
种群水平生态风险评价方法概述及其在环境管理中的应用
2020-05-06 13:55廖伟刘娜冯承莲徐建王子健金小伟
生态毒理学报 2020年1期
廖伟,刘娜,冯承莲,徐建,王子健,金小伟
1. 南昌大学资源环境与化工学院,鄱阳湖环境与资源利用教育部重点实验室,南昌 330031 2. 中国环境科学研究院,环境基准与风险评估国家重点实验室,北京 100012 3. 中国科学院生态环境研究中心,北京 100085 4. 中国环境监测总站,北京 100012
生态风险评价是对有毒污染物生态危害量化的重要手段,是依据生态风险评价的原则和方法,以生态毒理学研究为基础,评价污染物进入环境后产生生态危害的可能性及程度。其目的是得出一个化学污染物浓度阈值(安全阈值)或风险值,为环境管理决策,特别是为相关环境标准或基准的制定提供参考依据[1]。环境暴露浓度和毒性阈值浓度是生态风险评价的2个重要组成部分,根据不同的毒性效应测试方法,生态风险评价方法包括3个层次:以单物种测试为基础的外推法,以多物种测试(微宇宙或中宇宙)为基础的生态风险评价,以种群或生态系统为基础的生态风险模型[2]。由于受研究基础和经费等因素限制,目前大部分化学污染物的毒性数据来自于个体或组织水平的单物种毒性测试[3-4]。然而,基于个体水平毒性测试所推导的安全阈值,忽略了物种通过竞争和食物链等作用产生的间接效应,也没有考虑时间和空间产生的影响,往往得到一些片面的结果,不能保证种群水平上的安全,难以满足对种群、群落及生态系统的评估,如Walton等[5]和Bechmann[6]分别发现污染物在对蚤状溞和分叉小猛水蚤个体死亡无影响的浓度下,导致种群数量下降。因此,从种群效应中得到污染物长期暴露下种群的动态生长、种群大小或生长繁殖等数据,能够保证生物群落中种群的持续性和生命力,具有更深远的生态学意义,比个体效应更适合用于污染物的生态风险评价[7-8]。
种群是指共同生活并繁殖的同一物种,如一群植物、动物或其他生物[9]。一般情况下,一个种群有明确的界限,如一个小型湖泊中生存的某种鱼是一个种群;另外,种群的界限也可以根据研究目的或研究方法划定,如栖息在一片叶子、一棵树、一排树或整片林地的某个物种[10]。基于个体水平生态风险评价的效应评估主要关注个体数量、死亡率、生殖状态、生长速率、新陈代谢、个体大小、年龄、个体生活史和个体环境暴露等特征及属性。而种群水平生态风险评价效应评估主要关注于生物量、种群数量、种群密度、平衡数量、自然增长率、性别比例、遗传多样性和可用的生境空间分布等特征[11]。因此,个体水平的效应评估主要由急性(如半数致死浓度LC50、半数效应浓度EC50)、慢性毒性数据(如无观察效应浓度NOEC等)计算获得,测试指标主要有致死、生长抑制、繁殖以及生化指标(如卵黄蛋白原)等[12-13]。而种群水平效应评估终点主要包括种群大小、恢复时间、性别比例、平衡数量、耐受性分布、基因分布和种群增长率等[11,14]。与个体水平相比,种群水平的生态风险评价包括了从组织、器官、个体、种群到群落和生态系统等高水平的模型,综合了个体毒性效应、竞争、迁移、栖息地以及其他生物和非生物因素的累积影响,在维持和保护动植物生存数量、保持生态系统稳定性等方面具有个体水平生态风险评价无法替代的效果。种群水平生态风险评价弥补了个体水平评价方法在时间和空间等方面的不足,因此,越来越多的研究者利用个体水平的毒性效应(如组织响应等)建立种群水平生态风险评价模型来预测种群响应,为研究种群水平生态风险评价效应评估指示及终点提供更多的选择。
因此,综述现有基于种群水平的生态风险评价方法不仅能促进该方法的研究和发展,还能直接用于环境管理决策,对控制进入水环境的污染物种类和数量、保护水体生物多样性及整个水生态系统的结构和功能具有重要意义。本文综述了文献中现有的种群水平生态风险评价关键科学问题、方法和应用,以期为进一步开展种群水平生态风险评价研究提供有益借鉴,同时也为环境风险管理中种群水平生态风险评价的应用提供理论依据。
1 种群水平生态风险评价研究的科学问题(Scientific issues in population-level ecological risk assessment)
