
玉米Zmglp基因的克隆及生物信息学分析
2020-05-23 02:07苗馨心单立杰吴委林郑大浩朴世领
延边大学农学学报 2020年1期
苗馨心, 单立杰, 吴委林, 郑大浩, 朴世领
(延边大学农学院,吉林 延吉 133002)
玉米不仅是世界上产量最高的谷类粮食作物,也是重要的经济作物和饲料作物。在我国,玉米的播种面积和总产量已经超过水稻和小麦,成为我国第1大粮食作物[1]。然而玉米病害一直是玉米生产中的重要限制因素,其中,弯孢菌叶斑病发生严重时,病斑密布整片叶片,形成大面积坏死,致使叶片枯死[2],严重影响了我国玉米生产的安全。
植物在生长发育过程中进化出了应对生物与非生物胁迫的防卫反应。作为病程相关蛋白(pathogenesis-related proteins, PRs)家族的一员, 类萌发素蛋白(germin-like proteins,GLPs)是一类普遍存在的功能性糖蛋白[3], 具有产生过氧化氢(H2O2)的SOD和草酸氧化酶(OXO)的酶活性[4-5],与植物抵御外源物质伤害及多种逆境胁迫密切相关[6],在植物的生长发育和防卫反应中有重要作用[3]。据报道,在受到真菌感染后,植物细胞内GLPs的表达上调,并且通过H2O2信号途径来调节植物对真菌的防御,进一步分析,GLPs通常与细胞壁非共价结合[7-8],通过NO和H2O2的积累,来调控POD、CAT、SOD、PAL、Glu等诸多防御酶的活性,进而增强抵御弯孢菌的扩展[9]。
虽然前期研究已经证实Zmglp基因在玉米抗弯孢菌叶斑病中起到重要作用,但尚未在不同抗性材料之间进行该基因的克隆及生物信息学分析。该研究克隆了抗病自交系Mo17与感病自交系黄早4的Zmglp基因,通过生物信息学方法分析该基因的理化性质、蛋白质构象及生理功能,为进一步深入研究Zmglp在抵御弯孢菌侵染的作用机理奠定基础。
1 材料与方法
1.1 材料
选取抗病品种Mo17和感病品种黄早4,种子经催芽处理12 h后,播种于农学院教学实验基地大棚中,待至长出3片叶时,取第3片叶片为试验材料。
