
雌核发育一代翘嘴鲌抑制素βB基因SNP筛选及其与生长性状的关联性分析
2020-06-02 10:13余钧剑迟美丽贾永义刘士力竺俊全顾志敏
江西农业大学学报 2020年2期
余钧剑,迟美丽,贾永义,刘士力,竺俊全,顾志敏*
(1.浙江省淡水水产研究所,浙江 湖州313001;2.宁波大学 海洋学院,浙江 宁波315211)
【研究意义】翘嘴鲌(Culter albur nus)俗名白鱼,隶属于鲤科、鲌亚科、鲌属,其适应性较强,广泛分布于我国长江中下游地区各水域。翘嘴鲌肉白且细嫩,味美而不腥,一贯被视为上等经济鱼类,具有很高的营养价值,是有名的“太湖三白”之一[1-2]。通过对遗传背景一致的全同胞翘嘴鲌苗种同塘养殖实验发现,性成熟时雌鱼体质量高于雄鱼15%以上,因此养殖全雌翘嘴鲌具有较好的市场前景[3]。雌核发育技术可实现雌性单性化;并且雌核发育鱼类为母系遗传,能够保持母本优良性状,可以大幅度缩短选育周期,高效地实现选育目标,是快速育种繁殖的有效手段[4-7]。翘嘴鲌具有较高的经济价值和营养价值,适应于我国各大淡水养殖环境,而雌核发育技术可快速实现有利基因纯合固定。因此,为了快速而高效地改良翘嘴鲌经济状,获得具有良好养殖性能的翘嘴鲌新品种,开展对雌核发育翘嘴鲌快速生长个体的选育工作,以及从相关基因层面探索翘嘴鲌快速生长机制具有重要意义。【前人研究进展】抑制素(inhibin,INH)是一种二聚体糖蛋白激素,主要由雄性睾丸支持细胞和雌性卵巢颗粒细胞分泌[8]。抑制素由α亚基和不同的β亚基通过二硫键连接而成,其中β亚基有两种类型(βA和βB)。α亚基和βB亚基构成的抑制素称为抑制素B[9]。抑制素属于转化生长因子β(transforming growth factorβ,TGFβ)超家族成员,该超家族的多个基因均已证明直接或间接与肌肉生长发育相关[10]。而且抑制素在多个组织内均有分布,通过激素调控、自分泌和旁分泌的方式来参与细胞功能和细胞增殖[11]。在人类中都发现抑制素水平与子痫前期胎儿的生长受限有关[12]。Vassalli等[13]通过小鼠的活化素/抑制素βB基因突变的研究表明抑制素βB基因在小鼠胚胎发育后期起作用,这些研究表明抑制素基因可能参与到机体的生长发育过程。【本研究切入点】继限制性片段长度多态性(RFLP)和简单重复序列(SSR)后的第3代分子标记——单核苷酸多态性(single nucleotide polymorphism,SNP)是指在基因组上单个核苷酸的变异[14],SNP标记具有突变位点数目多、高度的遗传稳定性、有利于实现高通量检测等优点[15]。四引物扩增受阻突变体系PCR法(tetra-primer amplification refractory mutation system PCR,tetra-primer ARMS PCR)是一种在普通PCR基础上发展起来并专门用于检测SNP的衍生技术。