
余甘子天然居群果实的表型多样性分析及综合评价
2020-06-03 02:15王德源朱亚艳戴晓勇
贵州农业科学 2020年4期
王德源, 朱亚艳, 戴晓勇, 余 娜, 许 杰*
(1.贵州省林业调查规划院, 贵州贵阳 550003;2.贵州省林业科学研究院, 贵州 贵阳 550005)
表型多样性是遗传多样性与环境多样性的综合体现,通过表型性状检测遗传变异的方法近年来被广泛应用于多个物种的种质资源遗传多样性研究[1-5]。表型多样性的分析有助于了解物种遗传分化和进化潜力,有利于优异种质的筛选和保护利用[6-7]。
余甘子(PhyllanthusemblicaLinn.)属于大戟科(Euphorbiaceae)叶下珠属(Phyllanthus),是一种生长在我国亚热带、热带部分地区,特别是干热河谷地区的多年生落叶小乔木或灌木,俗名橄榄、滇橄榄、油甘子等[8]。其果实风味独特,富含维生素C、多酚、黄酮、多糖等化合物[9-11],其中Vc含量为0.6%~0.92%,是苹果的160倍,猕猴桃的5倍[12-13],可直接食用或作药用,具有很好的保健作用,是卫生部审核并颁布的药食同源植物,许多营养学家把余甘子果实、猕猴桃、山核桃并列为我国三大高营养果,联合国粮农组织也把其列为有待积极开发的果树,我国广西、福建已经有大面积人工栽培,但野生资源以云南最为丰富[14]。野生余甘子在云南有广泛分布,独特的地理气候条件使其进化积累了丰富的遗传变异,但很多野生资源仍未得到保护利用。开展余甘子天然居群果实表型多样性分析,了解不同居群的表型变异特征和多样性水平,有助于筛选优良资源,对余甘子资源保护与利用具有重要意义。为此,对6个天然居群余甘子果实表型性状进行多样性分析,并进行综合评价,以揭示野生余甘子果实形态在居群间和居群内的分化程度和变异规律,为余甘子种质资源的收集、保护以及优良种质的选择利用提供科学依据。
1材料与方法
1.1试验材料
