叶绿素含量在对照处理略高,在轻度胁迫处理和中度胁迫处理明显降低。随着胁迫时间的延长,巴东木莲幼苗叶片组织相对含水量降低,丙二醛含量先升后降再升,脯氨酸含量逐渐升高,保护酶SOD活性均呈先增加而后降低的变化趋势。研究表明:在不同程度的干旱胁迫条件下,巴东木莲幼苗的生长受到一定程度的抑制,但能够通过调节自身的保护酶系统活性和渗透调节物质含量来减轻干旱伤害,维持植物体的正常生理代谢功能,从而表现出一定的抗旱耐旱潜力。
关键词:巴东木莲幼苗;干旱胁迫;生理生化
中图分类号:Q945.78 文献标识码:A 文章编号:1004-3020(2020)02-0006-05
Growth and Physiological Response of Manglietia patungensis Seedlings to Drought Stress
Wang Yimin(1,2) Xie Chunpei(1) Wang Yubing(1) Li Wei(2) Gao Han(2) Lei Hua(2)
(1.Biotechnology Research Center of China Three Gorges University Yichang 443001;2.Management Office of Three Gorges Botanical Garden Yichang 443111)
Abstract:Drought stress by artificial simulated water control in potted plants,To investigate the response of the growth and physiological characteristics of Manglietia patungensis seedlings to drought stress .With the increase of drought stress,the seedling height,ground diameter and biomass of M. patungensis seedlings decreased gradually,the soluble protein content increased gradually,the chlorophyll content was slightly higher in the control treatment,and the treatment under mild stress and moderate stress was significantly reduced. With the prolongation of stress time,the relative water content of the leaves of M. patungensis seedlings decreased,the content of malondialdehyde increased first,then decreased,the content of proline increased gradually,The protective enzyme SOD activity increased first and then decreased. The research shows that under different degrees of drought stress,the growth of M. patungensis seedlings is inhibited to a certain extent,but it can reduce the drought damage by regulating the activity of its protective enzyme system and the content of osmotic regulatory substances. Maintain the normal physiological metabolic function of the plant body,thus showing a certain drought tolerance potential.