1.2 方法
1.2.1 玉米叶片DNA的提取
剪取100 mg玉米新鲜叶片,加入液氮后充分研磨,使用改良尿素法[10]提取玉米DNA,通过凝胶电泳检测DNA纯度,保存于-20 ℃冰箱,待用。
1.2.2Zmglp基因克隆与测序
首先,以玉米标准基因组数据库(http://www.maizegdb.org)上下载的玉米Zmglp DNA序列Zm00009a028485_T001为基础,引用文献中[11]所设计的Zmglp正向引物ZmglpF1: 5’-GTTGCCATGGCCAAAATGGTG-3’,反向引物为ZmglpR1: 5’-CAGATTAACAGCATGCGGCAC-3’。在玉米标准基因组中引物区间内目标DNA序列长度为1 026 bp。
以玉米自交系Mo17和黄早4基因组DNA为模板进行RT-PCR扩增。采用梯度PCR的方法摸索扩增所需最佳温度,PCR 反应体系(25 μL)为:DNA模板2.5 μL,2×Go Taq Green Mix 12.5 μL,ddH2O 7.5 μL,10 μM上下游引物各1.25 μL。PCR 反应程序为:95 ℃预变性5 min;95 ℃变性1 min,56.9 ℃,退火30 s,72 ℃延伸180 s,35个循环;72 ℃延伸5 min;4 ℃保存。PCR扩增产物经1%琼脂糖凝胶电泳检测,回收后连接pMD19-T,再转化到DH5α中,挑取阳性单菌落进行液体培养,后稀释划线。准确无误后从大肠杆菌中提取和纯化质粒,经琼脂糖凝胶电泳检测正确后,再进行PCR验证,验证目的条带正确后,将相应的菌液进行扩大培养,保存菌种后,送到上海生工公司测序。
1.2.3 生物信息学分析
测序后得到的DNA序列,通过DNAStar软件和DNAMAN软件进行同源性分析。