1.1 评价终点(Assessment endpoint)
种群研究为风险评价提供了各种各样的终点,包括种群数量(population size)、平衡数量(equilibrium abundance)、稳定性(stability)、恢复时间(recovery time)、灭绝时间(time to extinction)、最小存续数量(minimum viable population)、耐受性分布(tolerance distribution)、大小及年龄结构(size and age structure)、性别比(sex ratio)、基因多样性(genetic diversity)和种群增长率(population growth rate)[14-16]。其中常用的评价终点是种群的个体数量,内禀增长率和灭绝时间等[16]。种群数量是野外调查和试验测试最直接的参数,常常被用于个体较小、难以鉴定年龄的生命周期较短的物种[17]。由于影响种群数量的因素较多而且波动较大,实际应用相对较少。种群增长率能够综合个体水平上很多指标之间的相互作用,比其他种群评价终点更敏感,因此常常被用作种群水平上的评价终点[8,18-20]。Forbes和Calow[18]将污染物对个体生活史(存活和繁殖)以及种群增长率的影响进行了比较,认为种群增长率与个体生活史参数相比能更好地评估污染物毒性效应。另外,种群灭绝时间是一个综合性更高的种群评价终点,常常用于评价有毒化学品对鸟类种群的风险[21-22]。种群水平的生态风险评价包括时间和空间上的种群信息,需要明确考虑生境及种群特性,根据期望的评价结果选择相关的评价终点十分重要。
1.2 密度依赖(Density dependence)
由于自然环境中空间、食物等资源有限,大部分自然种群不可能长期按指数增长,而是在一定范围内受到密度依赖反馈的调节,使种群维持在适度的范围内[23]。研究表明,在物种内部和物种之间,各种生物过程都可能产生密度依赖[24]。例如,随着密度的增加,种群的繁殖力逐渐降低,死亡率逐渐升高[25]。上述过程是种群可持续生存的基础,在保护生物学和害虫管理方面发挥了重要作用。当生态风险评价加入生物密度因素时,需要考虑2个重要问题:化学污染物对种群的毒性效应与密度依赖是相互独立的加和作用,还是随种群密度的变化而变化?如果二者之间是相互影响的,是拮抗作用还是协同作用?在拮抗作用下,种群密度较高时,化学品的毒性作用较弱,说明该种群的密度依赖程度较高,能够在受到污染物暴露损伤时,通过提高出生率得到补偿;在加和作用下,化学品的毒性效应不受种群密度影响;在协同作用下,种群密度越高,化学品的毒性效应越强,说明该种群的密度依赖程度较低,对多种胁迫因素敏感性较高(图1)[11,18,26-27]。一般来说,外部的死亡率因素往往与密度依赖关系表现出复杂的相互作用。与生物个体水平相比,种群水平的繁殖方式更复杂,化学污染物可能通过多种毒性效应对生物种群产生影响,在种群预测模型中加入其他生物学功能(如食物资源竞争),可以使预测结果更接近真实情况[28]。如果排除密度依赖,可能会削弱模型的预测能力,特别是对于长期毒性效应的预测[29]。因此,在种群水平风险评价中,获得种群密度依赖关系的可靠参数对种群水平生态风险预测具有十分重要的意义。
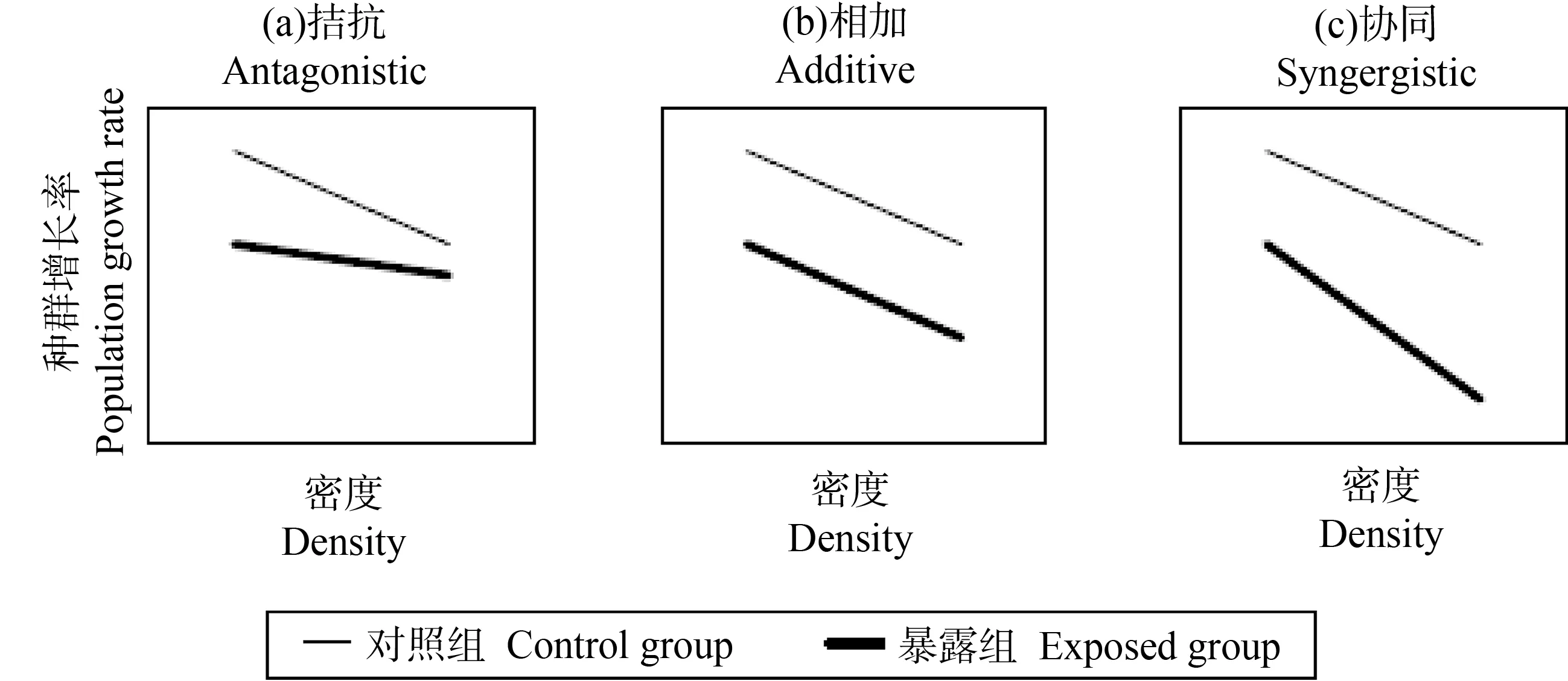
图1 密度与有毒化学品相互作用的主要类型[11]Fig. 1 Main types of interaction between density and a toxicant[11]
1.3 遗传变异(Genetic variation)
