
Cd胁迫下白桦光合及叶绿素含量的响应研究
2020-05-13 05:14赵串串温怀峰
陕西科技大学学报 2020年2期
赵串串, 温怀峰
(陕西科技大学 环境科学与工程学院, 陕西 西安 710021)
0 引言
2019年1月1日,伴随着《土壤污染防治法》的生效,土壤污染防治也将进入“新纪元”.研究发现木本植物具有生物量大、根系发达、对重金属耐性好、经济可行,抑制污染向食物链中转移等优势,将其应用于重金属污染土壤的修复具有良好的前景[1].重金属胁迫对植物的危害是在各种环境因素(也包括植物本身)的综合作用下进行的,即使是同一种金属元素,同一浓度,在不同的环境条件下,对植物的危害也会表现出明显的差异[2-4].镉(Cd)胁迫对木本植物光合作用的影响可能存在较为复杂的调控机制[5].冯建灿等[6]研究认为叶绿素荧光动力学参数是评价植物逆境生理的有效方法.
研究表明,桦树对逆境胁迫具有较好的适应性和较强的调控能力[7],目前研究主要集中在盐胁迫[8],干旱/涝渍胁迫[9,10]及高/低温胁迫[11,12]等方面.重金属胁迫对白桦(BetulaPlatyphyllaSuk.)生理指标影响有待进一步揭示.本文以2 年生白桦幼苗为试材,通过盆栽试验,基于叶绿素荧光技术,研究Cd胁迫对白桦幼苗光合荧光特性及叶绿素含量的影响,探究白桦对高浓度Cd胁迫的适应性及耐受性.
1 实验部分
1.1 研究对象
1.1.1 幼苗
本研究以2年生木本植物白桦幼苗为对象,白桦幼苗取自青海省林业厅造林育苗苗圃.
1.1.2 土壤基质
以土壤、蛭石、蚯蚓粪(V∶V∶V=3∶1∶1)混合物为盆栽基质.将CdCl2·2.5H2O(AR)与去离子水配成母液,然后稀释成所需的处理浓度,施入相应的盆栽土壤中.将白桦幼苗栽种于直径15 cm、高25 cm的塑料盆中,土壤质量约5 kg,盆底放置两层塑料纸.根据课题组前期实验及文献资料,张春燕等[13]报道了废水灌溉土壤中Cd含量为 10.69~173.64 mg/kg.韦月越[14]研究报道了某矿山土壤Cd含量最高达448.50 mg/kg.杨园等[15]研究结果表明土壤中Cd浓度在150 mg/kg使杨树叶绿体内部的基粒结构发生改变,从而对光合作用产生抑制.本文土壤Cd浓度设定为0,50 mg/kg,100 mg/kg,200 mg/kg 4组,依次记为control,50 Cd,100 Cd,200 Cd,每组设3个重复.
1.2 实验仪器
LI-6400XT (LI-COR Inc.,Lincoln,Nebraska,USA) 便携式光合系统;752紫外-可见光分光光度计 (上海舜宇恒平仪器有限公司).
1.3 测定方法
1.3.1 白桦光合荧光参数测定
采用LI-6400XT测定白桦叶片光合作用气体交换参数,光合光响应曲线和叶绿素荧光参数.胁迫前于2017年6月9日测定,实验启动后分别于胁迫3周(wks)、5 wks后于09∶00~11∶30间分别进行测定.实验结束后测定白桦叶绿素含量.
光合荧光参数及叶片光合作用对光合有效辐射(PAR)响应曲线测定,采用LI-6400XT.测定参数:最小荧光产量(F0)、最大荧光产量(Fm)、PSⅡ原初反应的最大量子效率(Fv/Fm)[16]及潜在活性(Fv/F0)、有效光量子化学效率(Fv′/Fm′),PSⅡ反应中心实际光量子化学效率(ΦPSⅡ),表观电子传递速率(ETR),及非光化学淬灭(NPQ),净光合速率(Pn),气孔导度(Gs),胞间CO2浓度(Ci),蒸腾速率(Tr),计算气孔限制值(Ls)[17].其中,Ls的计算公式为:Ls=1-Ci/Ca式中Ca为大气CO2浓度.PAR梯度设定为2 000μmol/(m2·s)、1 800μmol/(m2·s)、1 500μmol/(m2·s)、1 200μmol/(m2·s)、900μmol/(m2·s)、700μmol/(m2·s)、500μmol/(m2·s)、400μmol/(m2·s)、300μmol/(m2·s)、200μmol/(m2·s)、100μmol/(m2·s)、75μmol/(m2·s)、50μmol/(m2·s)、25μmol/(m2·s)和0μmol/(m2·s).使用直角双曲线修正模型对测得的光响应曲线进行拟合[18].试验期间,环境温度为28.34±2.03 ℃,大气CO2浓度为392.04±16.35μmol/(m2·s).
