
多环境下水稻株型相关性状QTL解析
2020-05-11 03:11刘化龙杨洛淼徐善斌刘华东邹德堂
东北农业大学学报 2020年4期
刘化龙,杨洛淼,徐善斌,刘华东,邹德堂
(寒地粮食作物种质创新与生理生态教育部重点实验室,东北农业大学,哈尔滨 150030)
水稻是世界重要粮食作物,理想株型可促进水稻充分利用光合,对其生长发育和籽粒产量形成具有重要作用。水稻株型涉及分蘖数、分蘖角度、株高等指标。株高决定水稻生物量和收获指数[1],较高的株高改善水稻通风和叶片光照条件,但易引起倒伏,反之则降低光合效率[2]。分蘖形态(分蘖数、分蘖夹角等)影响水稻生长空间、光合作用及产量,且受环境影响较大。因此,水稻株型构成因素需达到平衡点才可保障产量潜力,发掘具有携带优异株型遗传位点的水稻株型育种工作具有重要意义。
水稻株高是受多基因调控的数量性状,目前已有1 000多个和株高相关的QTL被报道,已克隆株高基因超过100 个。分蘖角度既受主效基因调控,也受微效多基因调控,在植物数量遗传研究中,大量和分蘖角度有关的主效基因被报道。la是首个被报道与水稻分蘖夹角有关的基因[3],随后又报道有er等基因[4]。与分蘖数相关基因目前已克隆不足10个,MOC1是单蘖基因[5],而TAD1与MOC1共表达,促进分蘖发生[6],另外,D27、D10和D17参与独脚金内酯合成,D3 和HTD2 则通过对独脚金内酯信号组分作用促进分蘖[7-8]。虽部分基因的功能得到验证,但水稻株型塑造需从株高、分蘖数、分蘖角度等性状共同展开,目前同时控制不同株型相关性状表达的基因尚未克隆。因此需要不断利用新材料挖掘具有连锁遗传的多效性区间,找到株型相关性状的热点区间,提高水稻株型改良效率。
水稻株型建成是多基因共同协作的结果[9],利用遗传背景一致材料对不同株型性状同时开展研究,可从遗传图谱上获得多个连锁位点。同时,多环境下表型鉴定有助于QTL 稳定性检测,对于水稻株型变异位点挖掘具有重要意义。因此,本研究利用重组自交系群体,通过多年多点环境鉴定,研究包括株高、分蘖数、分蘖角度在内的3个株型相关性状,并作QTL 和QTL×环境分析,通过性状和遗传相关联合分析,旨在发掘同时控制多个株型相关性状的稳定遗传位点,为水稻理想株型育种及相关位点精细定位奠定理论基础。
1 材料与方法
1.1 群体材料及遗传图谱
研究所选材料以空育131为父本,东农422为母本,配制重组自交系,其中空育131株型为矮秆紧凑型,东农422为高秆披散型。通过构建和改良群体遗传图谱,最终有155个SSR标记覆盖该群体190个株系。