遗传变异是物种和种群适应当地条件并进化过程的基础[30]。为了适应环境的压力和变化(如杀虫剂的使用和化学品暴露),生物会产生足够的遗传变异。遗传变异可以用来描述当前和历史的种群特性,例如,种群大小和种群结构。然而,由于学科发展缓慢,与遗传相关的种群动力学研究较少,遗传变异很少被用于生态风险评价。随着越来越多的基因功能被发现,基因信息的获取越来越容易,遗传信息的改变得到了更好的解释[31-32]。在生态风险评价方面,这些研究进展有助于更好地理解环境压力、遗传变异和种群持续生存之间的关系。遗传变异包括中性变异、有害变异和适应性变异。中性遗传变异会通过随机遗传漂变而减少,通过突变和基因流动而增加。中性遗传标记的分析为确定种群结构、估计有效种群大小和迁移率提供了一种手段,并可以推断出群体之间的历史和地理关系。遗传学可以在种群状况、脆弱性和持续性问题上为种群水平的生态风险评价提供参考信息,并且有可能为经验方法和模型预测方法做出独特而有效的贡献,其中与适应性或毒性化学反应相关的特定功能基因变异对于种群水平的风险评价尤其重要。尽管将遗传学、统计学和毒理学相结合,研究毒性和耐受性的分子机制,可以更好地解释种群水平生态风险评价,但是目前还没有建立起相关的预测模型。因此,如何科学地将遗传变异应用于种群水平生态风险评价变得越来越重要。遗传信息可以通过考虑种群状况、脆弱性和持续性等因素来反映种群水平的生态风险评价。例如,遗传信息可以使种群结构和健康得到更好的表征,预测种群生存风险,解释长期胁迫因素(包括化学污染物)的影响。另外,遗传信息还能够用于阐明集合种群结构、与交互动力学有关的结构特征。从空间尺度上解释环境特征和压力源时,景观遗传学框架可以揭示多种环境和生物因素共同影响种群的作用规律。
生态风险评价引入遗传变异的关键问题在于遗传信息影响生态风险评价终点的机理研究和影响规律研究。化学适应的靶基因或潜在基因包括一些功能涉及毒理学或防御机制的基因,适应性化学变异常导致毒理学途径的下调或防御途径的上调[33]。虽然研究化学耐受遗传的生物化学或遗传机制仍然需要进一步深入,但目前随着这一领域学者的大量努力,昆虫对有机磷农药抗性的一些机制已得到验证,动物和植物对金属适应的几种生物化学机制也已经被报道,通常与螯合蛋白、金属硫蛋白上调等金属防御途径有关[34]。脊椎动物中,芳基碳氢化合物受体通路的基因及其在耐受剧毒二噁英类化合物中的潜在作用被很多人证实[35]。这些研究结果为化学污染物生态风险评价中引入遗传变异提供了基础。
1.4 空间结构(Spatial structure)
空间生态学认为,个体、种群及群落的空间位置与种群统计学、种间竞争和捕食者-猎物相互作用等具有同等的重要性,种群和群落的空间结构是构成生态学理论至关重要的组成成分[36]。因此,将更符合真实景观的空间结构和动物种群之间的关系引入生态风险评价,能够极大地降低生态风险评价的不确定性,为有效地制定保护对策提供科学依据。美国环境保护局(United States Environmental Protection Agency, US EPA)规定,为了在风险评价过程中充分反映时间和空间的影响,必须将化学品的毒性效应、迁移障碍和景观斑块结构的变化作为生态风险评价的附加因素[37]。
按照对空间处理方式的不同,空间结构分为3种不同的研究途径[36]:(1)集合种群(metapopulation)将空间看成由内部均质的生境岛屿构成的网络,物种不均匀地分布在斑块中。每个亚种群有不同的繁殖与死亡率,并且通过迁入和迁出过程关联在一起;(2)数学模型研究假定空间不仅是连续的而且是均一的,适用于连续空间内有局部相互作用和有限运动的生物个体;(3)景观生态学从更为复杂的空间格局出发,充分考虑了生境的空间异质性,生物体分布在一组相互作用、具有不同结构、功能和动态的异质性区域中。其中集合种群是研究物种空间分布的常用途径[38-39],McLaughlin和Landis[40]根据生物体在斑块之间迁移程度的强弱,将集合种群分为5种类型:孤立种群(isolated populations)、经典集合种群(classical metapopulations)、斑块种群(patchy populations)、大陆-岛屿集合种群和源-汇集合种群(mainland-island and source-sink metapopulations),以及连续性种群(continuous populations)。另外,随着3S(遥感、全球定位系统和地理信息系统)技术的发展,有关空间结构的数据和处理技术大大增加,为景观生态学在种群动态方面的研究提供了强有力的工具。
2 种群水平生态风险评价方法(Approaches to population-level ecological risk assessment)
