
植被恢复对库布齐沙漠非生长季土壤CH4、N2O通量的影响
2020-04-20 13:30:18段玉玺王伟峰李晓晶刘宗奇
生态学报 2020年6期
王 博,段玉玺,王伟峰,李晓晶,刘宗奇,刘 源
内蒙古自治区林业科学研究院, 呼和浩特 010010
CH4和N2O是大气中重要的温室气体,具有很强的吸收红外辐射并保存红外热能的能力,其增温潜势分别为CO2的28倍和265倍[1],对温室效应的贡献率在所有温室气体中分占第2和4位[2]。自工业革命以来,随着人类活动和土地利用变化,大气中CH4和N2O浓度年均增长分别为0.8%和0.3%,是造成全球气候变暖的主要原因之一[3]。因此,对陆地生态系统CH4和N2O排放规律和机制开展研究,是降低温室效应和减缓气候变暖的重要基础。
目前,全球荒漠化面积超过3600万km2,占地球陆地面积的1/4,荒漠生态系统已成为地球圈层气体型循环的重要组成部分。近年来,国内外学者对荒漠生态系统温室气体通量的研究逐步开展,如Castillo-Monroy等[4]研究发现荒漠生态系统中生物结皮是土壤CO2排放的重要来源;Mosier等[5]研究了耕作和施肥等方式对干旱区土壤CH4和氮氧化物通量的影响;Zhang等[6]认为沙漠化逆转可以改变土壤CO2的排放速率;Zhao等[7]研究了降雨对温带荒漠土壤温室气体排放的影响。国内学者如赵哈林等[8]对科尔沁沙地土壤呼吸特征进行了研究;徐冰鑫等[9]利用模拟增温试验分析了全球气候变暖背景下荒漠土壤温室气体的排放特征;滕嘉玲等[10]认为沙埋可显著影响生物结皮覆盖下土壤的温室气体通量。相关研究也表明,人工建植促进植被恢复是荒漠生态修复和实现沙漠化逆转的重要手段。在这一过程中,土壤-植被系统、环境水热因子等已被证实能够产生显著变化,而有关植被恢复对荒漠土壤CH4和N2O通量的影响鲜有报道,胡宜刚等[11]认为随着荒漠生物结皮的发育和演替,土壤对CH4和N2O的吸收量逐渐减弱,但影响机制尚不明确。
在高纬度和高海拔地区,非生长季土壤冻融循环和冰封是普遍存在的现象。土壤反复结冻和融冻,能够直接影响土壤水热平衡、有机质的分解和微生物动态,进而间接改变土壤温室气体的排放规律[12]。目前,对非生长季土壤温室气体通量的研究主要集中于高纬度苔原、草地和北方森林[13- 14],但荒漠土壤CH4和N2O通量在结冻-融冻循环过程中的变化特征尚不明确。因此,开展对干旱荒漠区非生长季土壤CH4和N2O通量及其影响机制的研究非常必要,可以丰富和完善陆地生态系统碳氮循环的理论体系。
本研究以库布齐沙漠东部植被恢复程度不同的5种沙地类型为研究对象,采用静态暗箱-气相色谱法,测定非生长季荒漠土壤CH4、N2O通量的动态变化,探讨植被恢复过程中环境因子对通量的影响,阐明荒漠土壤非生长季温室气体的“源/汇”属性,为科学评价人工干扰措施对荒漠生态系统碳氮循环的影响提供理论依据。