1.2 田间种植及调查
将群体及亲本材料分别于2016、2017 和2018年在海南崖城南繁基地(Hainan)和哈尔滨东北农业大学阿城实验实习基地(Harbin)种植。共6 个环境,定义Hainan 三年试验环境为E1(2016 年夏)、E2(2017 年夏)和E3(2018 年夏),Harbin 三年试验环境为E4(2016 年春)、E5(2017 年春)和E6(2018年春)。其中,Hainan 采用水育苗移栽,Harbin 采用旱育苗移栽。插秧规格为行长2 m,2 行区,株距10 cm,行距30 cm。水稻返青结束进入分蘖期后,调查每个株系分蘖动态及株高动态,采用挂牌方式从每个株系中选取均匀一致的3 个植株挂牌,每隔1 周调查1 次分蘖数(Tillering number,TN)、分蘖角度(Tillering angle,TA)及株高(Plant height,PH),直至穗抽出后,停止调查,此时分蘖数、株高和分蘖夹角稳定,最后1次调查数据用于分析。分蘖数以总分蘖计算,分蘖角度用量角器测量,测量选取同一株系中距离最远的两个分蘖间夹角,测量角度除以2即为分蘖角;株高测量以根基部到主茎倒一叶叶尖距离为准。
1.3 表型分析与统计
利用Sigmaplot 12.5 分析性状相关性及描述性统计,利用SPSS 18.0 软件一般线性模型(GLM),以基因型和环境为固定因子,性状为因变量作方差分析,作基因型方差(σ2G)、基因型×环境(σ2GE)和误差方差(σ2E)评价,参照Hallauer 等方法计算广义遗传力[10],计算公式为:
式中,n表示环境数量,r表示重复数。
1.4 QTL及环境互作分析
利用ICIMapping 4.2 完备区间作图法(Inclusive composite interval mapping,ICIM)分析加性QTL,以LOD值≥3.0作为QTL存在最小阈值,基因×环境互作利用QTL Network 2.0 混合线性模型复合区间作 图 法(Multi-model composite interval mapping,MCIM)分析。
不同性状QTL 之间相关性利用超几何概率函数计算[11]。
其中,n-遗传图谱长度2倍与两性状QTL置信区间平均数之和比值,m-两性状重合QTL 数目(两性状QTL置信区间大于50%重叠QTL数目),l-QTL数目较多性状检测到QTL数目,s-QTL数目较少性状检测到QTL数目[12]。