该技术在SNP两侧分别设计两条内引物,内引物的3′末端分别与两种等位基因相匹配,并在SNP两侧距离不等的适当距离设计两个外引物;根据扩增产物特异条带的长度大小和有无,实现单管PCR对SNP的分型[16]。Tetra-primer ARMS PCR技术用于检测单核苷酸突变具有快速、简便、低成本和可以区分等位基因是否纯合等优点[17]。目前,通过Tetra-primer ARMS PCR,已经成功在中华鳖[18]、中国对虾[19]和牛[20]中筛选到与生长相关的SNP位点,这些位点已被证明具有用作该生物生长性状分子标记辅助育种的潜力。作者所在团队多年来进行雌核发育翘嘴鲌选育研究,在此研究过程中发现雌核发育一代个体大小差异明显,是作为筛选生长相关分子标记的良好素材。因此,以上研究为以采用四引物扩增受阻突变体系PCR对雌核发育一代翘嘴鲌抑制素βB基因SNP位点分型提供了条件。【拟解决的关键问题】本研究通过翘嘴鲌转录组数据库对抑制素βB基因外显子序列进行扩增,利用直接测序获得候选SNP位点。然后采用四引物扩增受阻突变体系PCR的检测方法,对雌核发育一代翘嘴鲌抑制素βB基因进行SNP位点分型,研究不同SNP分型与其体长、体质量、体厚、体高、全长、尾柄长、尾柄高之间的关联分析,探讨抑制素βB基因的不同基因型对生长的影响情况,通过这些研究探索雌核发育翘嘴鲌快速生长的机制,获得特定的辅助选育标记,为翘嘴鲌的分子标记辅助育种提供重要参考。
1 材料与方法
1.1 材料
2017年6月通过对普通雌性翘嘴鲌的卵与灭活的鲤鱼精子进行授精及冷休克处理后获得的雌核发育一代翘嘴鲌幼苗,养殖于浙江省淡水水产研究所综合试验基地同一池塘内,保持生长环境及饲养条件一致。2018年3月对同一批次10月龄的雌核发育一代翘嘴鲌进行拉网,从该群体中随机取样144尾。所选个体分别用电子天平称取样品的体质量,用游标卡尺逐尾测量体长、体厚、体高、全长、尾柄长、尾柄高等多个形态性状。分别剪取每尾翘嘴鲌的尾鳍置于无水乙醇中,-20℃冻存备用,用于提取DNA。
1.2 基因组DNA的提取
使用TIANamp Genomic DNA Kit试剂盒(TIANGEN),提取雌核发育一代翘嘴鲌尾鳍中的DNA。使用10 g/L琼脂糖凝胶电泳,凝胶成像系统检测结果,确定提取DNA的质量。用Eppendorf Bio Spectrometer紫外分光光度计测定光密度值,确定样本DNA浓度和纯度,于-20℃保存,备用。
1.3 翘嘴鲌抑制素βB基因SNPs位点的筛选
从本课题组已有的翘嘴鲌转录组数据库中获得的抑制素βB基因mRNA序列,然后与本课题组已有的翘嘴鲌全基因组数据进行分析,可知翘嘴鲌抑制素βB基因含有2个外显子,采用Primer Premier 6.0软件设计2对引物:INHBBf1(5′-GGTGCTGTCCACGCTGGTGG-3′)和INHBBr1(5′-CGAAGCTGACTATCTCGTAT-3′);INHBBf2(5′-CACCAGTCTGTCATTCCAGT-3′)和INHBBr2(5′-ACCCGCAGGACTCGACGATC-3′)。