种群水平生态风险评价要求考虑生物群体的综合特征,评价方法主要包括经验方法(实例研究)和模型研究。经验方法主要是通过野外实验、微宇宙或中宇宙实验收集表象生物特性的数据,并根据研究目的对数据进一步加工分析。经验方法主要用于杀虫剂登记(pesticide registration)、标准实验室测试研究(standard laboratory studies)、水生生物种群效应的经验测量(empirical measurement of population effects-aquatic species)、陆生生物种群效应的经验测量(empirical measurement of population effects-terrestrial species)、陆生野生动物种群栖息地污染场地调查(terrestrial wildlife population inhabiting contaminated sites)和栖息地评估(habitat evaluation)[11],其中在污染场地评估方面的应用尤为重要。经验方法中收集的生物特性数据一方面可以作为暴露指标直接用于评估压力源的潜在效应,另一方面是进行参数化、测试概念化或种群模型数学化用于风险评价回顾或展望。
随着数学和计算机等学科的发展,越来越多的研究者关注种群模型预测方法的研究,虽然模型预测方法受到资源、时间或其他因素限制,无法对种群风险进行准确的预测,但是通过经验数据进行验证或确认的生态模型所做的预测结果已经十分可靠。目前,在种群生物学中已经发展了多种用于种群水平生态风险评价的建模方法,数据和种群模型的结合被证明在预测和评估化学污染物环境风险上非常有用。按照不同的标准,种群动态模型有多种分类方法。例如,根据空间异质性分为非空间种群模型、半空间种群模型和空间显示模型[41];根据种群结构信息(如年龄、大小和性别等)分为非生物结构化种群模型和生物结构化种群模型。随着计算机技术的发展,越来越多的模型应用于种群水平生态风险评价,各个模型相互补充相互嵌套成为种群水平生态风险评价新的发展方向[11,15,42]。图2根据模型在构建和使用时所反映的空间尺度和生物组织层次,将种群模型分为非结构化种群模型、生物结构种群模型、个体模型、集合种群模型和空间显式模型[11]。
3 种群水平生态风险评价模型(Modeling approaches to population-level ecological risk assessment)
3.1 非结构化模型(Unstructured models)
非结构化模型是种群水平生态风险评价模型中最简单的形式,在模拟种群动态的过程中不考虑空间格局和种群结构信息(如年龄、大小和性别等)对种群动态的影响,假定个体在环境里均匀分布且不发生迁移,个体之间在生活史和环境响应方面没有差异,种群动力学只考虑出生率和死亡率。最早的非结构化种群模型是指数增长模型(式1)[43],该模型假设种群增长不受种群密度影响,适用于进入生态系统的新物种,或由于环境变化导致种群迅速增长。

图2 种群水平生态风险评价模型分类[11]Fig. 2 Taxonomy of population models for population-level ecological risk assessment[11]

(1)
式中:N为种群数量,t为时间,r为种群增长率。当r>0时,种群增长;r<0时,种群减少;r=0时,种群处于稳定状态。
大部分情况下,环境里的资源是有限的,当某一种群密度增大时,每个个体分配到的食物数量必然减少,从而导致种群的出生率降低,死亡率升高。Logistic增长模型利用种群承载力对指数增长模型进行了改进(式2)。
(2)
式中:K为种群承载能力。当种群数量较少时,种群生长速率接近于最大速率r,种群呈指数增长;随着种群数量增加,种群增长速率逐渐降低;当种群达到最大容量K时,增长速率为零。当种群受到反复干扰时,Logistic增长模型可以表达成离散的时间方程。