2 结果与分析
2.1 表型变异
由表1可知,通过在哈尔滨和海南连续3年表型鉴定可知,双亲在株高、分蘖数和分蘖角上存在显著差异,且单个性状在6 个环境中差异不显著,表明环境因素对双亲性状影响较小。母本东农422 株高和分蘖角度高于父本空育131,分蘖数低于空育131,二者互为两种株型,空育131 株型偏紧凑,东农422株型较散。由此可知,该双亲衍生后代群体适于水稻株型相关性状遗传解析。后代群体3 个性状在6 个环境中均值介于亲本之间,从RIL 表型变异范围可知,极端值介于双亲之外,表现超亲现象,变异较明显。偏度值和峰度值绝对值小于1,表明3 个性状在6 个环境下表型分布呈近似正态分布,符合数量性状典型特征,适于QTL分析。遗传力分析表明,性状表型变异主要由基因方差决定,环境因素对表型影响较小。
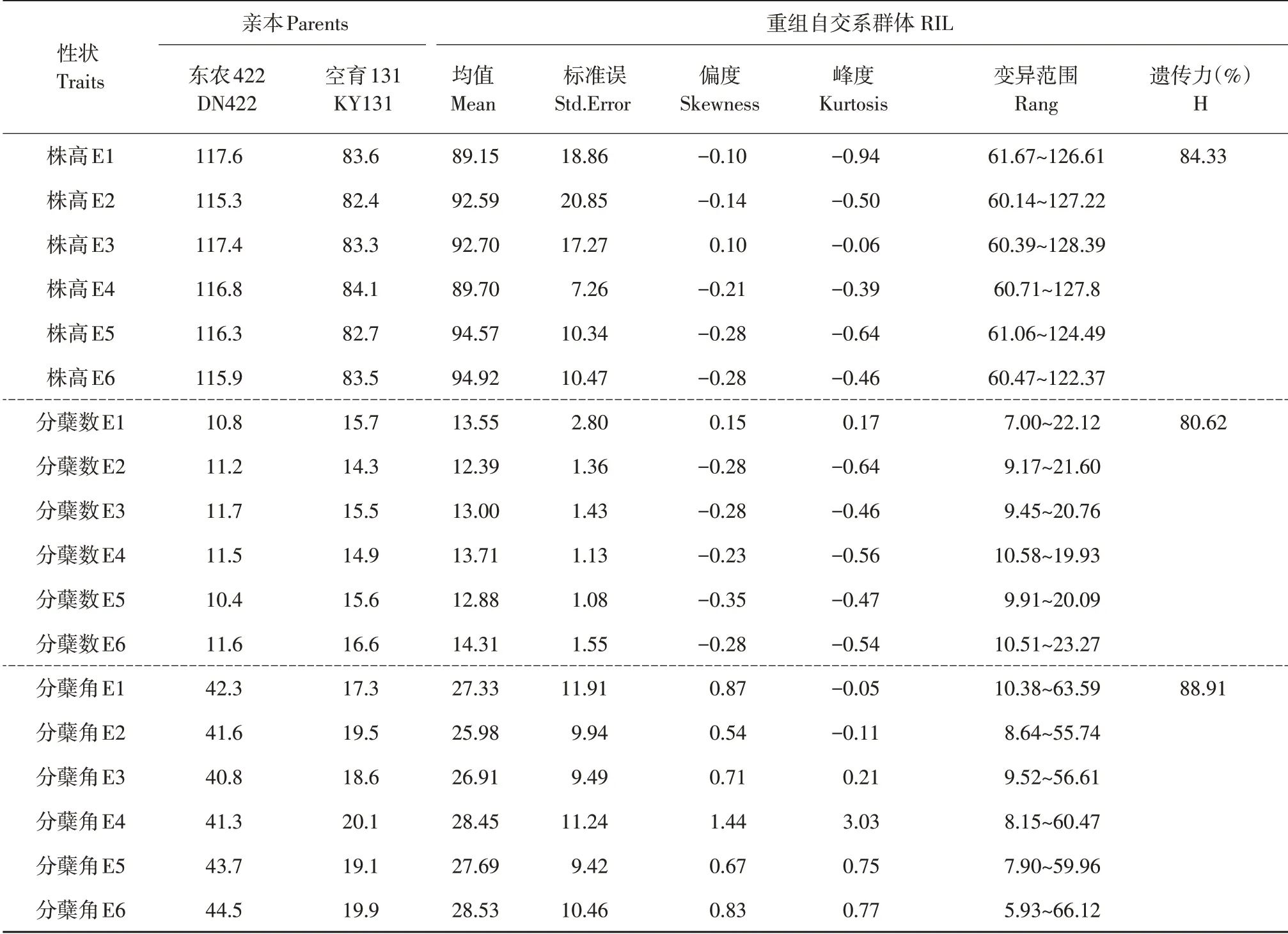
表1 株高、分蘖数和分蘖角度在6个环境下表型变异Table 1 Phenotypic variation of plant height,tiller number and tiller angle in six environments
由图1可知,株高和分蘖数在6个环境下均存在显著正相关关系,相关系数分别为0.206、0.443、0.540、0.683、0.829 和0.677,分蘖数和分蘖角度在E2(P=0.030)、E3(P=0.018)、E5(P=0.045)下具有显著负相关关系,而株高和分蘖角度相关性不显著(P>0.05),但其相关系数在5个环境中为负值。由此可知,株高同分蘖数及分蘖角度可能存在遗传相关。
2.2 QTL及基因×环境互作
通过分析3个性状在6个环境下QTL,在ICIM方法检测中共发现13 个位点LOD 值大于3,其中株高检测到4 个位点,除qPH1,其余3 个位点(qPH7、qPH11、qPH12) 均在多环境下检出,qPH11在海南两个环境中检出,qPH12在哈尔滨两个环境中检出,而qPH7 同时在两个地点共4 个环境中检出,该位点位于7 号染色体RM1362~RM1306 区间,贡献率分别为22.78%、16.79%、28.15%和28.86%,且加性效应来自于高值亲本东农422,该位点在4 个环境下分别增加株高8.64、3.60、6.35 和6.38 cm。分蘖数共有3 个位点检出,qTN7-1 和qTN7-2 分别在6 个和3 个环境中检出,qTN7-1 平均贡献率为25.08%。