目标DNA序列编码的蛋白质序列分析,利用NCBI在线工具(http://www.ncbi.nlm.nih.gov/),对蛋白质序列进行同源比对与保守域分析;利用ProtParam tool(https://web.expasy.org/protparam/) 在线工具和软件DNAMAN完成了对核酸及氨基酸序列组成分析、编码蛋白的理化性质分析;利用SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)在线工具预测蛋白质二级结构;利用ProtScale(https://web.expasy.org/protscale/)在线工具完成蛋白质亲疏水性分析;利用在线工具TMHMM 2.0 (http://www.cbs.dtu.dk/services/TMHMM-2.0/)对蛋白质跨膜区分析和Plant-mPLoc(http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/)进行亚细胞定位;利用NetPhos 3.1 Server(http://www.cbs.dTu.dk/services/NetPhos/)进行磷酸化位点预测;利用 YinOYang l.2(http://www.cbs.dtu.dk/services/YinOYang/)进行O-β-葡萄糖糖基化位点预测;利用DNAMAN 、MEGA6.06软件完成核酸和氨基酸序列的多序列比对和系统发育树的构建;利用SWISS-MODEL(https://swissmodel.expasy.org/interactive)预测蛋白质三级结构。
2 结果与分析
2.1 基因克隆
为摸索该引物从玉米自交系Mo17和黄早4中分离目标片段的PCR温度条件,在正向引物和反向引物的适宜范围内设置温度梯度,PCR仪自动设置的8个温度梯度为:52.0、52.5、53.5、55.1、56.9、58.4、59.4和60.0 ℃。经梯度PCR扩增,Mo17与黄早4均得到特异性条带(图1)。其中,Mo17与黄早4均在56.9 ℃时条带最清晰,该片段长度1 026 bp。