1.3.2 叶绿素含量测定
参考胡秉芬等[19]的方法,采用95%乙醇溶液为提取剂,研磨、浸提后,取上清液其在波长为663 nm、645 nm处的吸光度.
1.4 数据统计与分析
数据表示为平均值±标准误差,经方差分析后进行多重比较(Tukey′s 法,α=0.05).在SPSS 20.0中进行双因素方差分析(Two-wayANOVA)来揭示Cd处理与胁迫周数及交互作用对白桦苗木各光合与叶绿素荧光指标的影响,Cd浓度和暴露于Cd胁迫的周数作为每周测量数据的自变量.用Origin 8.6绘图软件作图.
2 结果与讨论
2.1 Cd胁迫对白桦叶片叶绿素含量的影响
Cd胁迫下白桦叶片的叶绿素含量如图1所示.由图1可知,Cd胁迫显著降低了叶绿素含量(P<0.05).其中,叶绿素a含量的变化最为显著,总叶绿素含量与叶绿素a/b的降低主要由于叶绿素a含量的减少造成.
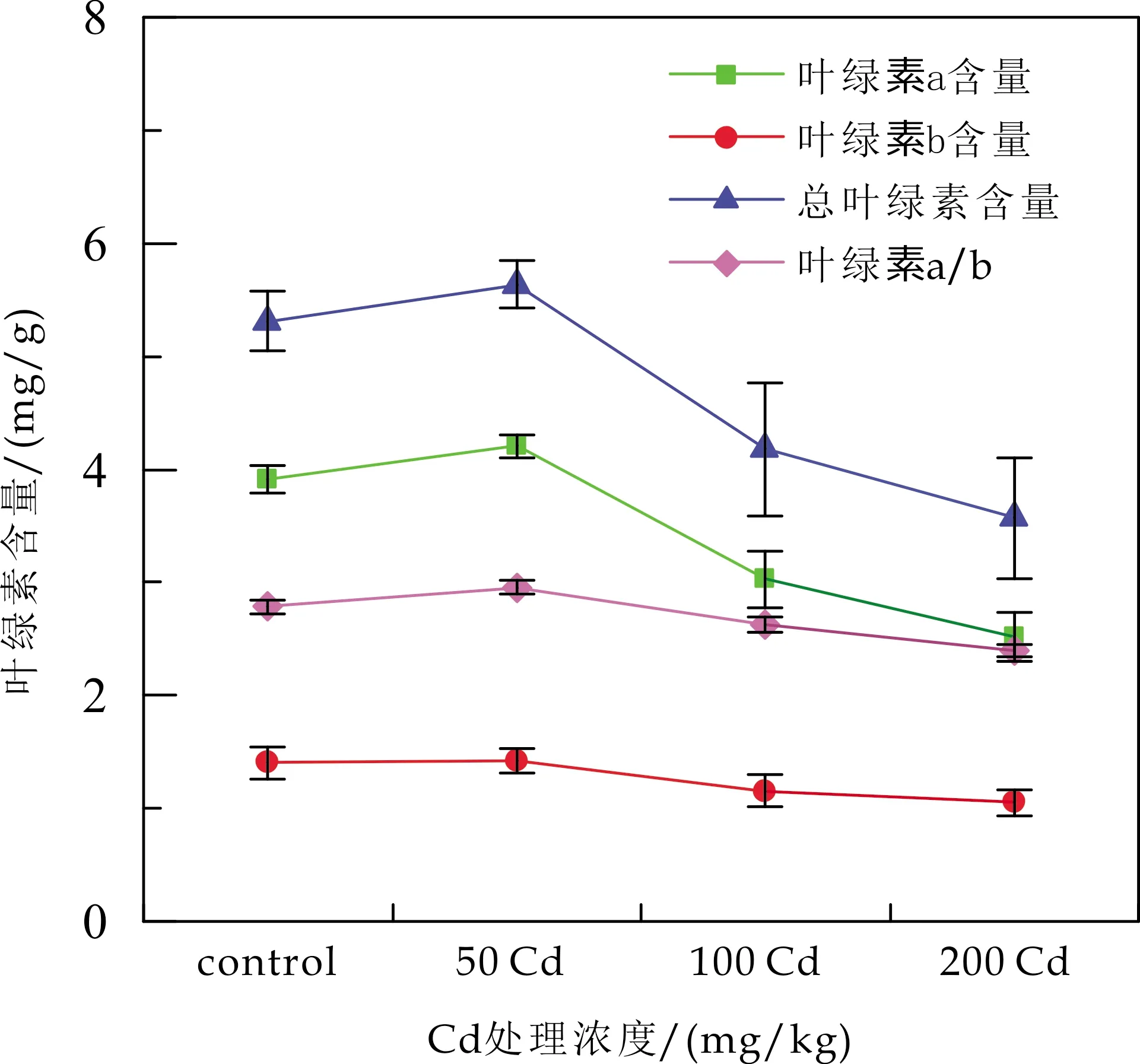
图1 Cd胁迫对白桦叶绿素含量的影响