1 研究区概况
研究区位于内蒙古自治区鄂尔多斯市准格尔旗境内的布尔陶亥治沙站(110°48′29.28″E,40°03′—42.13″N),地处库布齐沙漠东部,包含流动沙地、半固定沙地和固定沙地等沙漠地貌类型。该区为温带大陆性气候,季节变化明显,干旱多风,昼夜温差大。水热同期,年均气温6.1—7.1℃,年均降水量240—360 mm,年均蒸发量2560 mm,年均日照时数3138 h,无霜期130—140 d,年均风速3.3 m/s。试验区海拔1100—1300 m,土壤以风沙土为主,植被主要包括沙柳(Salixcheilophila)、柠条(Caraganakorshinskii)、油蒿(Artemisiaordosica)、猪毛菜(Salsolacollina)、沙竹(Psammochloavillosa)、沙米(Agriophyllumsquarrosum)、蒙古蒿(Artemisiamongolica)等。
2 研究方法
2.1 样地设置
根据植被恢复程度及沙漠化逆转阶段将样地划分为以下类型:(1)流动沙地,植被盖度极小的裸露沙丘,生长少量一年生草本植物,地表无结皮发育;(2)半固定沙地,当先锋灌木油蒿在沙地定植后,随时间推移,群落逐渐稳定,风蚀逐渐减弱,地表有少量物理结皮分布;(3)藻结皮固定沙地,2012年在部分半固定沙地呈网格状扦插沙柳提高植被盖度,因冠层荫蔽地表逐渐形成发育初期的藻结皮(叶绿素a含量为0.31 μg/g,伪枝藻素含量为0.28 μg/g,类胡萝卜素含量为0.16 μg/g);(4)地衣结皮固定沙地,2002年行带式扦插造林后形成以沙柳为优势种的固定沙地,土壤及微气象条件改变促进地衣结皮发育(叶绿素a含量为0.95 μg/g,伪枝藻素含量为1.72 μg/g,类胡萝卜素含量为0.31 μg/g);(5)苔藓结皮固定沙地,研究区内植被恢复的最优阶段,植被盖度最大,基本无风蚀,有较厚的凋落物层,地表结皮已发育为演替后期的苔藓结皮(叶绿素a含量为1.93 μg/g,伪枝藻素含量为7.62 μg/g,类胡萝卜素含量为0.58 μg/g)。各样地基本情况见表1。
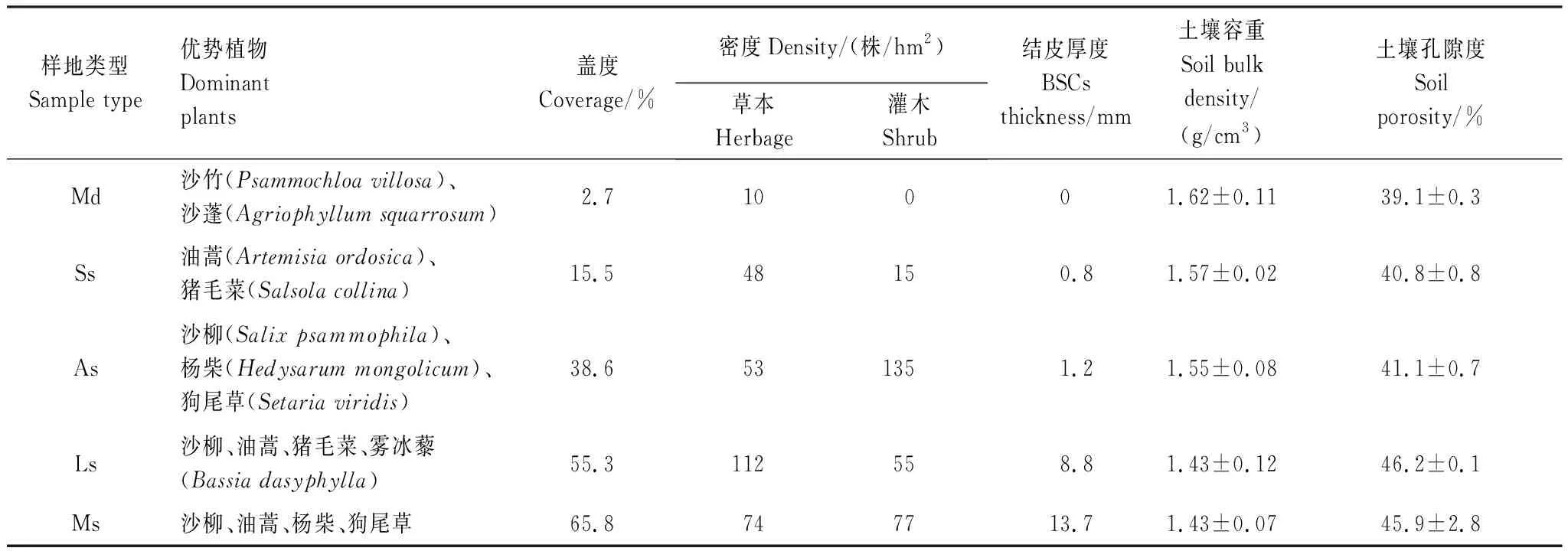
表1 样地基本信息
Md:流动沙地,Mobile dune;Ss:半固定沙地,Semifixed sand;As:藻结皮固定沙地,Algal crust fixed sand;Ls:地衣结皮固定沙地,Lichen crust fixed sand;Ms:苔藓结皮固定沙地,Moss crust fixed sand