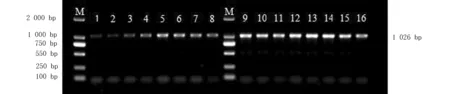
注:M:Marker 2000;Lane 1~8:ZmglpMo17 PCR;Lane 9~16:ZmglpHuangzao4 PCR引物扩增的目标DNA片段长度为1 026 bp。
图1 玉米自交系Mo17和黄早4的基因组DNA为模板的PCR条件
Fig.1 PCR reaction conditions using DNA from maize inbred Mo17 and Huangzao 4 as template
2.2 目标序列分析
2.2.1 目标DNA序列及其目的基因编码的蛋白质序列分析
从玉米自交系Mo17和黄早4中均成功分离到含有完整目的基因的DNA片段,在NCBI网站上通过Blast比较,确认从自交系Mo17所克隆得到的Zmglp基因序列与已公布的Zmglp序列(登录号为AY394010.1)的基因序列相似性达100%,证实所得到的序列是玉米Zmglp。从自交系Mo17中克隆的基因片段长度为1 026 bp,从自交系黄早4中克隆的基因片段长度为1 019 bp(图2),2个自交系都具有完整的开放阅读框(ORF),且开放阅读框所编码的蛋白质完全一致(图3),均由212个氨基酸组成。
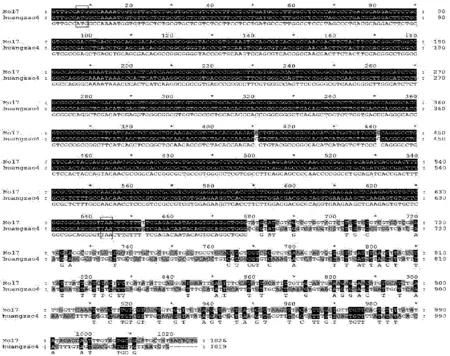
图2 玉米自交系Mo17和黄早4基因序列的比较
经进一步序列比较发现,二者在ORF区的410 bp处与440 bp处存在碱基同义突变,且3’端有大量碱基突变(图2)。根据此推测可能该基因在不同品种中调控机制不同,表达量可能会有差别。

图3 玉米自交系Mo17和黄早4编码蛋白质的比较
2.2.2 目标序列的生物信息学分析
2.2.2.1Zmglp编码蛋白的理化性质分析