这类模型需要的数据量小,应用简单,但精确性不高,普遍用于筛选水平的风险评估。Adams等[44]2005年应用该方法评估了农田中反复喷洒农药对蚜虫的种群影响;Hendriks和Enserink[45]1996年利用实验室测试和野外调查的方法分别评价了多氯联苯暴露条件下大型溞(Daphnia magna)和鸬鹚(Phalacrocorax carbo)的种群数量变化;Nakamaru等[21,46]利用实验室和野外数据将非结构化模型应用于滴滴涕(DDT)对银鸥(Larus argentatus)和雀鹰(Accipiter nisus)的种群影响研究;Barnthouse[47]使用Logistic种群增长模型比较了农药暴露条件下不同类型水生生物的恢复率。
3.2 生物结构化模型(Biologically structured models)
由于个体基因和环境等因素的影响,种群的个体之间存在多种结构差异,如年龄、发育阶段、体长、性别和遗传信息等,这些差异既影响个体生命活动参数(如存活率、繁殖率、增长率、新陈代谢和能量消耗),也决定着群体行为[11]。生物结构种群模型考虑了种群的生物结构,表达了不同生命阶段对压力源的不同响应,能够更准确地反映物种的特征,比非结构化种群模型具有更高的真实性和精确性[23],同时也需要更多的数据和较高的费用[11]。目前应用最广泛的生物结构种群模型是Euler-Lotka方程和预测矩阵及其变体。
3.2.1 Euler-Lotka方程
在研究污染物对种群增长率的影响时,Euler-Lotka方程是相对简单的方法。Euler-Lotka方程由Lotka等[48]1925年提出,该方法将年龄作为影响生物繁殖力和死亡的唯一参数,利用生命表参数对繁殖力进行定量分析,只需通过实验室测试或野外调查获取种群的年龄结构,不考虑种群密度对种群增长率的影响[23]。其最基本的形式可用数学方程表示为(式3):
(3)
式中:x为生物的年龄,n为种群中生物的最大年龄,r为内禀增长率,lx为生物从出生到x岁的存活率;mx为x龄生物繁殖后代的平均数。由有毒化学品暴露导致的r值的变化作为慢性毒性指标,用作种群效应的评价指标,可以通过实验测定的存活量和繁殖量计算[49],也可以采用生物量动态模型估算[50]。
由于水蚤等桡足类动物为两性生殖,一般通过计算产幼雌体的比率、每个雌体产下的无节幼虫数量和后代雌体所占的比率的乘积得出繁殖量,因此对亲代种群增长率的研究需要持续2个完整的世代。Breitholtz等[51]在研究麝香类污染物对美丽猛水蚤(Nitocra spinipes)的种群毒性效应时,对Euler-Lotka方程进行了改进,用本世代的个体存活率、性别比和产幼雌体比率的乘积代替每个雌体的产雌率,忽略了污染物对美丽猛水蚤不同世代影响的差别,将实验周期缩短到24 d;黄瑛[52]在研究三丁基氧化锡对火腿许水蚤(Schmackeria poplesia)的繁殖毒性效应时,考虑了不同世代的差异,又对Breitholtz的方法进行了修正。
3.2.2 预测矩阵
年龄结构或发育阶段结构预测矩阵是对种群生长率潜力模型(Euler-Lotka方程)的重要扩展[25]。最简单的矩阵模型是线性的“Leslie矩阵”[53],它包含的信息与种群潜在增长率模型相同,但信息以几何形式或矩阵形式表示。种群丰度随时间变化的常见表达形式为矩阵方程(式4):
N(t)=LN(t-1)
(4)
式中:N(t)和N(t-1)是向量,包含了每一年龄组生物体的数量(N 0,…, Nk);L是矩阵,定义为(式5):
(5)
式中:Sk为生物从一个时间段到另一个时间段的存活概率;fk为生物在年龄k时的平均生育率。
这种结构一般是通过将种群特征或生命率分配给种群中不同类别的个体来反应,假定同一个类别的个体具有相同的生命率。该方法的困难之处在于如何给不同类别种群分类,以及如何充分反映物种生活史的复杂性。已有研究对生物生长阶段的确定提供了参考的方法[54-55]。Kuhn等[56-57]假定化学物质及其他环境胁迫因子与生物过程之间不存在相加或协同作用,用年龄结构投影矩阵模型和28-d毒性测试分别评估了20种化学品对糠虾(Americamysis bahia)的毒性效应,证明当前美国的水质标准在种群水平上对糠虾具有保护作用。此外,大部分模拟种群行为超过一代的模型都会引入密度依赖。Barnthouse等[58]应用随机、密度依赖的种群模型测试生活史、捕捞量、死亡率、环境变化及化合物毒性,预测暴露于化合物的鱼类种群反应的不确定性。