胁迫5 wks后,w(Cd)为50 mg/kg 时,与control相比,白桦叶片的叶绿素a含量增加了7.67%;w(Cd)为100和200 mg/kg时,白桦叶片的叶绿素a含量分别减小了22.51%和35.55%,同时总叶绿素含量和叶绿素a/b也分别降低了21.28%、32.77%和5.40%、13.67%.表明Cd胁迫会影响叶绿素的含量,主要通过在类囊体膜上大量累积,从而破坏了叶绿体结构以及叶绿素合成酶的活性[20],从而导致叶绿素含量的降低.当w(Cd)为50 mg/kg 时,与胁迫前相比,胁迫3~5 wks后,白桦的Pn下降后上升;同时,Gs、Ci也显著增大,更多CO2通过气孔进入叶肉细胞,叶绿素的含量上升,说明白桦叶绿素合成的阻碍得到缓解,刺激白桦光合作用增强,使Pn上升[21].
2.2 Cd胁迫对白桦苗木光合特性的影响
Cd胁迫下白桦苗木气体交换参数测定如表1所示.由表1可知,与胁迫前相比,Cd胁迫显著降低了白桦的Pn、Gs、Tr及Ls;同时,不同浓度的Cd处理基本使白桦的Pn、Gs、Tr及Ls表现出先减后增的趋势.当w(Cd)为50 mg/kg时,胁迫5 wks后,与胁迫3 wks后相比,白桦的Pn上升到胁迫前的97.70%,且与胁迫前无显著差异;与胁迫5 wks 后的control相比,白桦的Pn增加了11.84%.