在线预测玉米Zmglp的理论等电点蛋白质相对分子质量得出:Zmglp理论PI=6.01,Mw=21 887.34,分子式为C997H1562N248O288S8,Leu的含量最大,占12.3%,含有 19个芳香族氨基酸(Phe、Trp、Tyr),占8.9% ,Zmglp负电荷氨基酸残基(Asp+Glu)的数目共有12个,正电荷氨基酸残基(Arg+Lys)的数目共有10个,不稳定指数为23.72(<40),因此,该蛋白是稳定的。
对蛋白质进行疏水性/亲水性分析时,采用Kyte&Doolittle标度计算,结果显示Arg亲水性最强;Tle疏水性最强(图4)。总平均亲水性值(GRAVY)为0.526,因此,该蛋白为亲水性蛋白,能溶解在水溶液中。
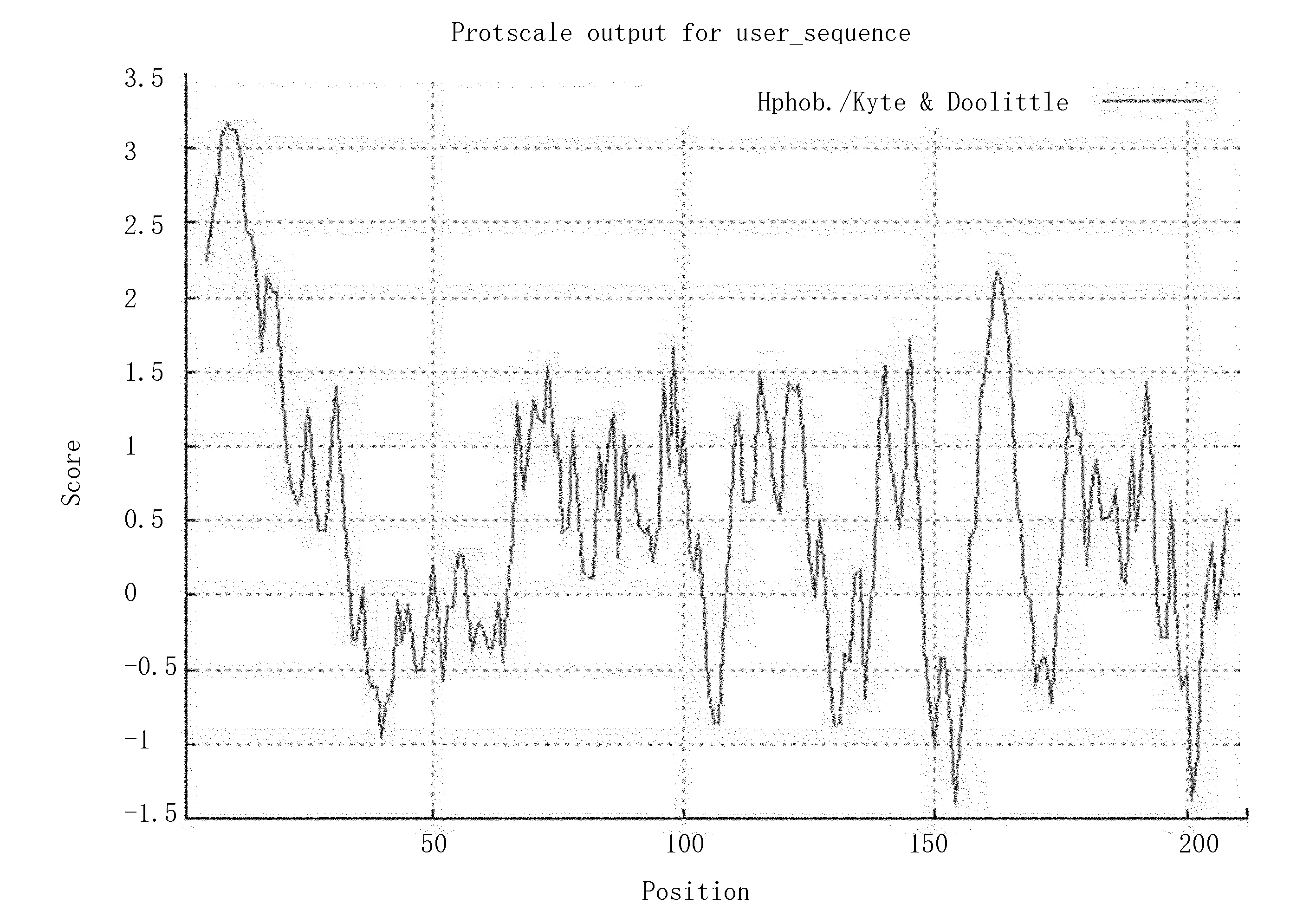
注:峰值<0,表示亲水性,峰值>0,表示疏水性。
图4Zmglp蛋白的亲疏水性及其区域
Fig.4 Hydrophilicity and region ofZmglpprotrin
2.2.2.2 玉米Zmglp蛋白质二级结构预测和分析
利用SOPMA预测玉米Zmglp蛋白的二级结构(图5),结果表明:α螺旋(Hh):62,29.25%,β折叠(Tt):26,12.26%,延伸链(Ee):58,27.36%,无规则卷曲(Cc):66,31.13%。
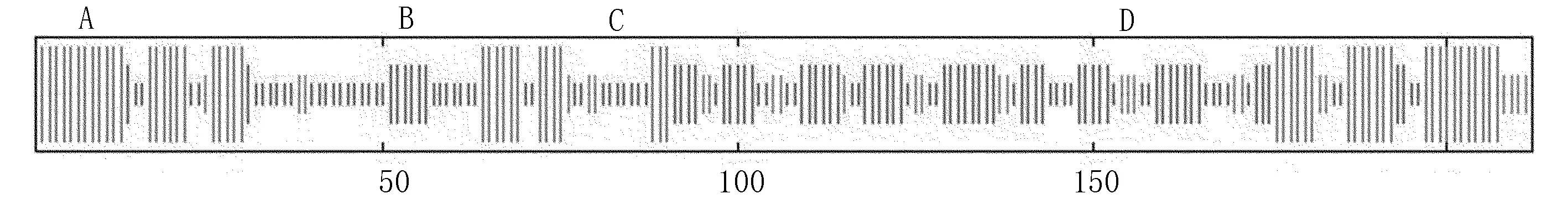
注:A为α-螺旋;B为延伸链;C为无规则卷曲;D为β转角
图5 Zmglp蛋白质的二级结构预测
Fig.5 Zmglp protein secondary structure prediction
2.2.2.3 玉米Zmglp蛋白质信号肽分析
根据信号肽序列特征,采用神经网络方法或隐马氏模型方法,根据物种的不同,对信号肽位置及切割位点进行预测。用SignalP对Zmglp进行信号肽的分析(图6),可更直观分析氨基酸的Score,可知信号肽存在几率是0.997 8,切割位点在22和23位氨基酸之间。
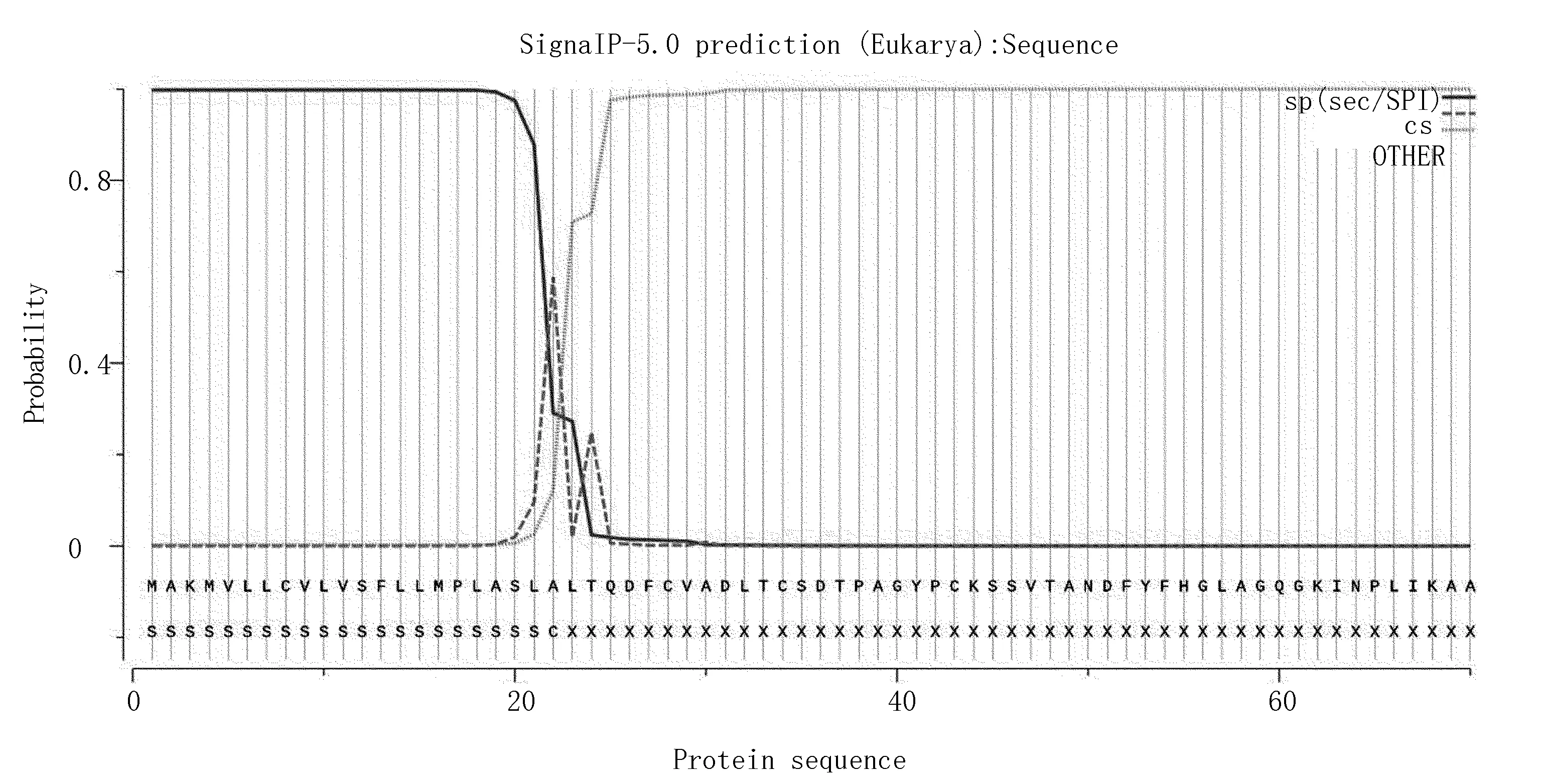
图6 Zmglp的信号肽分析
2.2.2.4 玉米Zmglp蛋白质的保守功能域分析