3.3 个体模型(Individual-based models, IBM)
种群的健康是个体健康程度的集体表现,由于个体之间对环境行为和生理反应不同,受到随机事件影响的可能性也不同,例如,在相似的胁迫条件下,每个个体的繁殖能力或死亡的概率具有一定的差异。个体模型是根据生物个体特征,建立设计所有水平细节的生物个体模型,通过方程解析解法,或对成百上千个个体的活动进行数值模拟,来推测种群总体水平的性质。IBM模型涵盖了大量特定的公式用于模拟个体年龄、大小和性别等,通常考虑多个生物体停留和交互的特殊情况或环境,要求从生物个体级别信息推测种群总体响应[59]。
建立个体模型的方法主要有2个,一个强调蒙特卡洛模拟,另一个强调方程的解析解法。其中解析解法的结果更具有普遍性,容易验证和理解[25]。个体解析模型一般强调生理特征,如代谢、生长和化合物毒性代谢动力学。早期IBM方法常用于模拟溞类生长-繁殖模型[60-61]、鱼类种群管理[62-63]。Kooijman和Metz[60]首次将其运用于毒物代谢动力学,通过研究有毒化合物对大型溞(Daphnia magna)个体生长和繁殖的影响,预测种群增长率。随着计算机和统计学的发展,研究者根据自己的需求倾向于多个模型组合。Hallam等[64]将该方法进行拓展,增加了以热动力学为基础的水中污染物摄取模型,并依据溶解的污染物浓度定义死亡率;Martin等[65]建立了DEB-IBM模型(dynamic energy budget- individual-based models)详细地研究水蚤种群变化的机理机制;Muller等[66]基于IBM模型结合生理结构种群模型(physiologically structured population models, PSPMs),得到种群动态的积分方程。
3.4 空间模型(Spatial models)
生物聚集程度区分了非结构化模型、生物结构化模型和个体模型(图2),这些模型均假定在均质空间条件下进行预测,没有涉及复杂空间结构[11]。随着计算机和地理信息系统等技术的发展,研究者对生物随机迁移、食物链和污染物暴露的空间异质性有更多的关注和预测方法[25]。应用较为成熟的两类空间模型为集合种群模型(metapopulation models)和空间显示模型(spatially explicit models, SEMs)。
3.4.1 集合种群模型
集合种群是由空间上彼此隔离,而在功能上又相互联系的2个或2个以上的亚种群组成的种群斑块系统[67]。集合种群模型通过反映种群亚单位的空间布局和环境,增强了模型的生态真实性,有利于阐明空间变化的化学暴露对栖息于空间异质化环境中种群的影响,包括斑块的大小、距离和密度,种群在斑块间的迁移速率和迁移形式,以及种群在斑块内的灭绝速率[11]。常见的集合种群模型有Levins模型(Levins models)、晶格模型(lattice models)、关联函数模型(the incidence function model)和源-汇动态模型(source-sink dynamics models),其中Levins模型是最早的集合种群模型,建立了给定时间内某一物种占据栖息地百分比、占据生活区后的消失速率及繁殖速率之间的关系[68-69](式6):
dP/dt = cP(1-P)-eP
(6)
式中:P为某一物种生活的斑块占总栖息地的百分比,c为种群在生活区的繁殖速率,e为种群在生活区的消失速率,t为连续时间。
该模型假设环境中的大量离散斑块之间通过物种迁移互相联系,忽略了斑块之间的差异性,如大小、栖息适宜性和斑块间距。平衡状态下,物种所在斑块占的比例为(式7):
P= 1-(e/c)
(7)
因此,只要种群在斑块内的繁殖速率大于灭绝速率(e/c<1),亚种群就会一直存在。