通过这两对引物来筛选外显子1和外显子2上的SNP。从144尾雌核发育一代翘嘴鲌DNA样本中选取25个DNA样本扩增上述两对引物。25个DNA样本选取标准如下:体质量最大的10尾个体的DNA样本,依次分成两组;体质量接近平均值的5尾个体的DNA样本,分成一组;体质量最小的10尾个体的DNA样本,依次分成两组。将每组5尾个体的DNA样本相混合,作为模板,进行PCR扩增。PCR反应体系共25µL:上、下游引物各0.5µL,DNA 1µL,2×Taq PCR MasterMix 12.5µL,ddH2O 10.5µL。其中上、下游引物浓度均为10µmol/L,稀释后的DNA浓度约为50 ng/µL。PCR扩增程序:94℃预变性5 min;94℃变性30 s,58℃退火30 s,72℃延伸30 s,共进行30个循环;72℃延伸5 min,4℃保存。将扩增产物用1.5%琼脂糖凝胶电泳30 min,置于Bio-Rad凝胶成像系统Image-lab下观察,拍照。选取目的条带清晰明亮且无杂带的PCR产物送到上海生工公司测序。测序结果用BioEdit软件进行分析,共获得2个SNP位点。
1.4 四引物扩增受阻突变体系引物设计
利用tetra-primer ARMSPCR引物在线设计程序进行引物设计(http://primer1.soton.ac.uk/primer1.html),产物片段大小控制在150~700 bp。针对每个SNP位点设计两条3′末端分别与SNP两个等位基因碱基配对且延伸方向相反的内侧引物和2个外引物,同时在内引物3′端2或3位碱基引入错配以增加扩增特异性(表1,错配碱基用斜体字母和下划线标注)。引物由上海生工生物工程技术服务有限公司合成。

表1 翘嘴鲌抑制素βB基因SNP引物序列Tab.1 Sequen of SNP primers on INH B B gene in topmouth culter
1.5 SNP位点的分型与验证
本实验也对各个位点的tetra-primer ARMS PCR反应体系进行了优化。PCR反应体系共25µL,主要包括10.1µL ddH2O、1µL 20 ng/µL模板、12.5µL PCR Mix和2µL 10 Mm内外引物。引物浓度均为10µmol/L,稀释后的DNA模板浓度约为50 ng/µL,Mix为2×Taq PCR MasterMix。设定12个温度梯度(依次为54.0,54.3,55.1,56.2,57.2,58.8,60.2,61.5,62.8,63.9,64.7,65.0℃)为退火温度,探索每组引物的退火温度,根据SNP位点分型情况和条带的清晰程度,以确定最佳的退火温度。在最适退火温度下,在25µL反应体系中对内、外引物各浓度比例组合在4个比例水平(4∶1、2∶1、1∶1、1∶2)上进行优化。以上探索实验中PCR反应程序为:94℃预变性5 min;然后进行30个循环,每个循环包括94℃变性30 s,退火30 s(退火温度Tm,表1),72℃90 s;最后72℃5 min。PCR产物用2%琼脂糖凝胶电泳40 min,置于Bio-Rad凝胶成像系统Image-lab下观察,拍照。