野生余甘子:6个居群的野生余甘子采自云南省凤庆县,各居群地理位置和样本数见表1。原则上每个居群的样本数均应在10株以上,株距大于30 m,但部分居群由于样本偏少,故采集的样本不足10株。所选单株从东西南北4个方向随机采集树冠中部和中上部的成熟果实进行表型性状测定。
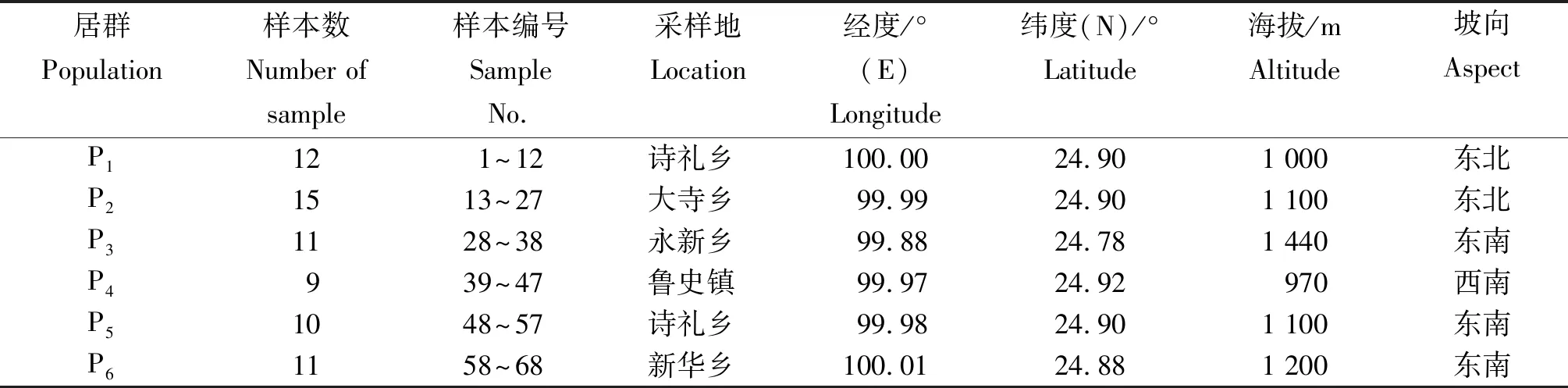
表1 野生余甘子样品的居群地理位置
1.2果实表型性状观测
在余甘子果实成熟期观测表型性状,选取果实形状等6个描述性状和单果重等9个数量性状作为研究指标分析余甘子果实的表型多样性。描述性状采用目测观察法进行等级划分(表2)。数量性状分别采用游标卡尺和电子天平进行测量。每个样株随机采集30个果实,新鲜果实即时用游标卡尺测量果纵径、果横径、外果皮厚、果核纵径、果核横径(精确到0.01 cm),计算果形指数(果纵径/果横径),用电子天平测定单果重、外果皮重、果核重(精确到0.001 g)。

表2 余甘子果实描述性性状的等级划分
1.3数据处理与分析
利用Excel 2007统计描述性状的变异情况,并计算Shannon-Weaver多样性指数(H')[15]。利用SPSS 22.0对数量性状进行标准化后,计算平均值、标准差(SD)和变异系数(CV),并进行相关性分析、主成分分析和聚类分析,用SAS计算表型分化系数(Vst)。
2结果与分析
2.1余甘子果实表型性状多样性
2.1.1描述型性状从表3可知,6个居群68份余甘子材料果实的6个描述性状均有不同程度的变异,多样性指数为0.683 1~1.660 6,平均为1.084 1,其中果实颜色和果实形状多样性指数较大。各性状在其描述级别上均有分布,但不均匀,果实形状有5种表型,以圆形和瓜形为主,分别占总分布频率的29.81%和34.85%;果实颜色有6种表型,翠绿色和黄白色居多,深绿色最少;果基形状有3种表型,以浅凹型所占比例最大;果顶形状有3种表型,以平为主;果腹缝线形状有凹平2种表型,分别占总分布频率的42.94%和57.06%;果核粘离度以易离核为主,占总分布频率的70.74%。

表3 余甘子果实6个描述型性状的频率分布及多样性指数