Levins模型考虑的基本过程与变量都是保护生物学中的基本问题,然而对于真实的种群管理来说过于简单而无法应用。Spromberg等[70]在式(7)的基础上,建立了一般性的集合种群模型,研究了某一个斑块被持久性有毒污染物和可降解有机物污染后,生活区的空间排列与连通对集合种群的影响。Chaumot等[71-72]扩展了Leslie矩阵法,用集合种群模型研究了河道网络中镉排放(建立镉排放位置与浓度的函数)对具有一定空间分布的褐鳟(Salmo trutta)种群的影响,计算不同暴露条件下褐鳟的种群生长率、年龄结构与空间分布,发现尽管排放到不同河流层次中的镉对种群增长率具有相同的毒性效应,仍然会影响褐鳟种群的空间分布。
3.4.2 空间显式模型
空间显示模型需要研究者将理论与个体生物觅食行为和繁殖结合在一起,将其与特定可测量的栖息地特征关联,推测栖息地变化对种群的影响,强调生态真实性和预测的准确性。生物体分布在真实的区域中,这些区域由不同类型的栖息生活区构成,且适合于研究物种生存。与集合种群模型相比,SEMs能更好地描述实际种群动态。由于考虑了景观生境类型、数量、质量和排列等特征以及环境压力源的空间分布,模型汇总大量的参数需要经验数据来验证[73]。随着计算机和地理信息系统等技术发展,空间显式模型得到了较大的发展和应用。在有害废弃物站点管理方面,Linkov等[74]研究了比目鱼暴露在疏浚材料弃置地点的空间显示建模,研究将暴露模型与生物结构或基于个体的模型、种群动态公式相结合,以描述危险废物场址污染物空间分布不均的情况下的原地沉积物对比目鱼种群水平影响。
3.5 动态能量预算(Dynamic energy budget, DEB)
动态能量预算为应用较多的生物能量学模型,其理论基础为在生物全生命过程中,能量流动和物质循环保持动态平衡,生物从外界获取的物质和能量通过同化作用贮存在体内,并用于自身的维持、生长和繁殖。Kooijman等[75-77]发展了基于k规则的动态能力核算模型(k-rule DEB model),提出能量比例k分配给维持和生长,剩余能量(1-k)用于发育和繁殖。为了实现生命的优化,生物通过改变自身维持、生长和繁殖过程的能量配比来应对外界环境胁迫[65,78]。DEB不仅将生物体的生理特征与种群生长率和年龄分布联系在一起,同时,也将暴露浓度和有毒化学品作用模型与个体生理特征联系在一起[79]。因此,DEB可用于个体模型参数推导,也可以作为一个嵌入模型,用于预测其他模型涉及的特征参数,如Rose和Cowan[63]以代谢、生长、觅食行为及猎物选择等参数建立了鱼类生命阶段和年龄的方程,其中关于鱼类个体的生物能学、繁殖及觅食行为等详细模型均需要通过DEB相关模型方法来进行模拟校正。DEB模型主要应用于栖息在相似的环境中、生活周期相对简单的水生生物,在实验室和野外实验均得到了应用。Klanjscek等[80]将DEB模型应用于细菌种群研究,Chizinski等[69]报道了模式生物斑马鱼和黑头软口鲦的生物能量学模型。目前利用能量分配理论进行种群外推的实例研究主要集中在溞类[68]、糖虾[81]、摇蚊[82]和鱼盘螺[73]等生命周期较短、易繁殖的水生无脊椎动物。
4 种群水平风险评价在环境风险管理中的应用(Application of population-level ecological risk assessment in environmental risk management)
环境管理的最终目的是保持生态系统功能的完整性、稳定性和持久性。种群水平风险评价方法为各国环境管理制定相关法律法规、提升生态系统关联性、以及为管理制度提供必要信息用于支持实用的、和谐的和易实施的管理决定提供了很多有力的参考依据,广泛用于新产品生态风险评价、控制性危险废物的管理、不受控危险废物的豁免、生物监测、资源管理战略合作、资源分配风险分析比较、非化学压力源管理和资源管理保护等。早在20世纪40年代,发达国家就已经开始进行种群水平的风险评价研究(表1)。美国环保局分别在1993年[83]和1998年[37]的风险评价指南中将种群风险评价列为生态风险评价的方法之一,2003年将以保护食鱼性水鸟种群为目标的水质标准的修订列入了研究计划。日本在最近几年也展开了种群水平风险评价的研究[84]。