1.6 数据处理
采用SNPStats软件统计基因型个体数、基因型频率和等位基因频率等遗传参数。采用SPSS 22软件的独立样本T检验和一般线性模型,对雌核发育一代翘嘴鲌各个样本生长性状(体长、体质量、体厚、体高、全长、尾柄长、尾柄高)与抑制素βB基因各位点基因型进行相关性分析。
2 结果与分析
2.1 雌核发育一代翘嘴鲌抑制素βB基因SNPs位点的筛选
对雌核发育一代翘嘴鲌抑制素βB基因外显子1和外显子2的序列进行分段克隆,直接测序后进行峰图比较筛查SNPs位点。在抑制素βB基因外显子2序列上获得2个SNP位点(图1和图2),分别命名为c.717G>T和c.778G>A。
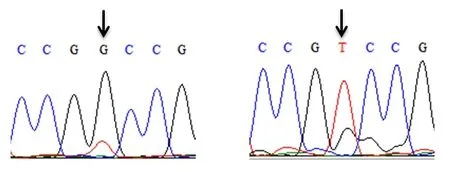
图1 翘嘴鲌抑制素βB基因c.717G>T位点测序峰图Fig.1 The sequencing peak chart of c.717G>T locus on INHBB gene in topmouth culter
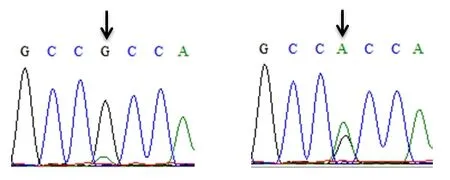
图2 翘嘴鲌抑制素βB基因c.778G>A位点测序峰图Fig.2 The sequencing peak chart of c.778G>A locus on INHBB gene in topmouth culter
2.2 雌核发育一代翘嘴鲌抑制素βB基因的SNP位点ARMS PCR分型结果
通过调整tetra-primer ARMS PCR反应条件,最后确定在c.717G>T位点的tetra-primer ARMS PCR反应体系中,退火温度为61.5℃,内、外引物各浓度比例为2:1(即内引物上游引物用量、内引物下游引物用量、外引物上游引物用量、外引物下游引物用量分别为0.6,0.6,0.3,0.3µL)时,c.717G>T的SNP位点获得良好分型效果。在c.778G>A位点的tetra-primer ARMS PCR反应体系中,退火温度为58.8℃,内、外引物各浓度比例为1∶1(即内引物上游引物用量、内引物下游引物用量、外引物上游引物用量、外引物下游引物用量分别为0.5,0.5,0.5,0.5µL)时,c.778G>A的SNP位点获得良好分型效果。对144个雌核发育一代翘嘴鲌样本基因组DNA进行扩增和分型,结果如下。