2.1.2余甘子果实表型性状及变异特征从表4可知,余甘子天然居群果实形态特征在居群间存在一定差异,其中P5居群余甘子果实的平均果横径、果纵径、单果重、单个果肉重、果肉厚度、出肉率均最大,分别为2.50 cm、2.30 cm、7.37 g、6.05 g、0.65 cm和81.18%,果大、可食率高。多重比较结果表明,P5居群的单果重、单个果肉重和出肉率与其余5个居群间差异显著;P1居群果小、可食率较高,其中,果横径、果纵径、单果重、单个果肉重、果肉厚度均最小,分别为2.07 cm、2.00 cm、5.20 g、4.04 g和0.46 cm,但出肉率并不是最小,为75.26%,其单果重、单个果肉重、果肉厚度与其他5个居群间呈显著差异;P4居群果大、可食率低,其中果横径、果纵径、单果重、单果果肉重、果肉厚度均居第二,但出肉率最小,为73.67%,其果核重与其余5个居群间差异显著。
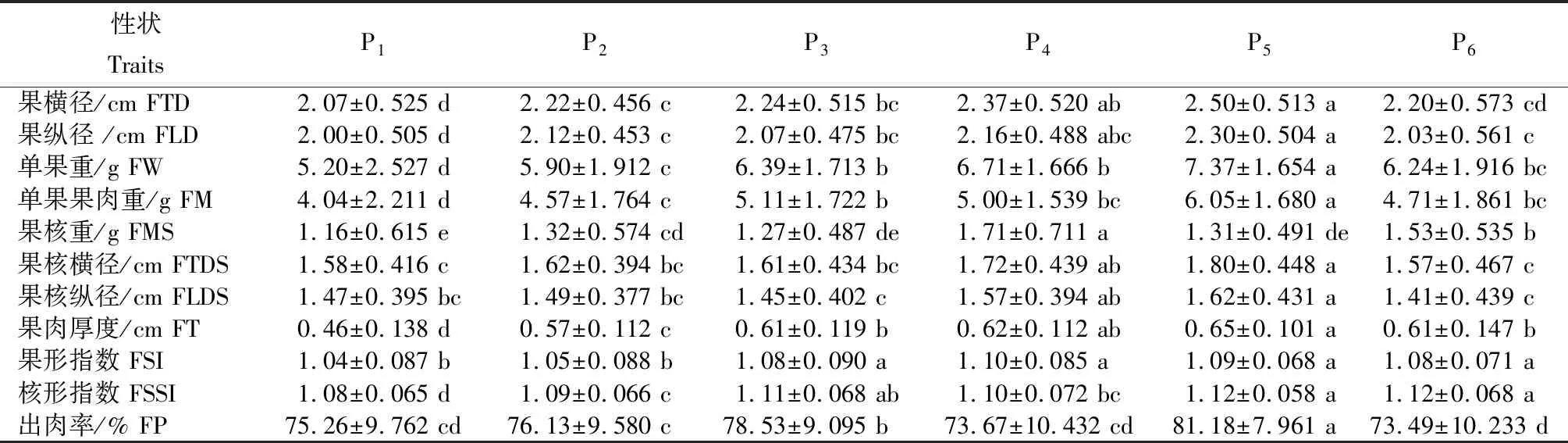
表4 6个居群间表型性状的平均值、标准差及多重比较
注:同行不同小写字母间表示同一性状在不同居群间差异显著(P<0.05)。
Note: Different lowercase letters in the same row indicate significance of difference in the same trait among different populations atP<.05 level.
从表5可知,11个表型性状在居群内不同个体间的平均变异系数(CV)为24.30%,变幅为6.16%~43.35%,其变异程度为果核重>单个果肉重>单果重>果核纵径>果核横径>果纵径>果肉厚度>果横径>出肉率>果形指数>核形指数。果核重、单个果肉重、单果重、果纵径、果横径的变异系数较大,均大于11个表型性状的平均变异系数,变异丰富,稳定性较低;核形指数(6.16%)和果形指数(7.89%)的变异系数较小,具有较高的稳定性。在居群水平上,P1和P6居群的平均变异系数较高,表型变异相对丰富,多样性程度相对较高;P5居群的平均变异系数最小,表型变异程度较低。说明不同性状对环境的适应性和不同生态环境导致了居群变异程度的差异。