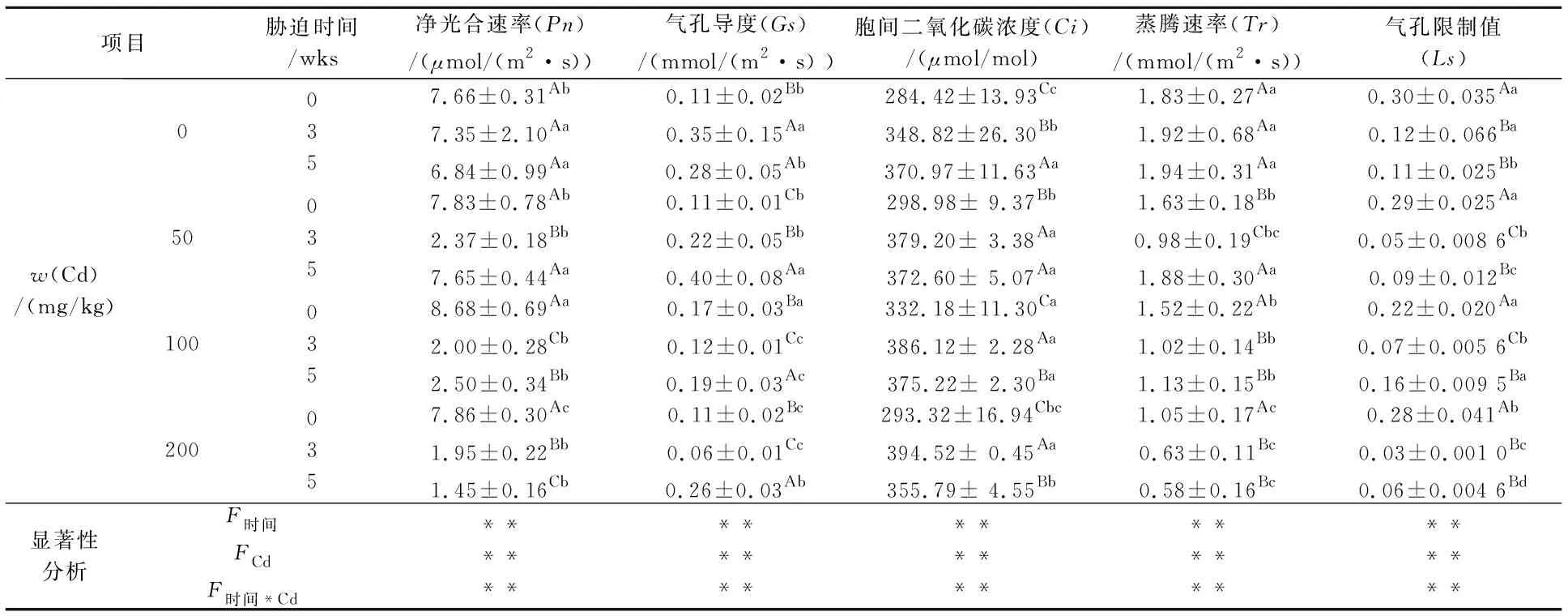
表1 Cd胁迫对白桦苗木气体交换参数的影响
注:A、B、C表示同一Cd处理下不同胁迫时间对白桦的影响在0.05水平上显著差异;a、b、c表示相同胁迫时间下不同Cd处理对白桦的影响在0.05水平上显著差异.**表示P<0.01.F时间,时间效应;FCd,Cd效应;F时间×cd,时间与Cd的交互效应.
双因素方差分析表明,胁迫时间与Cd作为独立因子时均显著影响了白桦叶片的Pn、Gs、Ci和Tr,当w(Cd)为100 mg/kg 时,交互条件下胁迫时间显著促进了Pn、Gs、Tr及Ls的回升,降低了Ci值;而当w(Cd)为200 mg/kg 时,胁迫显著抑制了白桦的光合作用,使叶片的Pn、Ci及Tr持续降低.
Cd胁迫下白桦叶片的Pn、Gs和Ls均呈显著下降(P<0.05)的趋势,Ci上升,表明白桦的光合作用主要受非气孔因素的限制,与前人的研究结论一致[17,22].w(Cd)为100 mg/kg时,胁迫全周期内,白桦的Pn下降后出现回升的趋势,表明该胁迫下白桦通过反馈机制调节产生了适应性,从而减弱了Cd对白桦光合作用的抑制[23].随着胁迫周期的延长,叶绿素含量以及叶绿素a/b显著减少,说明Cd胁迫对白桦叶绿素a的影响比叶绿素b大;Ci值呈现先增大后减小的趋势,可能由于初期Cd胁迫破坏了叶绿体的结构或使叶绿体酶活性发生变化,进而影响叶绿体的合成及光合作用,导致Ci增大[24];而Gs和Ci呈相反的趋势,胁迫5 wks 后,Gs增大,Ci反而减小,白桦对Cd胁迫产生了适应性.