从图2和3中可以看出,每个样品基因组DNA可扩增两条条带,一条外引物产物和一条内引物产物,无代表杂合型的第3条条带出现。说明雌核发育一代翘嘴鲌抑制素βB基因c.717G>T和c.778G>A位点的基因型均为纯合型。由图3可知,c.717G>T位点处所有个体均能扩增出511 bp段的外引物产物,GG基因型仅能扩增出片段长度为457 bp的条带,而TT基因型仅能扩增出片段长度为123 bp的条带。由图4可知,c.778G>A位点处,所有个体均能扩增出511 bp的外引物产物,GG基因型仅能扩增出片段长度为367 bp的条带,而AA基因型仅能扩增出片段长度为188 bp的条带。结果表明ARMS PCR技术可以用于雌核发育一代抑制素βB基因c.717G>T和c.778G>A位点SNP分型,且结果与直接测序所得结果一致。
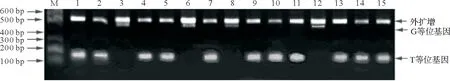
图3 翘嘴鲌抑制素βB基因c.717G>T位点的凝胶电泳结果Fig.3 Gel electrophoresis results of c.778G>A locus on INHBB gene in topmouth culter
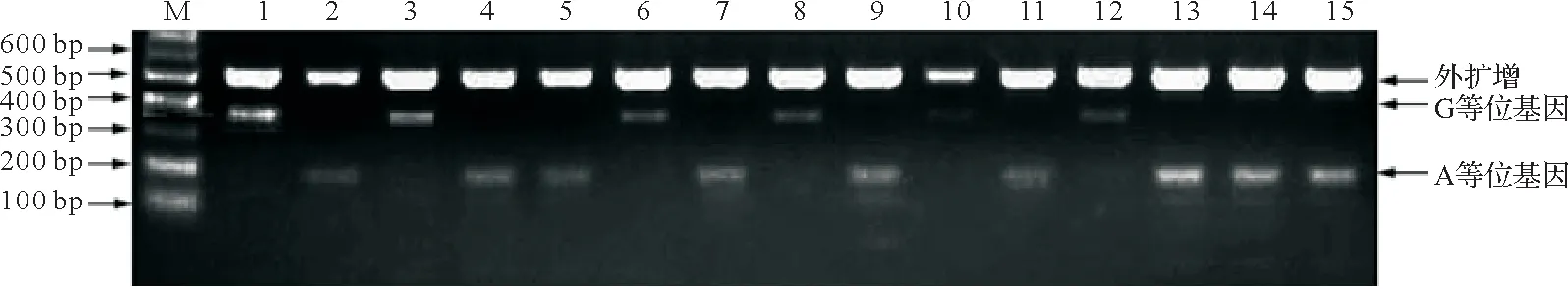
图4 翘嘴鲌抑制素βB基因c.778G>A位点的凝胶电泳结果Fig.4 Gel electrophoresis results of c.778G>A locus on INHBB gene in topmouth culter
2.3 抑制素βB基因c.717G>T和c.778G>A位点在雌核发育一代翘嘴鲌群体中的分布
由表2可知,通过统计144个雌核发育一代翘嘴鲌的基因型,发现抑制素βB基因c.717G>T和c.778G>A 2个SNP位点均只有两种基因型且无杂合型突变,均为纯合型。抑制素βB基因c.717G>T位点两种基因型样本个体数目相近,该位点基因型和等位基因频率也相近。另一方面,抑制素βB基因c.778G>A位点的GG基因型个体数高于AA基因型,GG基因型个体数是AA基因型个体数的2倍,基因型GG频率和等位基因G频率也显著高于基因型AA频率和等位基因A频率。因此,推测抑制素βB基因c.778G>A位点的等位基因G为优势等位基因。
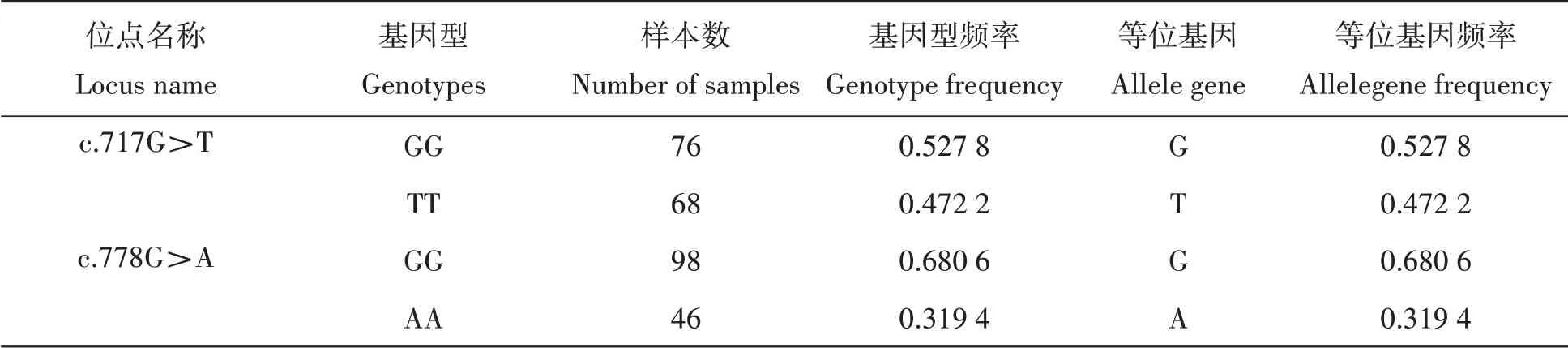
表2 c.717G>T和c.778G>A位点的基因型及其等位基因频率Tab.2 Genotypes and allele gene frequency inc.717G>T and c.778G>A locus
2.4 雌核发育一代翘嘴鲌抑制素βB基因c.717G>T和c.778G>A位点与生长性状的关联性分析