2.2 气体通量测定
使用静态暗箱-气相色谱法进行土壤CH4和N2O的采集和通量测定。静态暗箱由规格为320 mm×600 mm的圆筒形顶箱和基座组成,顶壁安装有搅拌风扇,使箱内气体混合均匀,基座预先嵌入样点土体进行稳定,采样时将顶箱扣入基座凹槽并向内注水以密封。气体样品采集在2017年11月—2018年4月的非生长季进行。根据研究区土壤冻结状态将非生长季分为3个时期,2017年11月13日—12月22日为土壤结冻阶段,2017年12月23日—2018年3月2日为土壤冰封阶段、2018年3月3日—4月23日为土壤融冻阶段。在5类样地内,分别选择生境相对一致的3个平坦地块作为采样点并进行基座嵌入。土壤结冻和融冻期每10 d采样一次,冰封期每月采样1次,非生长季共采样11次。每次采样时间为9:00—12:00,罩箱时间为30 min,即在罩箱0、15、30 min各取气体样品3次,每次采集气体3袋。采用100 mL医用带三通阀的注射器收集箱内气体,存储于铝箔气袋中带回实验室,采用气相色谱仪(Agilent 4890D,USA) 测定样品中CH4、N2O的浓度。
通过气体浓度随时间的变化,计算单位面积土壤气体交换量,正值表示土壤排放CH4和N2O,为“源”,负值表示土壤吸收CH4和N2O,为“汇”。气体通量计算公式如下:
式中:F为被测气体通量(μg m-2h-1);ρ为标准状态下气体密度(kg/m3);h为静态暗箱高度(m);dC/dt为箱内气体浓度变化斜率;T为采样时箱内平均温度(℃)。
2.3 环境因子测定
研究区气象资料由小型自动气象站记录(HOBO,USA)。气体采样时同步监测样地土壤及空气温湿度。土壤温湿度使用水分速测仪测定(TRIME PICO,Germany),按0—20、20—40、40—60、60—80、80—100 cm划分为5个土层。地表(地上5 cm)及空气(地上100 cm)温湿度使用手持气象跟踪仪测定(Kestrel 4500,USA)。
样地植物群落和土壤性状调查于2017年夏末进行。各样地类型随机布设3块10 m×10 m的灌木样地,每块灌木样地内随机布设5块1 m×1 m的草本和凋落物样方,统计样地内灌木生长参数并计算平均值,依此选择标准株3株,采用全挖法获取灌木地上、地下生物量。采用收获法获得草本地上、地下及凋落物生物量。每块样地内同样分5层挖取3个土壤剖面,各层混合取样带回实验室阴干、粉碎、过筛,采用重铬酸钾-浓硫酸氧化外加热法测定有机碳含量,采用凯式定氮法测定土壤全氮含量,采用环刀法测定土壤容重并计算孔隙度。
2.4 数据处理