Key words:Manglietia patungensis seedlings;drought stress;physiological and biochemica
巴东木莲Manglietia patungensis为木兰科Magnoliaceae木莲属的常绿阔叶树种,零星分布于湖北、湖南、江西、重庆等地的局部地区[1-2]。巴东木莲不仅树形优美、花大美丽,具有很高的观赏价值;而且該物种是木莲属分布最北的种,在木兰科植物系统发育与演化上具有重要的研究价值[3]。现有资料表明,巴东木莲因种子萌发力差,林下幼苗及幼树极为罕见,天然更新困难而处于濒危状态。目前,巴东木莲的相关研究主要集中于分类学、生物学特性[4]、群落学[5]、保护遗传学[6]、种子生物学[7]、繁殖生物学[8-9]、组织培养[10]等方面,而与巴东木莲抗旱性的相关研究还尚未见报道。植物在生长发育过程中总是不可避免的经受着干旱等逆境的危害,特别是在幼苗向幼树的过渡阶段,其更容易受干旱胁迫的影响,一旦此阶段出现过度干旱,其后果是致命的。为了更好的保护和利用巴东木莲,对其幼苗进行干旱胁迫研究显得至关重要,其研究结果可为该物种的保育提供一定的科学依据,同时对巴东木莲的种群恢复具有十分重要意义。
1 材料与方法
1.1 试验材料
以湖南省小溪国家级自然保护区收集的巴东木莲种子为试验材料,2月底播种,待种子萌发后具一片真叶时,筛选生长基本一致且无病虫害的芽苗栽种于直径8 cm,高12 cm的营养钵中,每盆装腐殖土0.8 kg,置于温室培养,确保合适栽培条件,待芽苗缓苗至具3片真叶时,选取长势基本一致的幼苗进行控水处理。
1.2 试验设计
采用盆栽人工模拟控水法干旱胁迫,共设置对照组(CK)、轻度胁迫(T1)、中度胁迫(T2)和重度胁迫(T3),其土壤含水量分别为:土壤含水量为土壤田间最大持水量的90%~85%、75%~70%、55%~50%和35%~30%。每个处理重复5次,每个重复30株。于7月1日开始采用称重法进行控水试验,连续控水50 d。每隔10 d的早上8点左右分别取第3~5叶成熟叶片,立即带回实验室采用液氮处理后置于-70℃低温冰箱备用。
1.3 指标测定方法
1.3.1 生长指标测定
利用卷尺和游标卡尺每隔10 d,随机选取30株测定苗高和地径,计算平均值。
1.3.2 生理生长指标测定
丙二醛(MDA)测定采用硫代巴比妥酸法[11];叶绿素含量测定采用紫外分光光度计比色法[12];游离脯氨酸(Pro)含量测定采用磺基水杨酸提取茚三酮显色法[13];超氧化物歧化酶(SOD)活性测定采用NBT还原法[14];可溶性蛋白含量测定采用考马斯亮蓝法[15]。
1.4 数据分析
采用EXCEL2007和SRSS17.0软件对数据进行分析。
2 结果与分析
2.1 干旱胁迫对巴东木莲幼苗生长的影响
干旱胁迫对巴东木莲幼苗的生长存在着一定的抑制影响,从图1、2中的可以看对照组的平均地径和苗高均比其它试验组要高,重度胁迫组的地径和苗高最低,且巴东木莲幼苗株高在重度胁迫和中度胁迫下普遍比轻度胁迫和对照组的低,说明随着干旱胁迫程度加重巴东木莲幼苗的地径和株高生长受到的影响也较大。
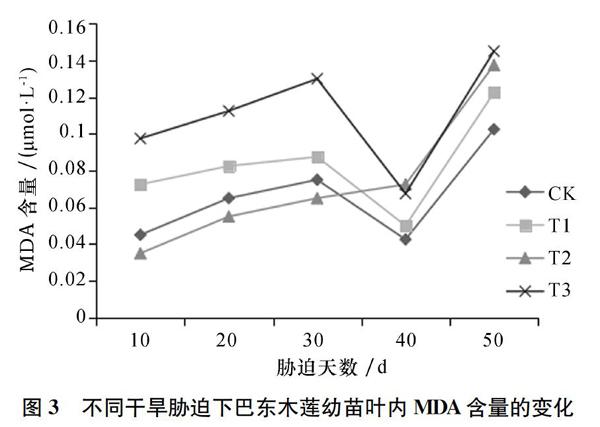