由表3可知,采用独立样本T检验,将抑制素βB基因c.717G>T和c.778G>A位点单倍型与雌核发育一代翘嘴鲌中个体的体长、体质量、体厚、体高、全长、尾柄长、尾柄高等7个性状进行相关性分析后发现:抑制素βB基因c.717G>T位点两种基因型和c.778G>A位点两种基因型与7个生长性状之间都存在显著差异(P<0.05)。其中抑制素βB基因c.717G>T位点GG基因型的个体的体长、体质量、体厚、体高、全长、尾柄长、尾柄高均显著高于TT基因型的个体(P<0.05),GG基因型的个体的平均体质量是TT基因型的个体的1.7倍。抑制素βB基因c.778G>A位点GG基因型的个体的7个性状也均显著高于AA基因型的个体(P<0.05),GG基因型的个体的平均体质量是AA基因型的个体的2.0倍。
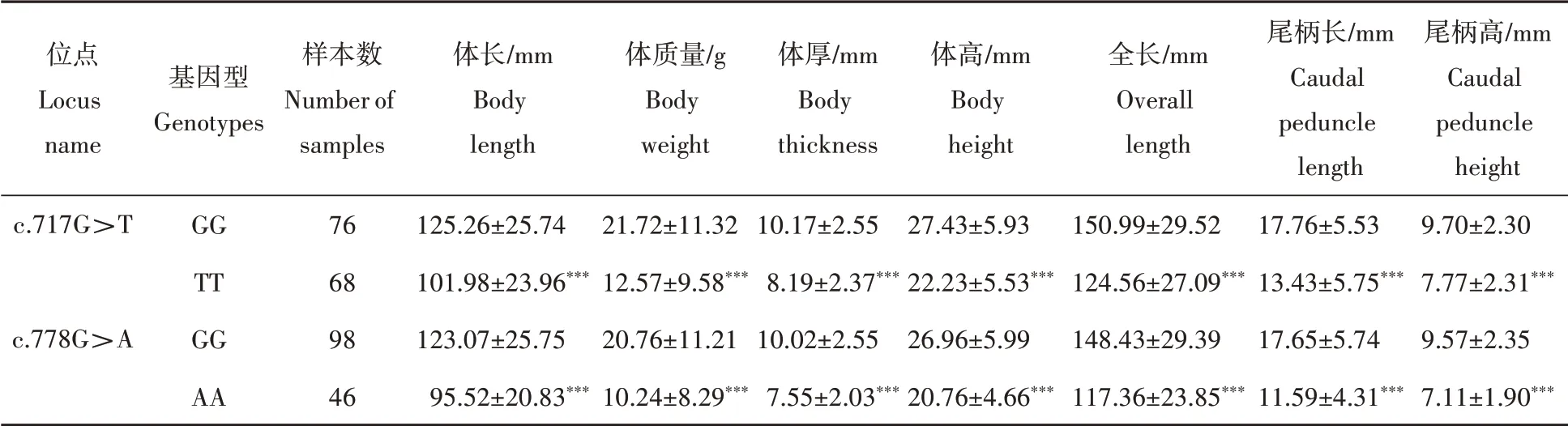
表3 翘嘴鲌抑制素βB基因SNP位点不同基因型的生长性状指标Tab.3 Growth traits of different genotype of SNP locus on IN HBB gene in topmouth culter
c.717G>T和c.778G>A位点2种基因型两两组合,可得到4种双倍型,分别为D1(基因型GG&GG,频率为50.7%)、D2(基因型TT&GG,频率为18.1%)、D3(基因型TT&AA,频率为29.2%)、D4(基因型GG&aAA,频率为2.1%)。除去频率低于3%的双倍型组合D4,采用一般线性模型,将3种双倍型与144尾雌核发育一代翘嘴鲌中个体的体长、体质量、体厚、体高、全长、尾柄长、尾柄高等7个性状进行相关性分析,结果见表4。分析发现双倍型D1(GG&GG)的个体体长、体质量、体厚、体高、全长、尾柄长、尾柄高等7个性状与双倍型D2(TT&GG)个体之间的差异不显著(P>0.05);而双倍型D3(TT&AA)个体的体长、体质量、体厚、体高、全长、尾柄长、尾柄高等7个性状显著低于双倍型D1(GG&GG)和双倍型D2(TT&GG)个体(P<0.05)。
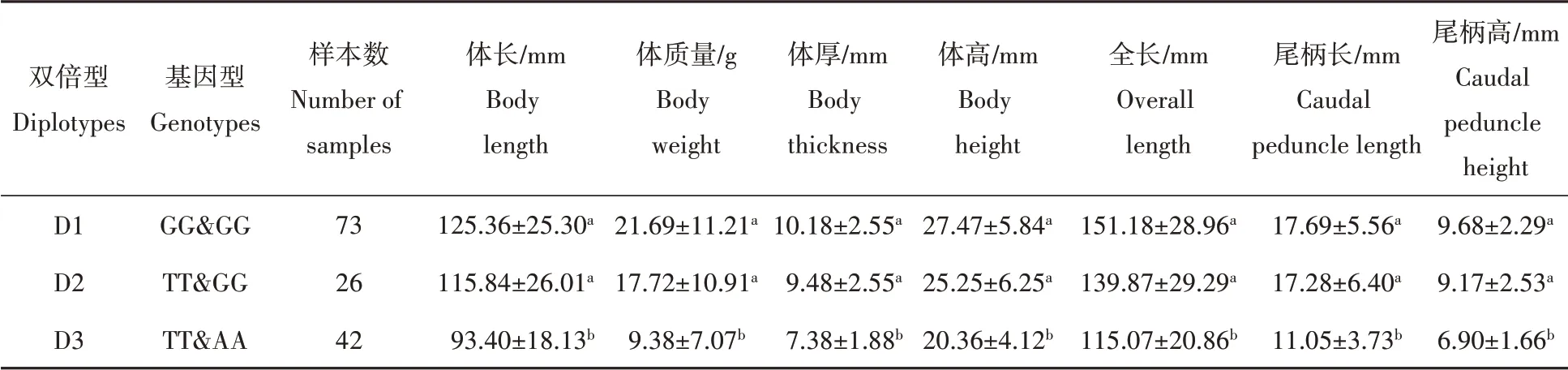
表4 抑制素βB基因不同双倍型的生长性状指标Tab.4 Growth traits of different diplotype of SNP locus on I N H B B gene in topmouth culter
3 讨 论
SNP是基因组水平上的单核苷酸的变异,具有密度高、遗传稳定性高和代表性强、易实现自动化分析的优点,在遗传多样性结构分析上具有广阔的应用前景,未来的SNP的分型检测方法将向更高效、更准确、更灵活的方向发展[21-23]。SNP是负责表型变异或者是选择的直接靶点,而且在斑节对虾、罗氏沼虾、太平洋牡蛎等水产动物中已被证明某些SNP和经济性状关联[24-26]。本研究通过直接测序方法对雌核发育一代翘嘴鲌抑制素βB基因进行分析,发现了2个SNP位点,并用四引物扩增受阻突变体系PCR成功对c.717G>T和c.778G>A这2个位点进行了分型和验证。分型结果显示2个位点均存在2种基因型,且均为纯合子。翘嘴鲌的雌核发育技术采用紫外线照射灭活的普通鲤鱼(Cy pr i nus car pio)精子来刺激普通翘嘴鲌的卵子,利用冷休克法抑制卵子第二极体的释放,从而促使染色体二倍化[27-28]。由于雌核发育翘嘴鲌在受精过程卵母细胞不分裂出第二极体,卵母细胞中同一条染色体的两条姐妹染色单体分离组成雌核发育翘嘴鲌基因组中的同源染色体,所以雌核发育翘嘴鲌基因型理论上为纯合子,这与本研究结论一致。