利用CD-Search软件分析蛋白质的保守域,结果表明:Zmglp蛋白具有1个cupin_OxOx保守功能域(图7),该功能域位于第20~208位氨基酸之间,属于Cupin类超级家族,具有草酸盐氧化酶活性。

图7 Zmglp的保守域及其功能
2.2.2.5 玉米Zmglp蛋白质翻译后修饰位点分析
用NetPhos3.1进行磷酸化位点分析结果显示:Zmglp蛋白磷酸化位点有10个(图8),其中,Ser 3个、Thr 5个、Tyr 2个,说明磷酸化位点修饰对Zmglp蛋白的结构或功能可能非常重要,这些磷酸化位点也可能参与该蛋白活性的调控。Ser磷酸化位点为Ser46、Ser128、Ser206;Thr磷酸化位点为Thr24、Thr33、Thr118、Thr130、Thr157;Tyr磷酸化位点为Tyr53、Tyr132。
用Yin0Yang1.2软件进行O-β-葡萄糖糖基化位点分析结果表明:Zmglp的O-β-葡萄糖糖基化位点有8个(图9),其中,Ser 3个,分别为Ser35、Ser45、Ser167;Thr 5个,分别为Thr72、Thr115、Thr118、Thr133、Thr157。
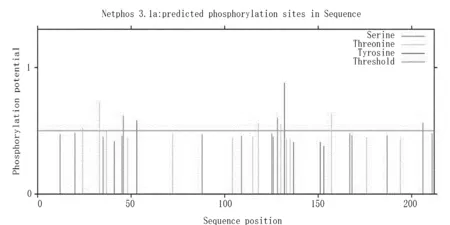
图8 Zmglp的蛋白磷酸化位点
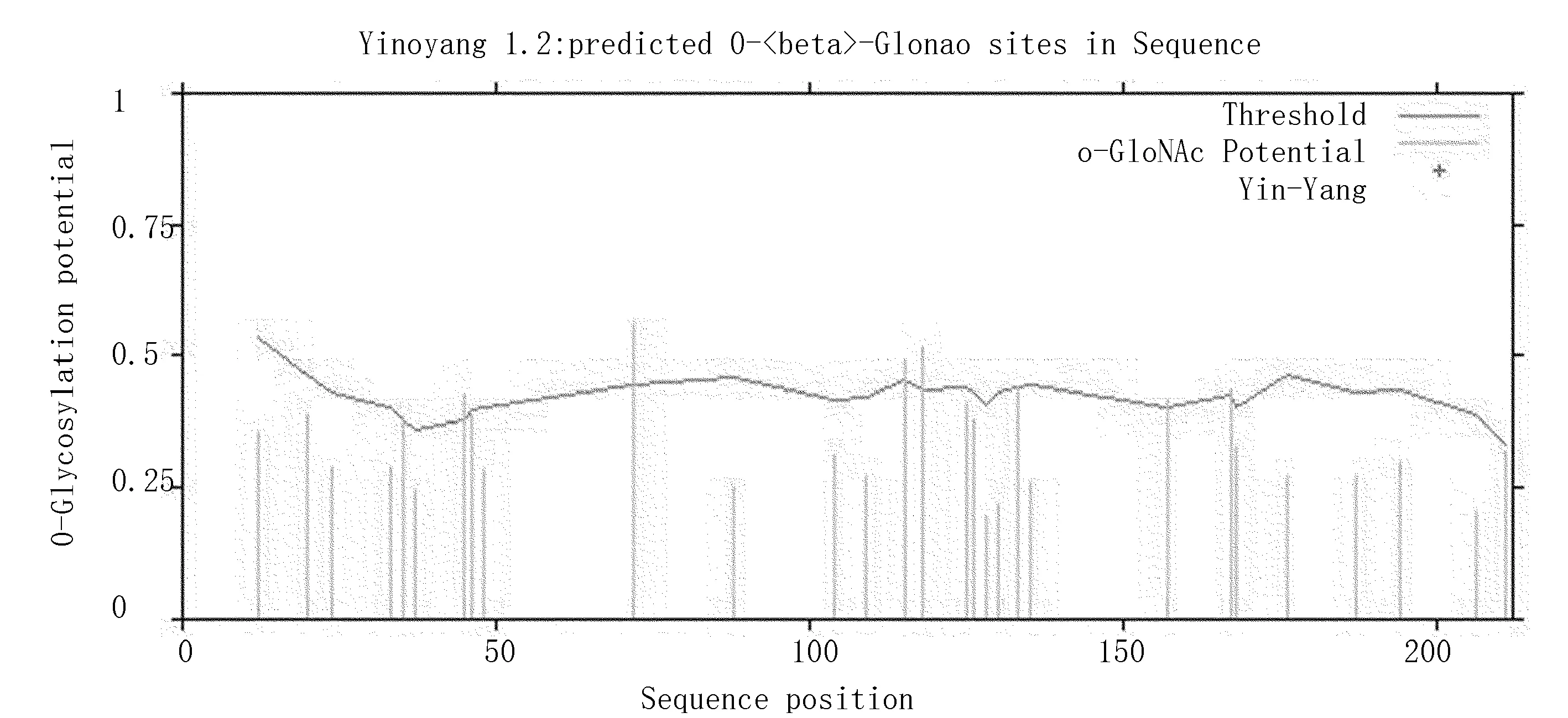
图9 Zmglp的O-β-葡萄糖糖基化位点
2.2.2.6 玉米Zmglp蛋白质跨膜区域与亚细胞定位预测
用TMHMM在线预测Zmglp蛋白质跨膜螺旋的结果表明:该基因编码蛋白在5~24个氨基酸位置之间有1个明显的跨膜区域,因此,预测Zmglp蛋白属于跨膜蛋白(图10),经Plant-mPLoc预测定位在细胞壁上。
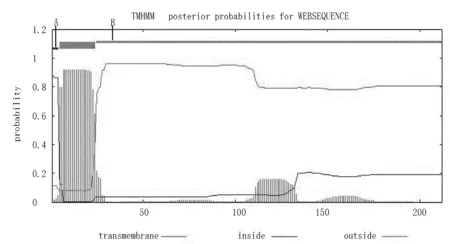
注:A表示伸入到细胞内的部分,B表示伸入到细胞膜外的部分
2.2.2.7 玉米Zmglp蛋白质三级结构分析