分蘖角度共检出6个位点,分别为qTA2、qTA7-1、qTA6、qTA7-2、qTA12 和qTA8,其 中qTA2、qTA7-2、qTA12 和qTA8 在多个环境中检出。由表2 可知,qPH7、qTN7-1、qTN7-2、qTA2和qTA8加性效应来自于东农422,可增加株高、分蘖数和分蘖角,qPH12、qTA7-2和qTA12加性效应来自于空育131,可减少株高和分蘖数。这13 个位点在染色体上的分布如图2所示。
通过对3个性状不同QTL位点作超几何概率函数计算,得出株高QTL 同分蘖数QTL 之间存在显著相关性(P=0.046),分蘖数QTL 同分蘖角度QTL相关性达显著水平(P=0.039),但在株高和分蘖角度QTL 之间并未检测到相关性(P=0.293)。由此可知,利用东农422和空育131为遗传背景后代群体在株高和分蘖角度遗传上可能不具备共性。
多环境联合分析表明,除qTN11、qTN7-2 和qTA6,其余8 个位点均有显著的加性×环境效应,其中qPH7、qPH12、qTN7-1、qTA8 和qTA12 同时具有加性和加性×环境效应,且区间RM1306-RM70加性×环境贡献率最大,为3.90%(见表3和图3)。说明这类位点表达和环境互作共同决定性状表现。
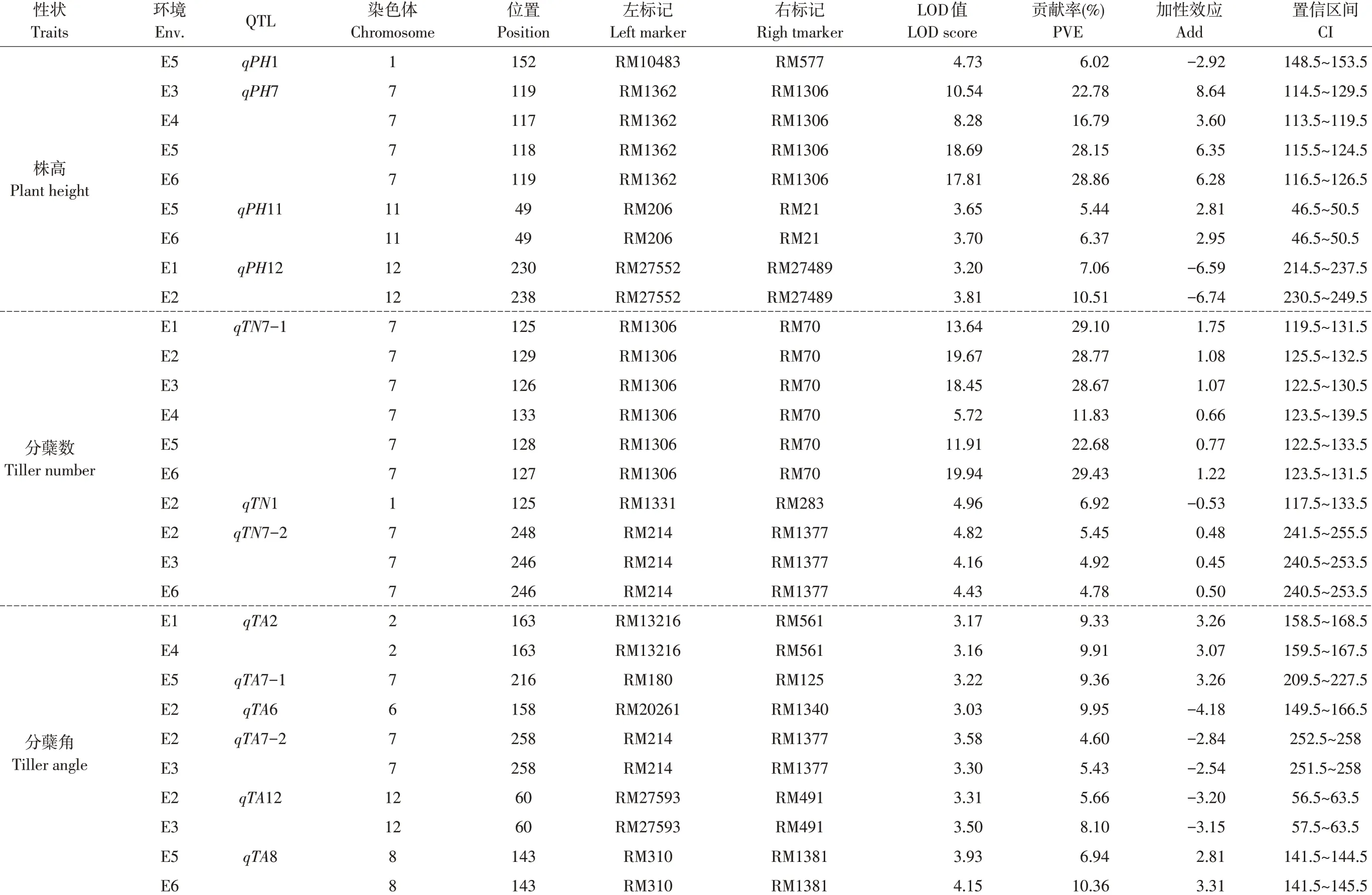
表2 基于完备区间作图法株高、分蘖数和分蘖角度在6个环境下QTL统计Table 2 QTL statistics of plant height,tiller number and tiller angle in six environments based on the ICIM
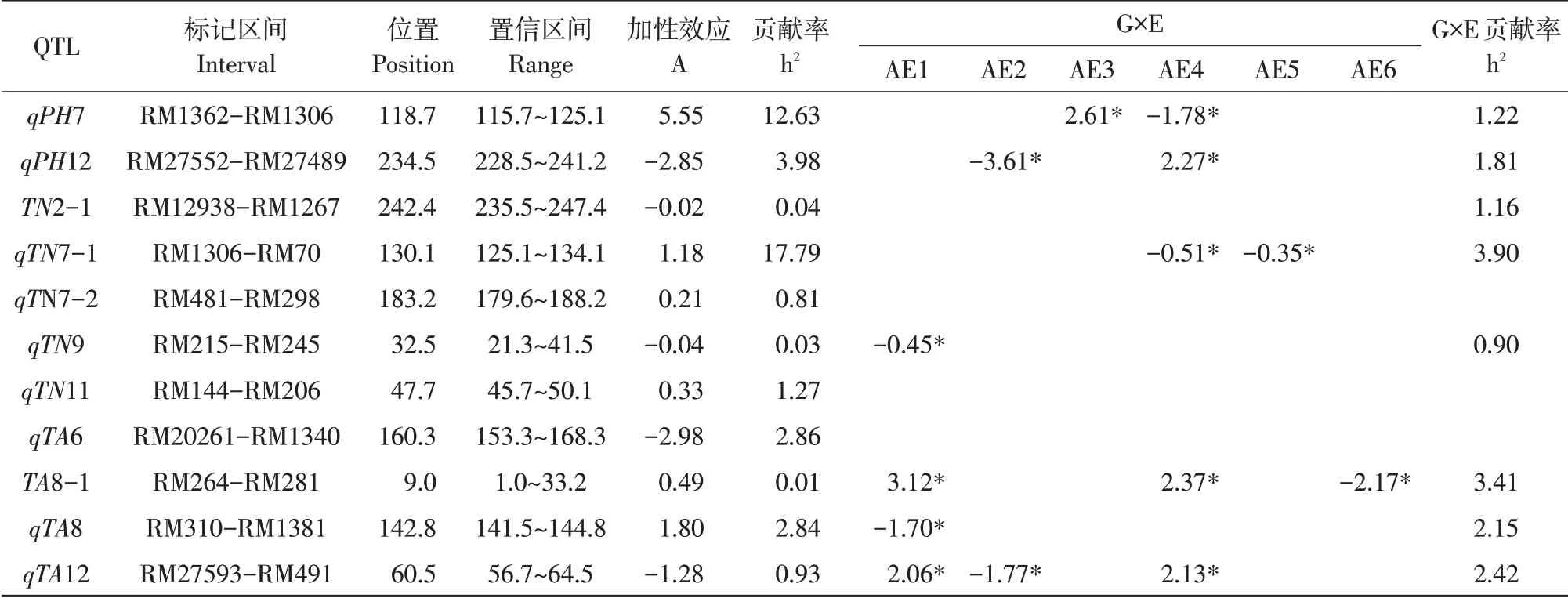
表3 株高、分蘖数和分蘖角度的QTL及QTL×环境互作分析Table 3 Analysis of QTL and QTL×Environmental interaction between plant height,tiller number and tillering angle
3 讨论与结论