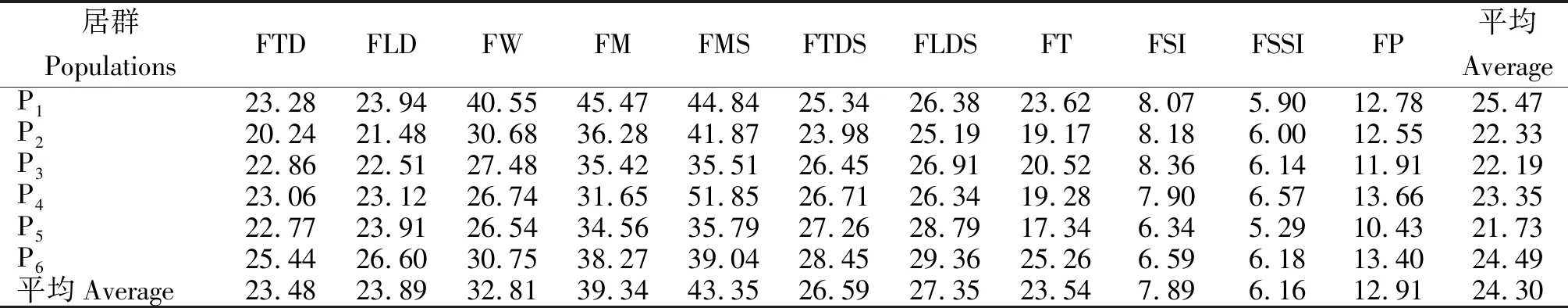
表5 6个居群表型性状的变异系数
2.1.3余甘子居群间果实表型分化特征表型分化系数(Vst)为某性状在居群间方差分量占居群总方差分量的百分比,指示表型变异在居群间贡献率的大小[16-17]。由表6可知,11个余甘子果实表型性状在居群间的平均方差分量百分比为20.95%,居群内为54.79%,说明余甘子果实表型性状在居群内和居群间均存在一定程度的变异,但居群内的多样性大于居群间的多样性,余甘子果实表型性状变异主要来源于居群内变异。其表型分化系数为16.67%~35.86%,平均为27.50%,单个果肉重最大,其次是出肉率和单果重,均大于30%,果形指数最少,核形指数次之,分别为16.67%和20.00%。

表6 余甘子果实表型性状的方差分量及群体间表型分化系数
2.2余甘子果实表型性状相关性
从表7可知,11个表型性状间存在不同程度的相关性。单果重、单个果肉重、果核重、果核横径、果核纵径、果肉厚度、出肉率均与果纵径和果横径呈极显著正相关,果横径、果纵径、单果重、单个果肉重、果核重、果核横径、果核纵径、果肉厚度间呈极显著正相关,果核横径、核形指数、出肉率与果形指数呈显著正相关,果形指数与果纵径呈极显著负相关,出肉率与果核重呈极显著负相关。果实表型性状间的显著相关性显示各性状在生长过程中的协调性。
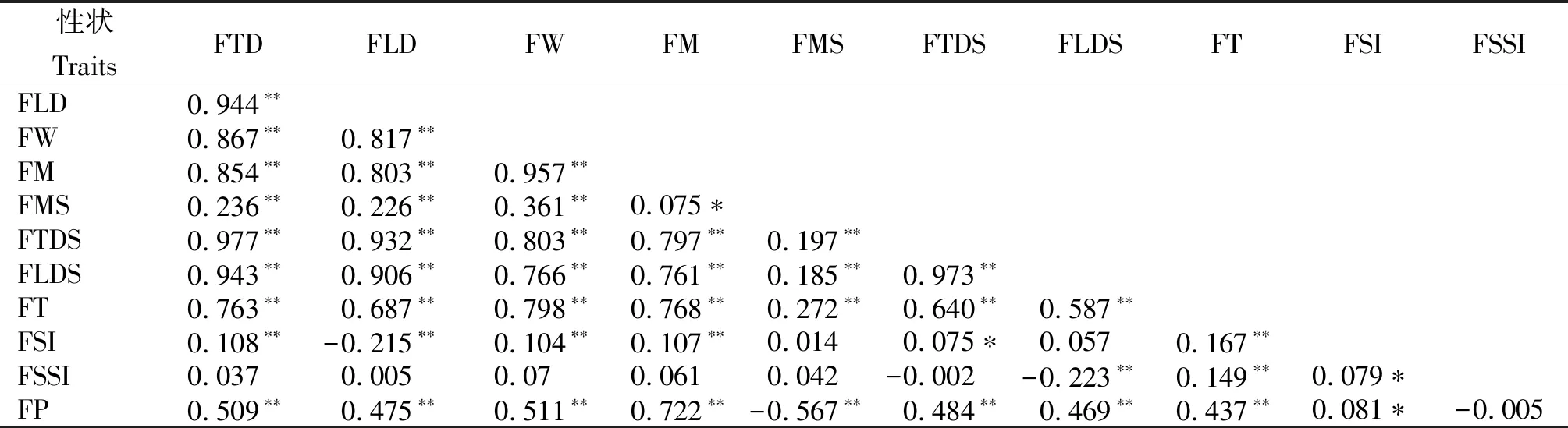
表7 余甘子果实表型性状间相关性
注:*和**分别表示相关性显著(P<0.05)和极显著(P<0.01)。
Note: * and ** indicate the correlation are atP<0.05 andP<0.01 level respectively.