为了解玉米GLP蛋白的高级结构,用SWISS-MODEL同源建模软件预测蛋白质三级结构。此蛋白的空间结构主要是由α螺旋、β折叠、无规则卷曲和延伸链组成,与二级结构预测结果基本一致(图11)。
2.2.2.8 玉米Zmglp氨基酸的多序列对比和系统发育树分析
从UniProtKB蛋白质数据库中选取10种禾本科的GLP蛋白和玉米Zmglp蛋白进行氨基酸的多序列比对和系统发育树分析。运用DNAMAN软件对它们的氨基酸序列进行多重比对结果显示,氨基酸序列一致性为71.68%,由此可见,GLP蛋白在进化上比较保守。运用MEGA6.06软件绘制系统发育树(图12),结果表明:玉米(Zmglp Huangzao4)和双色高粱(C5Z116)、Dichanthelium oligosanthes(A0A1E5VJG5)、黍稷(A0A3L6RHB0)、Panicum hallii var. Hallii(A0A2T7D8A4)、小米(K3YJN4)为一支; 展颖野生稻(A0A0E0AWB2)、二穗短柄草(A0A2K2D1T6)、串珠节节麦(A0A453R4Z7)、小麦(A0A3B6SAY4)、大麦(F2E7Z5)为一支。
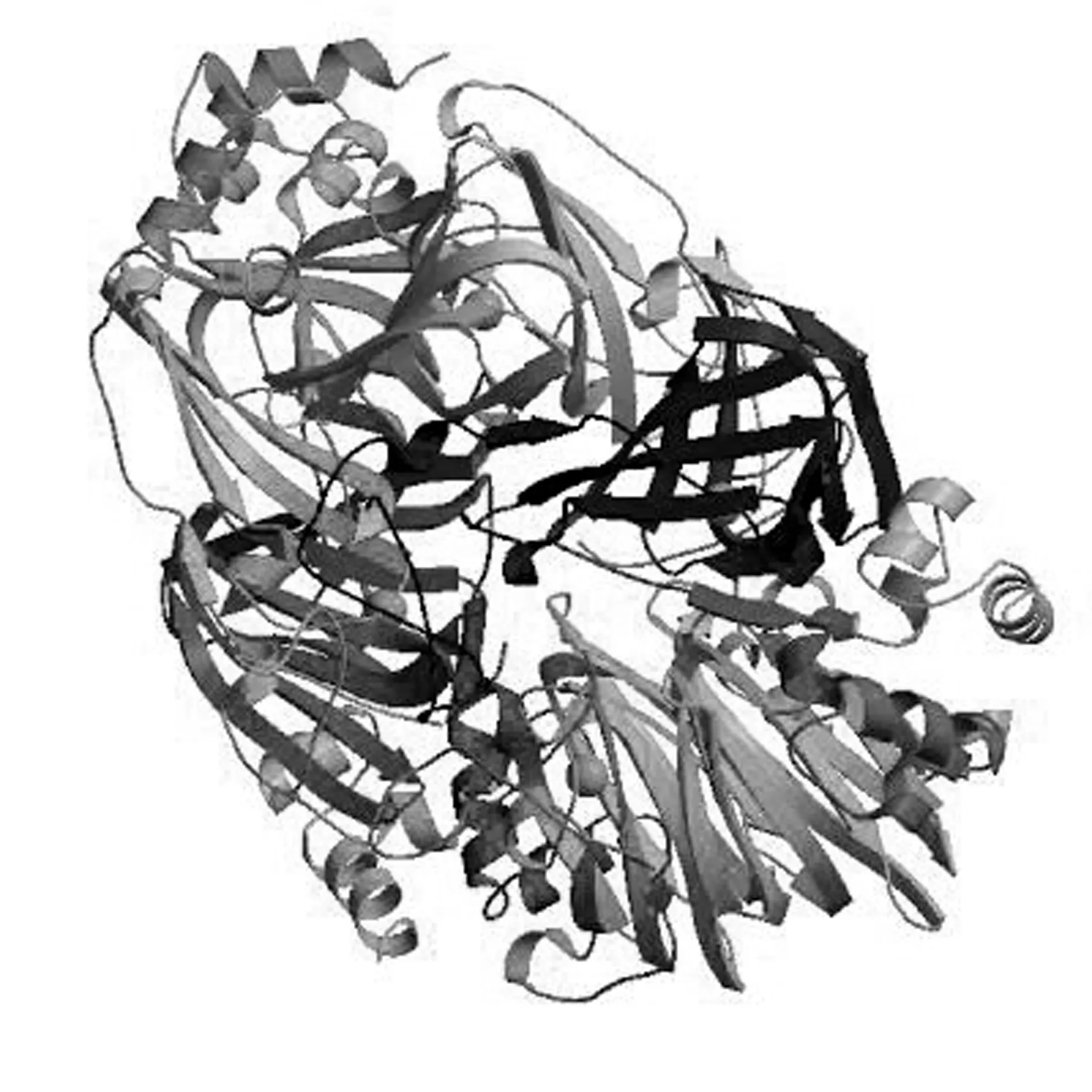
图11 Zmglp的三级结构预测
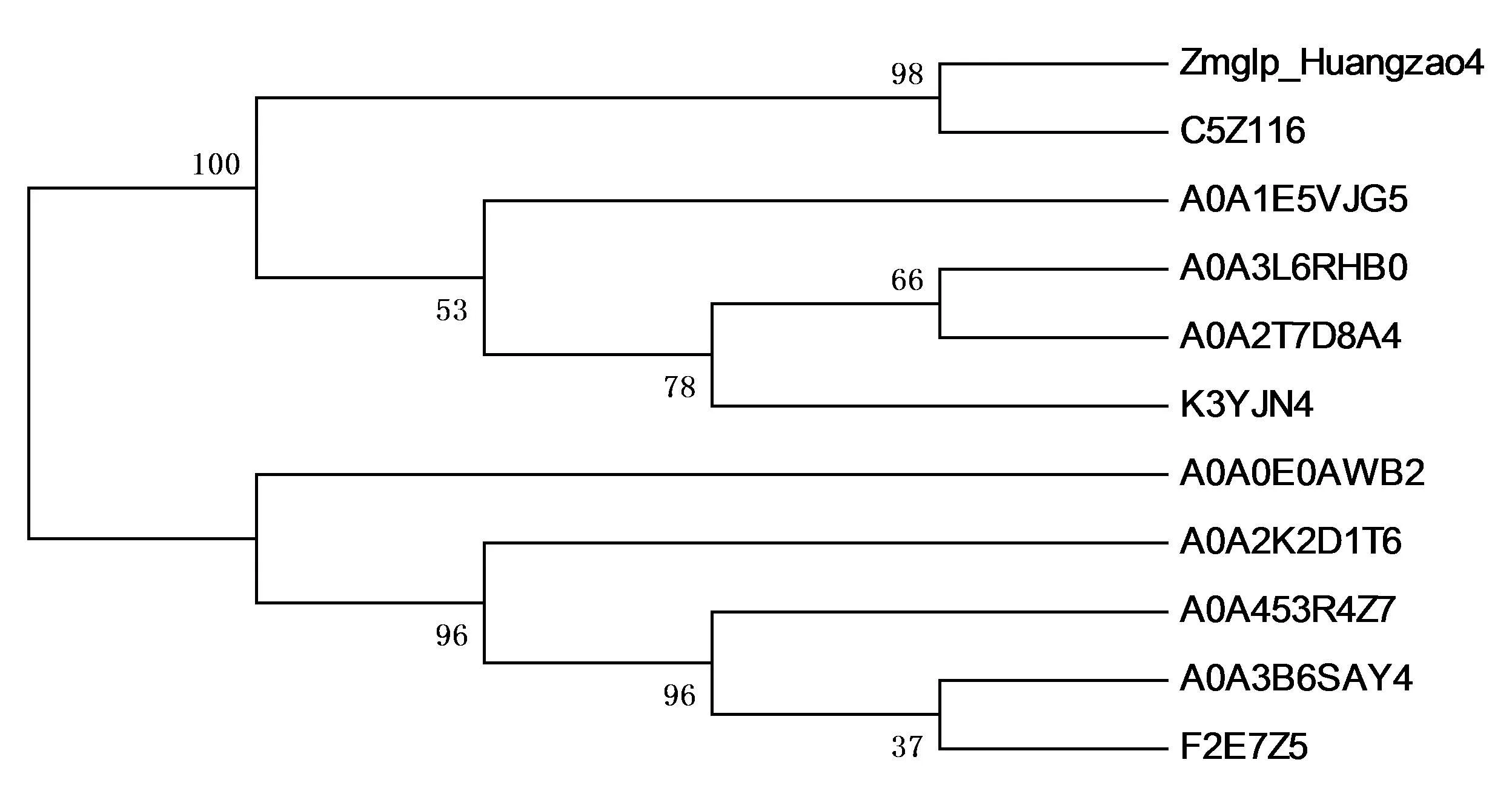
图12 玉米中Zmglp与其他物种GLP的进化树分析
3 讨论与结论
GLPs广泛分布在陆生植物中,且组成了一个庞大而功能各异的基因家族。据相关文献报道,萌发素和类萌发素蛋白在盐胁迫、干旱胁迫、抵御病原体以及重金属离子胁迫中起重要作用[12-15]。