株高、分蘖数和分蘖角度是水稻株型重要性状,本研究为发掘和水稻株型相关QTL 位点,连续三年两地环境下检测表型,从后代群体表型可知,3 个性状在海南表型均值高于哈尔滨(株高93.06>91.48, 分 蘖 数13.63>12.98, 分 蘖 角 度28.22>27.74),但同一性状在相同地点表型值差异不显著,且三性状广义遗传力均超过80%,说明表型变异主要受基因型主导。从相关性分析看,不同性状间相关性因环境而异,环境因素改变性状间相关性,但却影响其相关程度。在获得的QTL中,qPH7和qTN7-1贡献率超过20%,作为本研究发现的主效位点,这两个位点参与环境互作,但主要受基因型调控。
控制不同性状QTL 常位于相同区间,可能是数量性状位点一因多效或成簇分布的遗传现象。本研究发现的13个加性QTL位点中,RM1306标记分别同株高和分蘖数连锁,且该标记位点对株高平均贡献率为24.15%,对分蘖数平均贡献率为25.08%。因此,该标记可作为本研究发现的重要主效QTL,通过与物理图谱对比后,发现RM1362~RM1306 区间包含2 个注释基因(Os07g0668300 和Os07g0668200),基因产物分别为GDSL-脂肪酶/酰基水解酶和腺苷酸环化酶。与分蘖数连锁的区间RM1306-RM70 曾报道株高连锁位点,如Ph7a[13]、qPH7-2[14]、和OsBZR1[15]等,但这些位点均未发生遗传重合。由此可知,RM1306-RM70可能存在控制水稻株高的重要位点,但该区间内并未发现水稻分蘖数遗传相关位点,因此该位点可作为本研究中株高及分蘖产生共同连锁新区间。同时,这2个性状的遗传相关(P=0.046)和表型相关(P<0.01,见图1)具有一致性,验证研究可靠性。另外,本研究还检测到1个区间(RM214~RM1377)同时与分蘖数及分蘖角度有关,该区间通过物理位置比对,仅有300 bp距离(12 783 492~12 783 710 bp),该区间已报道8 个和水稻性状相关的位点,分别为gp7a、gw7、yd7a、tp7b[16]、qah7[17]、dth7.1[18]、qHd-7[19]和rn7a[20],其中仅tp7b 和分蘖数连锁,并未发现有分蘖角度位点存在,因此该位点可能与本研究检测到的qTN7-2 是同一位点。通过超几何概率函数计算发现,分蘖数和分蘖角度之间遗传相关(P=0.039)与性状相关(P<0.05)存在一致性。而剩余QTL 仅与一个性状连锁,其中,与qPH11 连锁的RM206 标记被证实是分布于粳稻资源中等位基因最多的标记,与qTN1 连锁的标记RM1331 则被报道和水稻叶色连锁[21-22],同样,qTA8 连锁标记RM310 和水稻叶边颜色连锁[23],而与qTA2 连锁的标记RM561 则与水稻耐冷性有关[24],其余位点并未检测到和水稻性状连锁的遗传位点,可能是研究发现的新位点。