2.3余甘子果实表型性状主成分分析
从表8可知,前3个主成分的累计贡献率达83.37%,表明前3个主成分代表了余甘子果实表型性状的大部分信息。第Ⅰ主成分的特征值为6.36,解释了11个表型性状57.84%的变化,特征向量值较大的为果横径、果纵径、单果重、单个果肉重、果核横径、果核纵径,说明第Ⅰ主成分主要是与果实大小相关的性状;第Ⅱ主成分的特征值为1.58,贡献率为14.32%,特征向量最大的为果核重,说明第Ⅱ主成分的主要决定因子为果核重;第Ⅲ主成分特征向量值为1.23,贡献率为11.21%,其中果形指数和核形指数的特征向量值较大,说明第Ⅲ主成分主要解释果实形状和果核形状特征。
根据特征向量矩阵及标准化的11个表型性状值,代入上述3个主成分中,由此可以获得3个因子的得分公式,如下:
表8 11个表型性状主成分分析
Table 8 Principal component analysis of phenotype traits inP.emblica
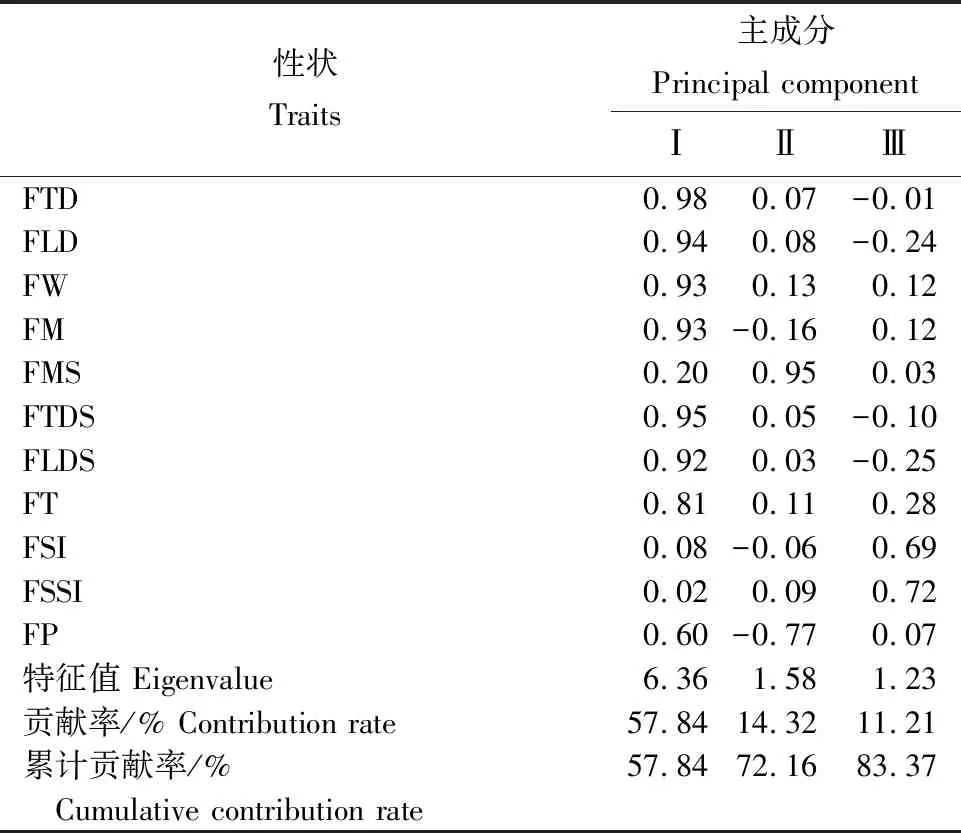
性状Traits主成分 Principal componentⅠⅡⅢFTD0.980.07-0.01FLD0.940.08-0.24FW0.930.130.12FM0.93-0.160.12FMS0.200.950.03FTDS0.950.05-0.10FLDS0.920.03-0.25FT0.810.110.28FSI0.08-0.060.69FSSI0.020.090.72FP0.60-0.770.07特征值 Eigenvalue6.361.581.23贡献率/% Contribution rate57.8414.3211.21累计贡献率/% 57.8472.1683.37 Cumulative contribution rate
F1=0.39X1+0.37X2+0.37X3+0.37X4+0.08X5+0.38X6+0.36X7+0.32X8+0.03X9+0.01X10+0.24X11
F2=0.06X1+0.06X2+0.0.10X3-0.13X4+0.76X5+0.04X6+0.02X7+0.09X8-0.05X9+0.07X10-0.61X11