生长性状是评价水产动物遗传育种有价值性状的重要标准之一,主要通过增加生长速率,达到提高经济效益的目的[29]。本研究以雌核发育一代的翘嘴鲌为研究对象,分别将抑制素βB基因c.717G>T位点2种基因型、c.778G>A位点2种基因型以及c.717G>T位点和c.778G>A位点组成的3种双倍型与雌核发育一代翘嘴鲌中个体的体长、体质量、体厚、体高、全长、尾柄长、尾柄高等7个性状进行相关性分析。抑制素βB基因c.717G>T位点GG基因型的个体与7个生长性状之间的显著性均显著高于TT基因型的个体(P<0.05)。但是c.717G>T位点的G→T突变使编码氨基酸的密码子CGG变成CGU,该突变为同义突变,未导致编码氨基酸的改变;并且将测序获得的抑制素βB基因序列与斑马鱼该基因序列进行序列比对分析,结果显示该位点位于抑制素βB基因开放阅读框内。c.717G>T位点的突变虽未导致氨基酸的改变,但近年来有研究表明,不同基因密码子存在使用偏好性,这可能导致mRNA二级结构的稳定性或者影响核糖体通过mRNA的速度受到影响,进而直接或者间接影响基因的转录效率、mRNA翻译以及蛋白质的结构与活性,来影响蛋白翻译[30-31]。调控核内靶基因转录受到影响后,可能使得在雌核发育一代翘嘴鲌7个生长性状上,纯合GG型个体显著高于TT型个体。抑制素βB基因c.778G>A位点的G→A突变使编码丙氨酸的GCC变成编码苏氨酸的ACC,为非同义突变;而且该位点也位于基因cDNA的开放阅读框内。c.778G>A位点GG基因型的个体的7个性状均显著高于AA基因型的个体(P<0.05),表明c.778G>A位点的多态性对雌核发育一代翘嘴鲌生长性状也产生影响作用。笔者推测这是由于基因的单核苷酸突变,使编码的丙氨酸突变为苏氨酸,这种变化可能导致其编码蛋白的结构产生变化,如大口黑鲈组织蛋白B基因C/G转换,以及皮尔蒙特牛和比利时蓝牛肌肉生长抑素基因的G/A转换,由于某个基因突变而导致该生物生长性状得到改良[32-33],可作为高效的分子标记应用于分子育种。
优异基因的聚合能得到理想的累加效应,基因聚合最早由Yadav等[34]研究改良芥菜(Bras sica juncea)抗逆和抗病性状时提出。目前,基因聚合效应已经在水产育种方面有报道。徐磊[35]研究了大黑口鲈(Micro pterus salmoi de)8个分子标记的基因聚合效应,发现随着优异基因型数量的增加,平均体质量同步递增。基因型的聚合效应并不是各基因型效应所造成的表型的简单相加,而且纯合基因型互相组合具有显著的累加效应,能够稳定的遗传给子代。