水稻表型方差不但受基因型方差控制,还与环境方差及基因型×环境方差有关,基因和环境互作是水稻数量性状普遍存在的遗传模式[25-26],本研究通过连续三年在两个地点开展水稻株型相关形状遗传位点挖掘,分析环境因素对表型变异的影响。结果发现,同一性状在不同环境下可能存在不同环境效应影响表型方差,如本研究检测到qPH7,仅在两个环境下具有显著的基因×环境互作效应,其余4 个环境均未检测到,而qTN7-2、qTN11和qTA6则在任何环境下均未检测到基因×环境互作效应。同时发现,在2016 年哈尔滨和海南两个环境中检测到的基因×环境互作效应最多,而2017 年和2018 年4 个环境中检测到较少,可能与年际间环境差异有关。完备区间作图法检测到的13 个加性QTL 中,5 个在混合线性模型中得到验证,但在多环境中同时检测到的位点却并未受环境影响,如qTN7-1 在6 个单环境中均检测到,但仅E4和E5产生显著环境互作效应,说明其余4个环境下性状表型主要受基因型控制,而在E4和E5条件下,表型变异还受环境方差调控。由此可知,水稻数量性状遗传较复杂,基因表达不但受遗传背景控制,还与环境相关。因此仅在特定作图群体中获得的稳定遗传位点才可广泛应用于水稻性状遗传改良,而未经过多环境验证的位点则难以保证其真实可靠性。
本研究通过6个环境表型鉴定,在7号染色体相邻区间(RM1362~RM1306,RM1306~RM70)发现和水稻株高及分蘖数相关主效位点,而分蘖角度相关位点虽在多环境下检出(qTA2、qTA7-2、qTA12 和qTA8),但并无稳定位点和株高及分蘖数重合,因此将RM1306临近区间作为株型相关连锁区间,为今后基因定位和功能验证奠定理论基础。
猜你喜欢
中华医学图书情报杂志(2022年1期)2022-11-18
中国现代医生(2022年21期)2022-08-22
作物学报(2022年7期)2022-05-11
天津医科大学学报(2021年1期)2021-01-26
三农资讯半月报(2020年2期)2020-03-09
农村百事通(2019年17期)2019-10-08
现代农业科技(2017年1期)2017-03-06
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
养生保健指南(2016年12期)2017-01-06