在前期差异蛋白分析中发现,高抗玉米自交系鲁原92的GLP蛋白受弯孢菌侵染而特异性诱导[16]。该研究选用亲缘关系较远的2个亲本,即来自美国Lancaster群的抗弯孢菌自交系Mo17与中国培育的塘四平头群的感弯孢菌自交系黄早4为材料,克隆的Zmglp基因片段在ORF区段内只有2个碱基的同义突变,而在3’端存在大量碱基突变,表达的蛋白质含有8个糖基化与10个磷酸化位点,与禾本科植物的同源基因的比对结果显示,氨基酸序列一致性达到了71.68%,说明Zmglp蛋白在玉米甚至是禾本科中功能非常保守,也暗示了该基因可能在禾本科植物的生长发育中主要受抗病性相关miRNA调控表达量的变化以及蛋白质的糖基化与磷酸化的作用而起到抵抗生物与非生物胁迫的重要功能。
进一步研究发现,生物或非生物因素的水杨酸、乙烯以及ABA信号等途径都可以诱导GLPs基因的表达[17-21],在拟南芥、水稻、小麦等植物的GLPs基因的研究上,证明类萌发素蛋白在植物体内主要通过酶(OXO、SOD、ADPPase)、受体(ABP19/20)和结构蛋白发挥生理功能[22]。在该研究中,Zmglp蛋白的N端含有信号肽和跨膜域,亚细胞定位在细胞壁上,且含有Cupin超级家族的具有草酸盐氧化酶功能cupin_OxOx保守域,据此推测,Zmglp蛋白可穿过膜结构通过酶(OXO、SOD、ADPPase)、受体(ABP19/20)和结构蛋白共价结合在细胞壁上,并接受生物或非生物胁迫的信号,由cupin_OxOx功能域催化产生的H2O2与草酸钙降解释放的Ca2+一起达到较高浓度后介导细胞壁交联,并传递信号调控相关防御基因的表达,起到抵抗生物或非生物胁迫的作用。
猜你喜欢
分子催化(2022年1期)2022-11-02
中国种业(2022年8期)2022-08-19
波谱学杂志(2022年1期)2022-03-15
昆明医科大学学报(2022年1期)2022-02-28
种子(2021年8期)2021-09-23
昆明医科大学学报(2021年3期)2021-07-22
烟草科技(2021年6期)2021-06-24
北京农学院学报(2020年1期)2020-03-09
电脑知识与技术(2018年19期)2018-11-01
分析化学(2017年12期)2017-12-25