F3=-0.01X1-0.22X2+0.11X3+0.11X4+0.03X5-0.09X6-0.23X7+0.25X8+0.62X9+0.65X10+0.06X11
根据F1、F2、F3数值,以及各主成分的贡献率权重(0.69、0.17、0.13)得出每个品种的综合得分公式:F=0.69F1+0.17F2+0.13F3,由上述公式计算F值,对68个余甘子单株表型性状进行综合评价,综合评价根据F值大小进行排序(表9),F值越高表型综合性越好。
从表9看出,排在前5位的分别是58号(P6居群)、52号(P5居群)、49号(P5居群)、55号(P5居群)、10号单株(P1居群)。其中P6居群的58号单株综合得分值(F值)最高,为1.82,说明58号单株的综合性状最好; P1居群的2号单株F值最低,为-2.24,表明2号单株的综合表现最差。
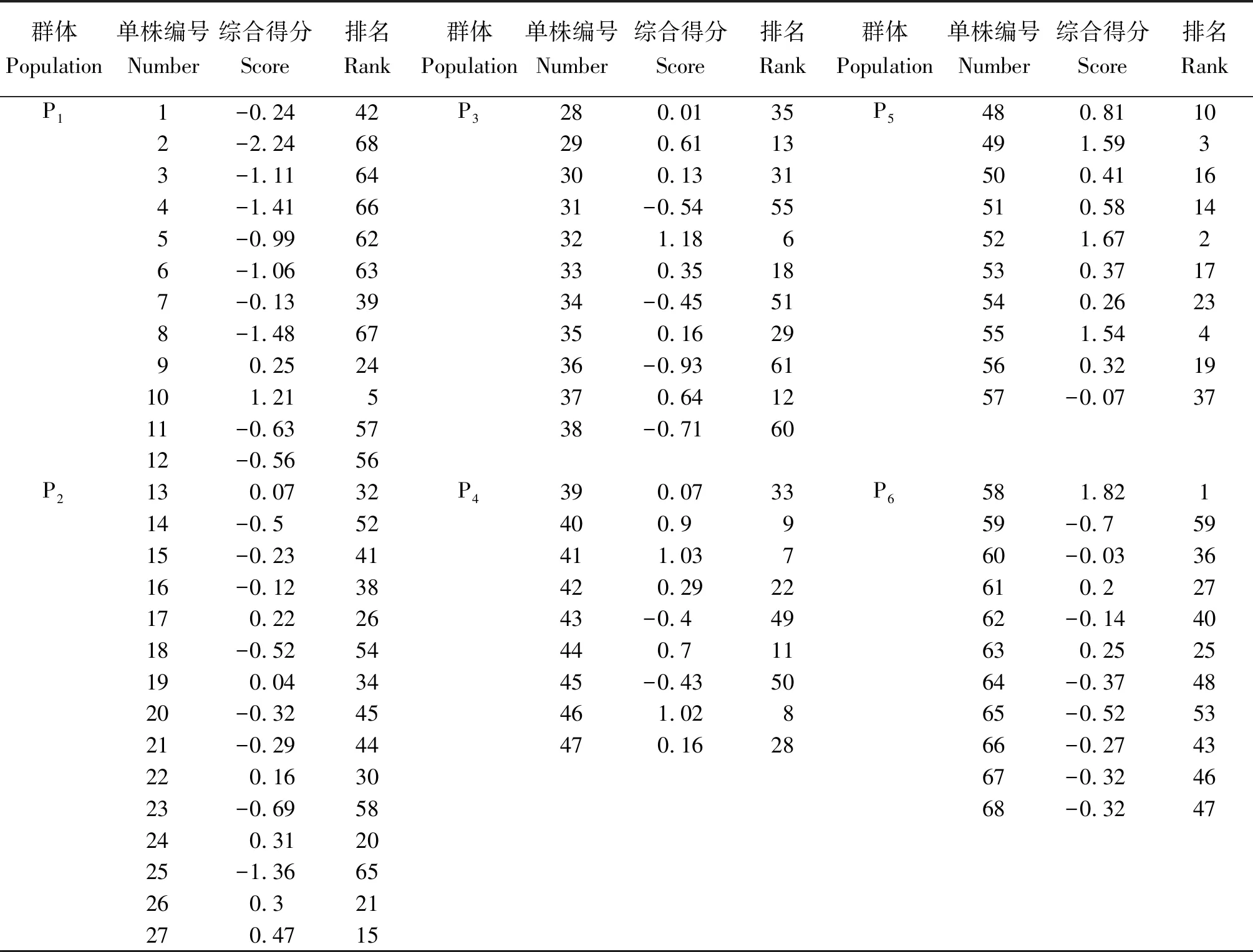
表9 余甘子果实表型性状的综合评价
2.4余甘子果实表型性状聚类情况
从图1可知,以欧式距离12.5为阈值,68个单株被分为5大类,Ⅰ类包括各个居群的45个单株;Ⅱ类包括P4居群的40号、41号、44号单株;Ⅲ类包括P1居群的3~6号单株;Ⅳ类包括P2居群的25号单株、P3居群的36号单株及P1居群的8号和2号单株, P1居群的2号单株单独聚为一小类;Ⅴ类包括各个居群的12个单株。其中Ⅲ类和Ⅳ类的8个单株综合排名均为倒数。总的来说6个居群的68个余甘子单株并未按地理距离而聚类,其果实表型性状无空间上的特异性。
3结论与讨论
表型性状变异是物种遗传变异的表现形式,也是基因型和环境条件共同作用的结果[18-19]。丰富的种质资源是选择育种的重要基础,表型性状在植物种属间亲缘关系鉴定和居群生物学分析中具有重要作用。表型多样性研究是植物遗传多样性研究中最基础、最直观的方法,能简便、快捷地对遗传资源作出评价[20-21]。选用17个果实表型性状对6个居群68个余甘子单株遗传多样性分析表明,余甘子果实形态特征变异丰富。