美国环保局认为种群水平的生态风险评价可以提供比特殊化学品风险熵更直观的风险管理决策,国家1号超级基金将生态风险降低到可以恢复或维护本地生物种群和群落健康的水平作为环境管理的参考依据。1993年美国环保局使用投影矩阵种群模型评估了受《有毒物质控制法》控制的氯化石腊对虹鳟鱼种群大小的影响[83];1994年美国环保局模拟评估了一项水坝和水库建设项目的生态风险,认为该项目可能导致具有特殊价值的鸟类种群灭绝,因此根据《清洁水法》第404条否决了该项目的建设许可证[85]。加拿大的风险管理一般也侧重于保护生物种群、群落和生态系统[86]。加拿大不列颠哥伦比亚省的水、土壤和空气保护部预计,在污染场地项目管辖范围内,1%的污染场地需要进行三级生态风险评估,该评估是一个详细的定量评估(主要是考虑种群水平生态风险评估),涉及多种不相关的化学品胁迫、多种栖息地和地形,以及广泛的地理区域。欧盟在风险评价技术指南[87]中将野生动物种群水平风险评估纳入不同级别生态风险评估框架,特别提出杀虫剂的环境风险评估重点是保护大部分非靶生物,这在大多数情况下涉及到种群水平的影响。日本为了防止对生态系统的不利影响,已修订或制定了相关国家法律、政策、条例或指示。日本在种群层面进行风险评估的第一个案例是化学物质管理,提出了壬基酚、共面多氯联苯和三丁基锡风险管理在种群水平的层面决策,建议种群增长率作为种群水平生态风险评价终点[84]。


5 展望(Expectation)
从种群角度来看,生物个体的死亡或损伤是没有意义的,因为多数生物个体只有很短的生命周期,仅很少部分能达到最大寿命,具有完整的生命周期。人们普遍认为种群的干扰与不稳定是正常的,而实验室中所观察到的稳态情况都不符合实际情况。正如自然界许多生物种群都因遭受频繁、极端的环境变异(如火山爆发、森林火灾和大规模环境污染)而大量死亡。因此,单纯的毒性测试无法预测种群水平的反应,利用种群数学模型将生物个体水平实验数据与种群反映水平相关联,是今后的重点研究方向。环境风险管理的本质在于增强个体数量的保护,但除了濒临灭绝的个体,没有必要保护每个个体的数量。风险管理者应着重从技术层面考虑种群水平风险评价方法,并在管理层面上以种群水平风险评价研究结果作为风险管理的重要参考依据。
种群生态风险评价主要以种群模型为基础,而模型的建立需要经过收集大量种群水平的野外数据、选择合适的模型以及估计参数等一系列过程,这一过程复杂且不确定性因素较多,限制了其在生态风险评价中的应用。种群水平生态风险评价需要进一步在技术层面上开展以下研究:
(1)研究种群水平生态风险评价评价终点对种群风险效应评估的指示效果,筛选出高稳定、易测量的评价终点;
(2)根据实例研究和模型预测的需要,加大物种特征参数的预测,将密度依赖、遗传变异和空间结构等种群水平生态风险评价中可考虑的关键科学问题加入到评价方法中,增加生态风险评价的真实性和可靠性;
(3)充分考虑模型构建的不确定性和实际应用的验证。
种群水平生态风险评价建立统一规范的指导文件,不仅能进一步促进种群水平生态风险评价方法研究和实例应用,还将为风险管理决策中考虑种群水平的风险提供有力的支持。将种群水平生态风险评价服务于管理决策层面,需要进一步确定评价方法的适用性、风险值的可接受性,丰富相关特定物种的种群参数的基础数据,提高风险值的关联,以保证和维持特定动植物的足够生存数量。此外,增强淡水生态系统中种群水平生态风险评价的应用,增加种群水平风险评价的专业培训和指导是管理层面上关注的重点。随着越来越多的风险管理者对种群水平风险评价方法和结果的认可,环境管理决策中体现出种群水平生态风险评价作为参考依据的文件越来越被人们所接受。因此,需要一个有效的转换方法或指示,将这些效应意义转化为生态风险评价,从个体生存、生长和繁殖,引申到种群的影响,用个体水平的效应去预测种群响应。
猜你喜欢
今日农业(2022年15期)2022-09-20
天天爱科学(2022年9期)2022-09-15
天天爱科学(2022年4期)2022-05-23
当代水产(2022年3期)2022-04-26
湖南电力(2021年1期)2021-04-13
航空世界(2020年10期)2020-01-19
劳动保护(2019年7期)2019-08-27
福建基础教育研究(2019年11期)2019-05-28
红土地(2018年7期)2018-09-26
中学科技(2015年1期)2015-04-28