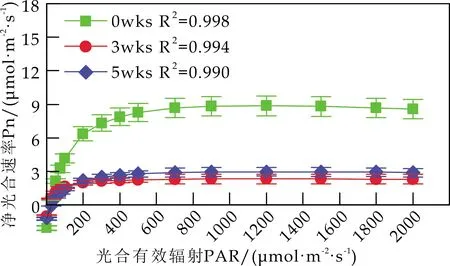
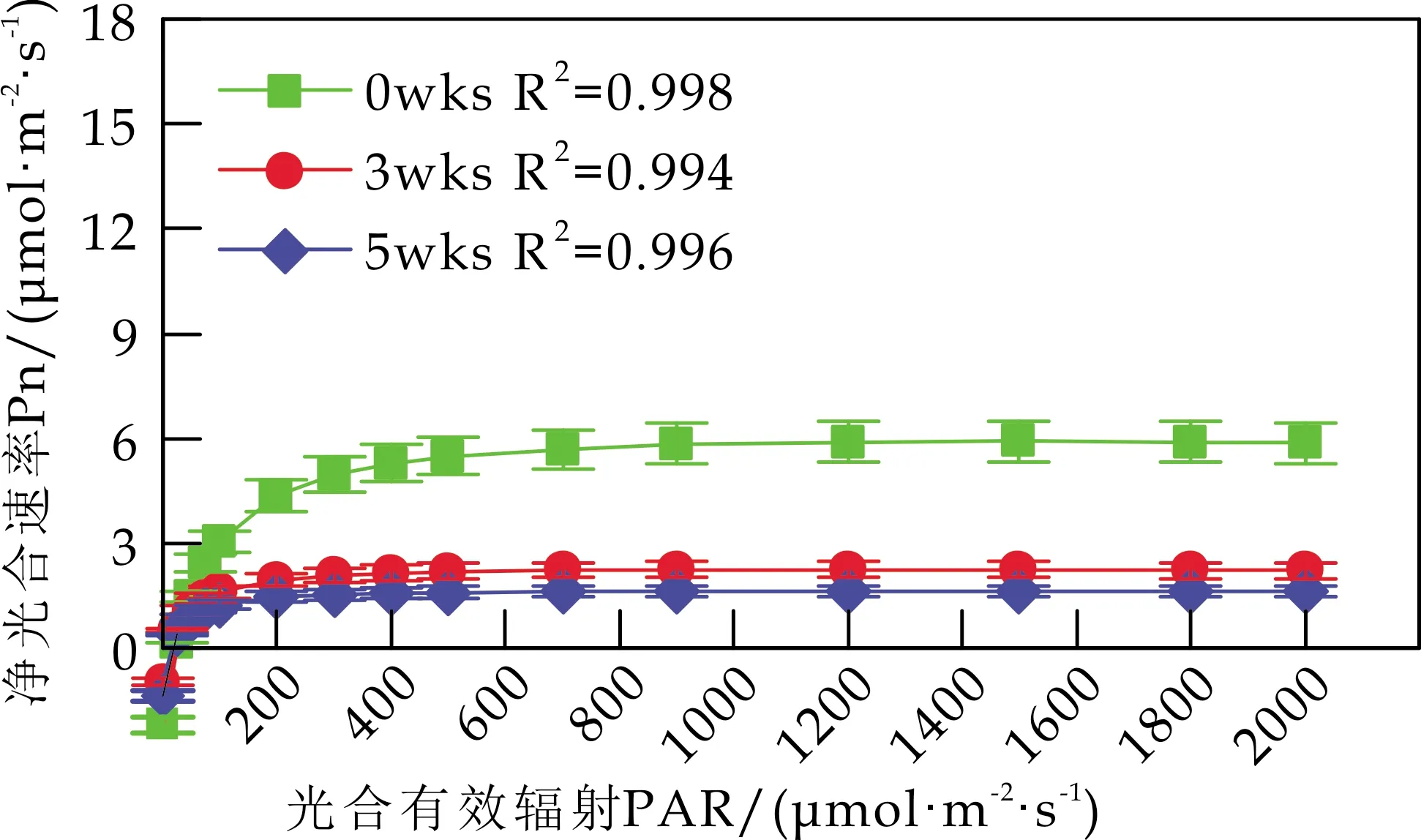
2.3 Cd胁迫对白桦苗木光合-光响应特性的影响
各组白桦叶片Pn的光响应曲线见图2.由图2可知,随着光强的增大,白桦的Pn在PAR<200μmol/(m2·s) 时迅速上升;当PAR达到700μmol/(m2·s)时,白桦的Pn逐渐平稳,达到饱和.不同浓度的Cd处理显著降低了白桦的光合作用强度,并使白桦叶片更容易达到光饱和状态;相同浓度Cd处理下,胁迫时间也显著抑制了白桦的Pn对光强的响应.