本研究中c.717G>T位点GG基因型和c.778G>A位点GG基因型并不存在交互作用,双倍型D1(GG&GG)个体的体长、体质量、体厚、体高、全长、尾柄长、尾柄高具有最大均值,但较其它基因型组合差异并不显著,因而这两个优异基因型不存在明显聚合效应。然而,双倍型与生长性状的关联性型分析显示,雌核发育一代翘嘴鲌抑制素βB基因c.778G>A位点基因型为GG时,双倍型D1(GG&GG)和双倍型D2(TT&GG)的个体的体长、体质量、体厚、体高、全长、尾柄长、尾柄高之间的差异均无统计学意义(P>0.05),c.717G>T位点碱基突变对生长性状的影响较小;而c.717G>T位点基因型为TT时,双倍型D2(TT&GG)个体的7个性状显著高于双倍型D3(TT&AA)个体,c.778G>A位点基因型GG个体的生长性状显著高于基因型AA个体。因此推断抑制素βB基因c.717G>T位点和c.778G>A位点的突变对生长性状的影响相独立和互不干扰,但是c.778G>A位点为对生长性状的影响更加明显。
综上所述,根据直接测序方法获得2个翘嘴鲌抑制素βB基因SNP位点,设计2组引物,分别进行四引物扩增受阻突变体系PCR,成功鉴别出雌核发育一代翘嘴鲌抑制素βB基因c.717G>T位点和c.778G>A位点的基因型,所得结果与直接测序所获得的突变类型一致。将这两个位点基因进行分型并与雌核发育一代翘嘴鲌中个体的体长、体质量、体厚、体高、全长、尾柄长、尾柄高等7个性状进行相关性分析,结果表明抑制素βB基因c.717G>T位点GG基因型和c.778G>A位点GG基因型的雌核发育一代翘嘴鲌具有生长优势,而且c.778G>A位点GG基因型对雌核发育一代翘嘴鲌生长性状的影响更加明显。因此,本研究表明采用tetra-primer ARMS PCR技术用于翘嘴鲌抑制素βB基因c.717G>T和c.778G>A位点SNP位点分型是可行的,抑制素βB基因c.717G>T位点GG基因型和c.778G>A位点GG基因型均可作为雌核发育一代翘嘴鲌分子标记辅助选育的候选基因。通过tetra-primer ARMS PCR技术可快速进行翘嘴鲌抑制素βB基因多态性的鉴别,在实际生产中可节省时间和育种成本,加快翘嘴鲌分子标记选育工作进程。
致谢:郑建波博士在实验过程中给予了指导,谨致谢意!
猜你喜欢
上海金属(2021年6期)2021-12-02
昆明医科大学学报(2021年3期)2021-07-22
烟草科技(2021年6期)2021-06-24
电脑知识与技术(2018年19期)2018-11-01
中华骨与关节外科杂志(2017年1期)2017-05-17
上海农业学报(2017年3期)2017-04-10
制造技术与机床(2015年10期)2015-04-09
中国中医药现代远程教育(2014年21期)2014-03-01
中国中医药现代远程教育(2014年17期)2014-03-01
作物研究(2014年6期)2014-03-01