试验数据处理及作图采用Excel 2007及SigmaPlot12.5软件进行,并采用SPSS 20.0软件进行统计检验,选取最小显著极差法(LSD)进行不同植被恢复阶段温室气体通量差异性检验(α=0.05);使用Pearson检验对气体通量与环境因子进行相关性分析。
3 结果与分析
3.1 非生长季荒漠土壤水热因子时空变化特征
如图1所示,5种类型荒漠土壤水热因子在非生长季变化基本一致,表现出鲜明的水热同期特征。自进入结冻期,土壤温度持续降低,在无降水补给的情况下,土壤含水量同样不断减小,此阶段5类样地土壤温度变化范围为-3.9—17.0℃,土壤含水量为1.8%—11.1%。冰封期内发生数次降雪事件,地表出现雪被,土壤温度和含水量处于稳定的低水平状态,此阶段5类样地土壤温度变化范围为-1.3—-10.5℃,土壤含水量为1.2%—4.8%。自进入融冻期,气温迅速回暖,土壤温度逐渐升高,积雪消融和冻层融化为土壤补充大量水分,导致土壤含水量显著增大,此阶段5类样地土壤温度变化范围为2.2—23.2℃,土壤含水量为3.8%—11.8%。
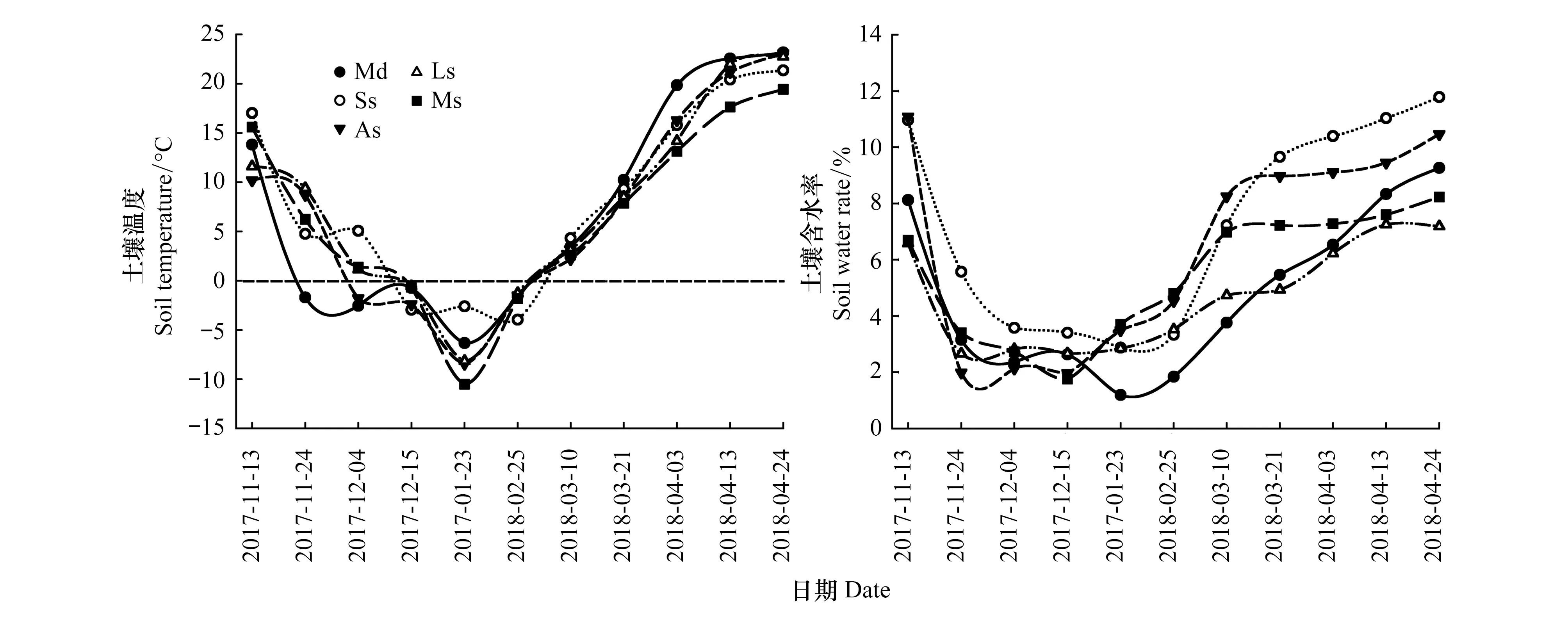
图1 非生长季0—100 cm平均土壤温度和含水量变化Fig.1 Changes in mean soil temperature and water rate between 0 to 100 cm depths during non-growing seasonT:土壤温度,Soil temperature;W:土壤含水量,Soil water rate
3.2 非生长季荒漠土壤CH4、N2O通量时空变化特征
由图2可知,5种类型荒漠土壤在非生长季均以吸收CH4为主,是CH4的汇。各样地CH4通量随时间的变化规律不同,流动沙地CH4吸收量最大值出现在12月4日(-42.0 μg m-2h-1),半固定沙地最大吸收量出现在4月24日(-46.4 μg m-2h-1),而在4月13日出现少量CH4排放(16.8 μg m-2h-1)。藻结皮固定沙地CH4最大吸收量出现在12月15日(-41.6 μg m-2h-1),其余时间吸收量无明显波动。地衣结皮固定沙地CH4通量变化呈单峰曲线,排放峰值出现在2月25日(6.1 μg m-2h-1),此前吸收量逐渐减小,土壤由汇转为源,此后吸收量逐渐增大,土壤由源转为汇。苔藓结皮固定沙地在试验期间均表现为吸收CH4,最大吸收量出现在4月13日(-112.2 μg m-2h-1)。
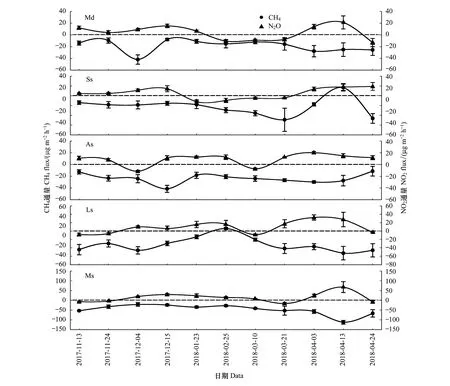
图2 荒漠土壤CH4、N2O通量时空变化Fig.2 Spatial-temporal variation of desert soil fluxes of CH4,N2O