6个果实描述性状表现出不同程度的遗传分化,Shannon-Weaver多样性指数在0.683 1~1.660 6,其中果实形状、果实颜色和果顶的多样性指数大于1,其余3个性状(果柄形状、腹缝线形状、果核粘离度)的多样性指数大于0.5。对11个表型性状的均值、标准差和多重比较分析表明,余甘子天然居群果实形态特征在居群间存在一定差异,其中P5居群果大、可食率高,P1居群果小、可食率较高,P4居群果大、可食率低。
变异系数可间接反映表现性状多样性的丰富程度,变异系数越大,说明性状的变异幅度越高,性状多样性越丰富[22]。研究表明,余甘子果实表型性状在居群间和居群内均存在一定差异且表现不一致。6个居群的平均变异系数为24.30%,变幅为21.73%~25.47%,P1和P6居群的平均变异系数较高,2个居群的果实表型性状多样较丰富;P5居群的平均变异系数最小,其果实表型性状变异程度较低。单果重、果横径等11个余甘子果实表型性状的变幅为6.16%~43.35%,平均为24.30%,其中果核重、单个果肉重和单果重的平均变异系数为38.50%,而果形指数和核形指数的变异系数分别为7.89%和6.16%,表明余甘子果实、种子的质量性状变异较大,而形状相对稳定,这与对无患子(Sapindusmukorossi)[18]、毛梾(Cornuswalteri)[19]种实表型多样性的研究结果一致。上述分析表明,不同性状对环境的适应和不同居群的环境差异导致了种群变异程度的差异。
野生余甘子在云南独特的地理气候条件使其进化积累了丰富的遗传变异。对供试材料的方差分析表明,6个居群余甘子果实表型性状在居群间的平均方差分量百分比为20.95%,居群内为54.79%,居群间和居群内存在广泛变异,但居群内变异大于居群间变异,说明余甘子果实表型性状变异主要来源于居群内变异,与瞿文林等[23]得出的结论相符合。余甘子果实表型性状的平均表型分化系数为27.50%,其中,单个果肉重、出肉率、单果重的表型分化系数较大,分别为35.86%、35.20%和30.28%,说明这3个性状相对于其他性状在居群间有较高的表型分化程度;而果形指数和核形指数的表型分化系数较小,分别为16.67%和20.00%,在居群间表型分化程度较低,较为稳定。余甘子果实表型分化在居群间与居群内的差异,一方面由于生态环境的差异,另一方面由于不同表型性状受不同基因型控制。居群内的变异是余甘子果实表型性状遗传变异的主要来源,在开发利用余甘子种质资源时应加大优良个体的选择和利用力度。