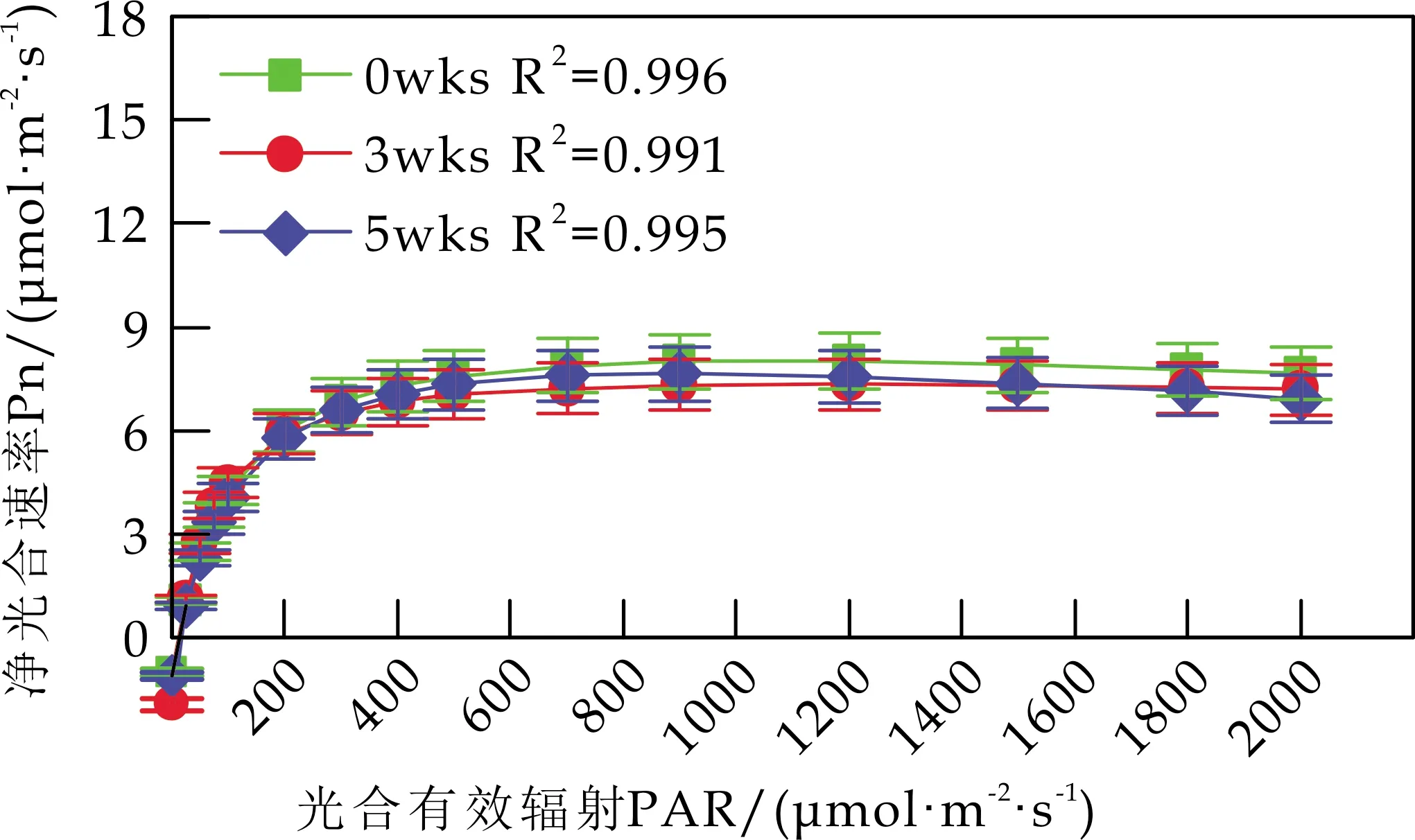
(a)control
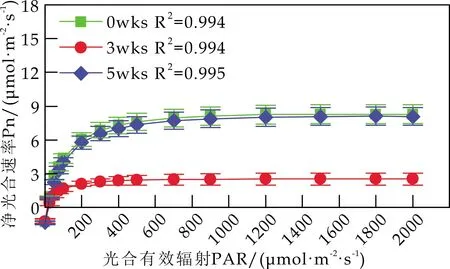
(b)50 mg/kg
(c)100 mg/kg
(d)200 mg/kg图2 Cd胁迫对白桦叶片光响应曲线的影响
由图2(b)可知,w(Cd)为50 mg/kg 时,胁迫5 wks后,白桦的Pn对光强的响应与胁迫前无显著差异.由图2(c)可知,w(Cd)为100 mg/kg 时,与胁迫3 wks后相比,胁迫5 wks后,白桦的Pn随着光强的增大,有小幅度上升.由图2(d)可知,w(Cd)为200 mg/kg 时,胁迫5 wks后,白桦的Pn对光强的响应持续减弱,表明高浓度Cd胁迫降低白桦对光的利用能力,分析原因是高浓度Cd胁迫降低了叶绿素的含量,尤其是叶绿素a的含量.