2.2 巴东木莲幼苗叶片内丙二醛对干旱胁迫的响应
丙二醛(MDA)是膜脂过氧化物的最终产物,反映了植物体内自由基一定程度上的活性情况。在对照和各胁迫处理下,巴东木莲幼苗叶片中MDA含量的变化趋势相似,随胁迫的持续,各处理中MDA含量如图3所示,呈现出先升高出现一个峰值,降低后再上升的现象。30 d时,重度胁迫的样品中MDA的含量首先达到峰值,比其他处理组的含量都高,此时重度胁迫下的巴东木莲幼苗内自由基活性最激烈;40 d时,中度胁迫处理的MDA含量比其他处理组的都高,说明中度胁迫40 d是巴东木莲幼苗承受的一个节点。随着胁迫时间的持续延长,各处理中MDA含量都上升,这说明巴东木莲幼苗的叶片是通过膜脂过氧化作用,增加膜的通透性来响应干旱胁迫的。
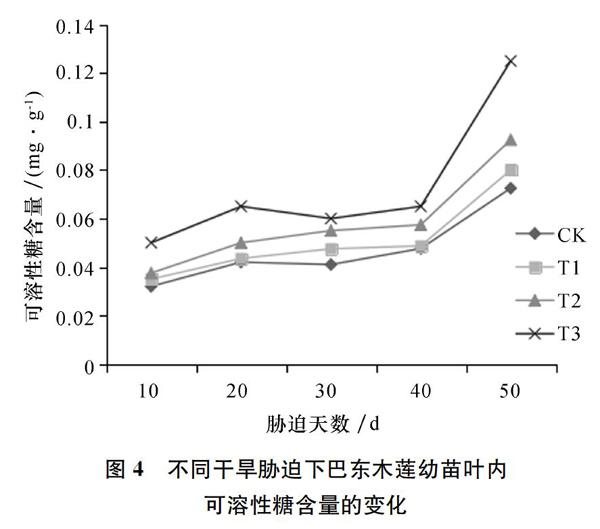
2.3 巴东木莲幼苗叶片内可溶性糖含量变化对干旱胁迫的响应
随着干旱胁迫程度的增加,如图4所示巴东木莲幼苗样本中可溶性糖的含量变化明显,逐渐升高,且胁迫时间越长,可溶性糖含量变化越大。重度胁迫处理样品内可容性糖的含量明显高于其它处理。可见,可溶性糖对抵御干旱胁迫有重要的调节作用,可溶性糖含量的变化可以反映巴东木莲幼苗对外界环境條件一定程度上的响应情况。
2.4 巴东木莲幼苗叶片内游离脯氨酸(Pro)对干旱胁迫的响应
随着干旱胁迫程度的增加,如图5所示巴东木莲幼苗样品中的脯氨酸(Pro)含量也相应增加,在同一处理水平,随着胁迫时间的延长,Pro的含量呈现先增加后微减的趋势。重度胁迫下尤其明显,Pro的含量在重度胁迫的第40天达到最高。巴东木莲幼苗遭遇旱胁迫时会通过增加脯氨酸的含量来抵抗干旱逆境。
2.5 巴东木莲幼苗叶内内SOD酶对干旱胁迫的响应
超氧化物歧化酶(SOD)是一种可以清除超氧阴离子自由基的酶。由图5可以看出对照组中SOD酶的活性变化幅度相对比较小。在胁迫期间无论是对照组还是胁迫组,SOD酶的活性都呈现先上升后下降的趋势。在处理30 d时SOD酶活性到最高,也就是说此时SOD酶在清除活性氧方面起到重要的作用。还发现干旱处理的后期SOD酶含量出现下降的趋势,但还是高于对照组水平。这可能是长时间干旱胁迫将会使巴东木莲幼苗内水分含量减少,活性减弱,抗氧化能力也逐渐减弱,幼苗出现虚弱状况,SOD酶仍具有活性,说明巴东木莲幼苗对长期胁迫的响应需要SOD酶调节。
2.6 巴东木莲幼苗叶片中叶绿素对干旱胁迫的响应
由图7可以看出对照组和轻度胁迫两个处理中叶绿素含量的变化相对比较明显,重度胁迫和中度胁迫处理中叶绿素的含量变化不是很大,而且都低于对照组。可见,水分的多叶绿素的含量也多,随干旱胁迫的加强和时间的延长,叶绿素含量变化变小,表明巴东木莲幼苗对干旱胁迫的响应,可以通过叶绿素含量降低,减弱光合作用程度,减少产生营养物质的量来调节,但是持续时间长会影响植物生长。
2.7 巴东木莲幼苗叶片中可溶性蛋白对干旱胁迫的响应
随着胁迫程度的增加,如图8所示巴东木莲幼苗叶片中可溶性蛋白的含量缓慢升高。在各胁迫处理下,可溶性蛋白的含量都高于对照组,且在重度胁迫时,可溶性蛋白的含量显著高于其他处理。表明巴东木莲幼苗在干旱期间可能会通过增加可溶性蛋白的含量来抵制干旱条件,满足自身的生长,也说明在干旱条件下,可溶性蛋白对植物抵制不良环境有重要作用。
3 讨论
3.1 巴东木莲幼苗生长指标变化