种质资源的遗传多样性直接影响着品种改良、新品种选育,目标改良性状的多样性越丰富,变异几率越大,越容易获得所期望的选择效果。余甘子果实表型性状在居群间和居群内丰富的变异为选择育种提供了丰富的材料。基于余甘子果实表型性状,采用隶属函数和主成分分析结合的方法,对68个余甘子单株进行综合评价,该方法已在植物表型综合评价方面得到应用[2,7,24]。主成分分析和综合评价结果显示,前3个主成分因子的累计贡献率达83.37%,表型性状综合得分F值排在前5位的分别是58号(P6居群)、52号(P5居群)、49号(P5居群)、55号(P5居群)和10号单株(P1居群)。P6居群的58号单株综合性状最好; P1居群2号单株的综合表现最差。通过比较综合得分F值的大小筛选出的优异种质资源,可以作为杂交育种亲本材料或优异种质在育种和生产中利用[20,24]。
通过对6个余甘子天然居群的果实表型多样性分析表明,余甘子天然居群在长期进化过程中产生一定程度的表型变异,且由于居群内的表型分化与生态环境因素的共同影响,居群内的变异大于居群间变异。在收集保存余甘子种质资源时,应注意不同层次的变异,既要考虑种群间的变异,也应考虑种群内变异,增加种群内个体数量,并优先收集有特殊表型的单株。
猜你喜欢
中国防痨杂志(2022年6期)2022-11-25
现代农业科技(2022年18期)2022-09-27
东北农业大学学报(2022年7期)2022-09-23
耕作与栽培(2022年1期)2022-04-07
——辽异1号
中国果业信息(2021年6期)2021-12-02
儿童绘本(2019年19期)2019-11-25
扬子江(2019年1期)2019-03-08
学与玩(2018年11期)2018-12-14
山东农业科学(2018年1期)2018-02-03
江苏农业科学(2017年7期)2017-05-23