2.4 Cd胁迫对白桦苗木荧光特性的影响
Cd胁迫下白桦苗木叶绿素荧光参数见表2.由表2可知,与胁迫前相比,Cd胁迫显著降低了白桦的Fm、Fv/Fm及Fv/F0;随着胁迫时间的延长,不同浓度的Cd处理使白桦的Fm、Fv/Fm和Fv/F0持续减小.w(Cd)为50 mg/kg时,胁迫3 wks后,白桦叶片的F0达到最大值为535.28;胁迫5 wks后,F0与胁迫前无显著差异,Fm、Fv/Fm及Fv/F0达到胁迫前的95.33%、98.73%和93.72%.其中Fv/Fm及Fv/F0与control相比增加了1.27%和4.13%.
双因素方差分析表明,胁迫时间与Cd作为独立因子时均显著影响了白桦叶片的F0、Fm、Fv/Fm和Fv/F0.w(Cd)为100 mg/kg时,交互条件下胁迫时间显著促使了F0的上升,降低了Fm、Fv/Fm和Fv/F0值.w(Cd)为200 mg/kg 时,胁迫时间使叶片的F0、Fm、Fv/Fm和Fv/F0显著降低.
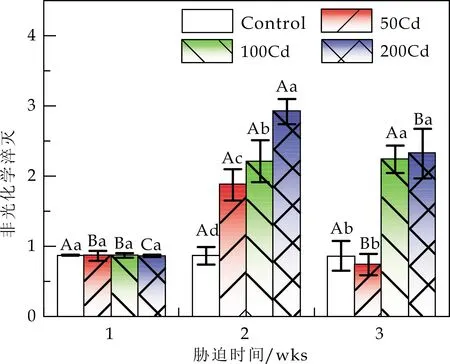
Cd胁迫对白桦光下荧光参数的影响见图3.由图3可知,胁迫全周期内,Cd胁迫显著降低了白桦的Fv′/Fm′、ΦPSⅡ及ETR,且不同浓度的Cd处理基本使白桦的Fv′/Fm′、ΦPSⅡ和ETR持续减小;同时白桦的NPQ值(除50Cd外)显著增加.非光化学淬灭(NPQ)反映PSⅡ反应中心的关闭程度[25,26],是植物免受光损失的重要自我保护机制[27].Andresen等[28]的研究表明,当胁迫强度超出一定阈值,这种保护机制的作用就会受到局限.如图3(d)所示,当w(Cd)为50 mg/kg时,胁迫5 wks后,与胁迫3 wks后相比,白桦叶片的NPQ显著降低 (降至最小值为0.74),占胁迫前该处理的94.27%;而Fv′/Fm′、ΦPSⅡ及ETR均显著升高,分别占胁迫前的112.12%、103.49%和105.94%.w(Cd)为100 mg/kg时,胁迫显著促进了NPQ的上升,说明白桦过剩光能主要以热耗散的形式散失,保护自身光合器官的生理活性[29];w(Cd)为200 mg/kg时,胁迫使叶片的Fv′/Fm′、ΦPSⅡ、NPQ及ETR值显著降低,说明白桦已丧失部分热耗散能力,NPQ保护机制作用受限.