生长量是评估植物抗性的明显指标。通过试验数据发现,干旱胁迫会对巴东木莲幼苗的生长产生一些程度的影响,在重度干旱胁迫下株高和地径都受到显著的影响。但也并没有出现死亡的现象,可见巴东木莲幼苗会通过调整自身内部生物量和生长量来适应干旱逆境。植物在遇到逆境时,会积累一些可溶性糖,来抵御外界环境条件带来的不适应[16]。巴东木莲幼苗也可以通过增加可溶性糖的含量增强其抵抗干旱环境的能力。有研究认为,干旱胁迫会使杨树中叶绿素含量下降,那是因为干旱时植物可利用的水分减少,植物组织内水分会出现亏缺,蒸腾作用会减弱,光合作用也会下降[17-18]。本研究表明,在胁迫处理期间,叶绿素的含量是先增高后降低,先增高的原因是幼苗调节内部新陈代谢活动响应干旱胁迫,减少能量的消耗,叶面积减小,相对光合速率提高生理性的调节响应逆境。后来随着胁迫程度的增加叶绿素含量降低,原因可能是随干旱胁迫的持续,水分减少,叶片细胞内膜脂过氧化反应,丙二醛含量增加,活性氧对叶绿素造成破坏。
3.2 丙二醛和SOD酶对干旱胁迫的响应
植物在逆境条件下,体内会发生脂膜过氧化物作用,以往许多研究都将丙二醛的量来指示细胞质过氧化程度和植物对逆境抵抗的强弱[19]。通过研究发现在水分处理初期,巴东木莲幼苗中MDA的含量有缓慢增加的趋势,随着干旱胁迫时间的延长,MDA的含量出现一个峰值后又下降,表明巴东木莲幼苗可通过膜脂过氧化作用来响应干旱逆境胁迫,胁迫中期通过保护酶清除活性氧来响应,在胁迫后期,MDA含量降低,是由于巴东木莲幼苗内各种酶对的胁迫响应、抗旱锻炼,巴东木莲幼苗可能有了一定的适应,胁迫处理组样品中MDA含量比对照组高,说明MDA对干旱胁迫的响应效果较明显,幼苗受伤害的程度不明显,这和红花玉兰幼苗受干旱胁迫的研究情况一致[20]。干旱胁迫下,不同植物的保护酶系统的防御能力不同,抗性强弱依靠各种酶的协调情况。SOD就是细胞抵御活性氧伤害的酶,可以提高植物适应干旱胁迫的能力[21]。从本研究可以看出,随着干旱胁迫的加剧,巴东木莲幼苗内SOD酶的活性也变强,以此增强保护酶活性可以清除活性氧,减少活性氧积累,抵御干旱逆境造成的伤害。
3.3 脯氨酸和可溶性蛋白含量对干旱胁迫的响应
脯氨酸亲水性较强,是植物体内重要的渗透调节物质,可以作为抗旱育种的生理指标[22]。本研究中随着干旱程度的增加,脯氨酸的含量明显增加,说明巴东木莲幼苗可通过增加脯氨酸来响应干旱胁迫,抵抗干旱的环境带来的不适应,提高渗透调节能力,防止细胞内水分过多的丢失。可溶性蛋白具有较强的亲水性,巴东木莲幼苗在干旱胁迫时,可溶性蛋白主动积累,调节提高细胞的保水力,降低组织的渗透势,响应干旱胁迫带来的伤害。随着干旱胁迫的加重,巴东木莲幼苗叶片中可溶性蛋白含量会降低,是由于对干旱胁迫的响应机制,组织内蛋白酶的活性升高,使可溶性蛋白水解,也可能是RNA转录和翻译因干旱胁迫不能正常进行,从而使蛋白质合成受阻[23]。本研中巴东木莲幼苗可溶性蛋白含量在轻度胁迫、中度胁迫和重度胁迫程度都比对照组的高,可能是因巴东木莲幼苗为响应干旱胁迫,内部有些相关的基因启动表达,导致体内正常的蛋白质合成受阻,导致可溶性蛋白含量变化,这与4个杨树新无性系的抗旱研究结果基本一致[24]。
综上所述,在干旱重度胁迫下巴东木莲幼苗株高和地径都受到显著的影响。但也并没有出现死亡的现象,巴东木莲可通增加可溶性糖的含量、调整MDA的含量先增后减再增,降低SOD酶的活性、增加脯氨酸的含量来调整内部保护酶系统的来适应干旱胁迫。可见巴东木莲幼苗自身具有较好应对干旱胁迫的调节能力,可以为巴东木莲保育和生境与濒危关系研究提供理论依据。
参 考 文 献
[1]傅立国.中国植物红皮书——稀有濒危植物:第一册[M].北京:科学出版社,1992:438.
[2]李晓东,黄宏文,李建强,等.湖南小溪自然保护区巴东木莲群落结构[J].植物科学学报,2006,24(1);31-37.
[3]李晓东,黄宏文,李作洲,等.濒危植物巴东木莲的分布及保护策略[J].武汉植物学研究,2004,22(5):421-427.
[4]桑子阳,张德春,王玉兵,等.19个木莲属树种的花粉形态观察及其分类学意义[J].植物研究,2016,36(1):43-51.
[5]葛刚,陈少风,王孝,等.江西官山自然保护区巴东木莲群落特征[J].亚热带植物科学,2009,38(4):44-49.
[6]何敬胜,李作洲,黄宏文.濒危物种巴东木莲的等位酶遗传多样性及其保护策略[J].生物多样性,2005,13(1):27-35
[7]陈发菊,梁宏伟,王旭,等.濒危植物巴东木莲种子休眠与萌发特性的研究[J].生物多样性,2007,15(5):492-499
[8]宗卫,费永俊,喻慧.巴东木莲实生苗培育及光合生理特征[J].福建林业科技,2013,40(2):56-59+69
[9]李宗艳,郭荣.木莲属濒危植物致瀕原因及繁殖生物学研究进展[J].生命科学研究,2014,18(1):90-94
[10]陈发菊,张丽萍,卢斌,等,长江三峡珍稀植物——巴东木莲冬芽的组织培养[J].生物学通报,2000(6):34-35
[11]高俊凤.植物生理学实验指导[M].北京:高等教育出版社,2006.
[12]郝建军,康宗利,于洋.植物生理学实验技术[M].北京:化学工业出版社,2007.
[13]李绍军,龚月桦,王俊儒,等.关于茚三酮法测定脯氨酸含量中脯氨酸与茚三酮反应之探讨[J].植物生理学通讯,2005(3):365-368
[14]蓝开蔚,黄建鸣,谢之荣,等.超氧化物歧化酶活性测定——Pyrogallol-NBT比色法与化学发光法的比较[J].生物化学与生物物理进展,1988(2):138-140.
[15]曲春香,沈颂东,王雪峰,等.用考马斯亮蓝测定植物粗提液中可溶性蛋白质含量方法的研究[J].苏州大学学报(自然科学版),2006(2):82-85
[16]柯世省,金则新.干旱胁迫对夏腊梅叶片脂质过氧化及抗氧化系统的影响[J].林业科学,2007,43(10):28-33
[17]吴强盛,夏仁学.水分胁迫下丛枝菌根真菌对枳实生苗生长和渗透调节物质含量的影响[J].植物生理与分子生物学学报,2004,30(5):583-588.
[18]段娜,王佳,刘芳,等.植物抗旱性研究进展[J].分子植物育种,2018,16(15):5093-5099
[19]王彦芹,张宁,李志军.苦豆子种子萌发期及幼苗期对盐、旱耐受性的探索[J].分子植物育种,2017,15(3):1090-1095.
[20]桑子阳,马履一,陈发菊.干旱胁迫对红花玉兰幼苗生长和生理特性的影响[J].西北植物学报,2011,31(1):109-115.
[21]R.Alonso,S.Elvira,F.J.Castillo,and B.S.Gimeno.Interactive effects of ozone and drought stress on pigments and activities of antioxidative enzymes in pinus halepensis[J].Plant Cell & Environment,2010,24(9):905-916.
[22]陈亚娟,李付广,刘传亮,等.植物渗透调节研究进展及与棉花耐旱遗传改良[J].分子植物育种,2009,7(1):149-154.
[23]Li X.,Li X.,P.,Chen L.Y.,Wang Y.B.,Li X.L.,and Chen F.J.,Genetic diversity of endangered Manglietia patungensis assessed by inter simple sequence repeat and sequence-related amplified polymorphism markers[J]. Biochemical Systematics and Ecology,2014,57:231-237.
[24]邱兴,吕小锋,李晓东.4个杨树新无性系的抗旱性研究[J].西北林学院学报,2015,30(4):99-108.
(责任编辑:唐 岚)
收稿日期:2019-09-09
基金项目:湖北省技术创新专项资助(NO:2018ACA132);国家自然基金资助(N0:31770654)。
作者简介:王毅敏(1986~),女,硕士研究生,工程师。从事植物多样性保护、繁育,林业生产及林业科研。
王玉兵为通讯作者。
猜你喜欢
农家科技下旬刊(2017年2期)2017-03-27
湖北农业科学(2017年1期)2017-03-09
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
安徽农学通报(2016年24期)2017-01-12
山东农业科学(2016年11期)2016-12-17
科技视界(2016年11期)2016-05-23