注:A、B、C表示同一Cd处理下不同胁迫时间对白桦的影响在0.05水平上显著差异;a、b、c表示相同胁迫时间下不同Cd处理对白桦的影响在0.05水平上显著差异.*、**分别表示0.01 (c)表观电子传递速率(ETR) (d)非光化学淬灭(NPQ)图3 Cd胁迫对白桦光下荧光参数的影响 PSⅡ原初反应的最大量子效率Fv/Fm、有效光量子化学效率Fv′/Fm′和PSⅡ实际光化学量子效率ΦPSⅡ是判断植物是否受到光抑制的重要指标.Fv′/Fm′反映开放的PSⅡ反应中心的有效原初光能捕获效率[30].ΦPSⅡ反映PSⅡ反应中心在有部分关闭情况下的实际原初光能捕获效率,即实际光能捕获能力.Guo等[31]和Semida等[32]研究发现,Fv/Fm反映的是最大PSⅡ光能转化效率,如果Fv/Fm小于0.80,说明叶片的PSⅡ反应中心潜在光合能力和潜在活性受损.Cd胁迫会导致ETR显著降低(P<0.05)[33],阻碍电子传递,进而抑制光合作用. 本研究中,胁迫周期内,w(Cd)为50 mg/kg 时,Fv/Fm、Fv′/Fm′和ΦPSⅡ无显著差异,分别为0.80、0.71和0.67,而ETR则显著升高(P<0.05);与control相比,Fv/Fm、Fv′/Fm′和ΦPSⅡ均有所提升,表明50 mg·kg-1的Cd处理促进了白桦叶片的光捕获能力及光合作用过程中的电子传递速率,从而促进了白桦光合作用.w(Cd)为200 mg/kg 时,Fv/Fm仍能保持较高水平为0.73,说明Cd胁迫对白桦潜在光合能力以及PSⅡ反应中心活性的影响较小.而Fv′/Fm′和ΦPSⅡ显著减小,分别为0.43和0.30,说明Cd胁迫对白桦实际光合能力的影响是存在的,可能由于高浓度Cd胁迫降低了叶绿素含量,直接抑制光合作用. (1)w(Cd)为200 mg/kg时,胁迫使白桦叶片的叶绿素含量显著减少(P<0.05),电子传递受阻,光捕获能力减弱,光能利用能力减弱,白桦热耗散能力被破坏,从而抑制其光合作用. (2)w(Cd)为50 mg/kg时,持续5 wks后,白桦的Pn与control相比增长了11.84%,而w(Cd)为100和200 mg/kg时,Pn显著降低,表明白桦对低浓度Cd暴露表现出兴奋效应. (3)w(Cd)低于100 mg/kg时,白桦叶片的ΦPSII、NPQ和ETR等参数基本维持在一定的范围,说明白桦可以依靠自身抗逆性缓解Cd对反应中心活性的抑制作用,使光合电子链的传递速率得到恢复进而缓解Cd胁迫对白桦的光合损害,表明白桦对Cd胁迫具有一定的耐受性.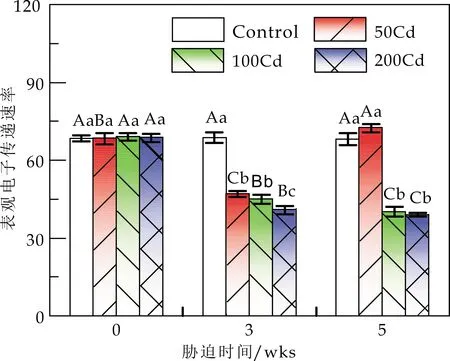
3 结论
猜你喜欢
阅读(科学探秘)(2020年8期)2020-11-06
文苑(2019年20期)2019-11-20
文苑·经典美文(2019年10期)2019-10-16
绿色科技(2019年2期)2019-05-21
女性天地(2016年10期)2017-04-25
Coco薇(2016年5期)2016-06-03
华人时刊(2016年16期)2016-04-05
考试周刊(2016年6期)2016-03-11
少儿科学周刊·儿童版(2015年10期)2015-11-07
食品工业科技(2014年6期)2014-05-10