荒漠土壤N2O通量随时间的波动较为剧烈,正负值变化无明显规律。总体来讲,5种类型荒漠土壤在非生长季对N2O的排放大于吸收,是N2O的源。流动沙地对N2O的最大排放量出现在4月中旬(21.4 μg m-2h-1),2月25日吸收量最大(-13.9 μg m-2h-1)。半固定沙地在冰封期及融冻初期均为吸收N2O,1月23日吸收量最大(-12.2 μg m-2h-1),其余时间均为排放,融冻末期排放最大(19.1 μg m-2h-1)。藻结皮固定沙地大部分时间为排放N2O,4月初排放最大 (20.2 μg m-2h-1),仅在12月初及3月初出现少量吸收。地衣结皮固定沙地分别在结冻初期、融冻初期和末期少量吸收N2O,其余时间均为排放,最大排放量出现在4月3日(32.5 μg m-2h-1)。苔藓结皮固定沙地N2O通量变化规律同地衣结皮相似,最大吸收量为3月21日(-16.5 μg m-2h-1),最大排放量为4月13日(67.6 μg m-2h-1)。
3.3 非生长季荒漠土壤CH4、N2O通量差异分析
如表2所示,5种类型荒漠土壤在非生长季CH4、N2O平均通量差异显著(P<0.05)。从流动沙地到苔藓结皮固定沙地,土壤对CH4的吸收通量逐渐递增,后者为前者的2.5倍,土壤对N2O的排放通量同样逐渐增加,后者为前者的3.6倍。非生长季荒漠土壤CH4累积吸收量和N2O累积排放量分别为-0.074—-0.182 g/m2和0.007—0.065 g/m2,各样地间均差异显著(P<0.05)。根据环境温度变化将非生长季分为结冻期、冰封期和融冻期,不同时期内荒漠土壤CH4、N2O平均通量差异同样显著(P<0.05)。
不同冻结时期荒漠土壤温室气体排放量对非生长季累积排放占比越大,吸收或排放贡献率越大。如表2所示,不同时期累积占比在样地间的分异较大。对CH4而言,流动沙地、地衣结皮和苔藓结皮固定沙地均在融冻期吸收贡献率最大,而半固定沙地和藻结皮固定沙地则在冰封期吸收贡献率最大。对N2O而言,流动和半固定沙地在结冻期排放贡献率最大,而藻结皮、地衣结皮和苔藓结皮固定沙地均为冰封期排放贡献率最大。就研究区而言,荒漠土壤在融冻期CH4吸收贡献率最大,在结冻期N2O排放贡献率最大。
3.4 土壤水热因子对CH4、N2O通量的影响
如表3所示,非生长季流动沙地与半固定沙地土壤CH4通量与土壤温度和含水量关系不显著(P>0.05),但随着沙地被固定,土壤水热因子开始不同程度影响CH4通量。藻结皮固定沙地CH4通量与深层土壤含水量(60—100 cm)呈显著负相关(P<0.05),地衣结皮固定沙地CH4通量与各土层温度及深层土壤含水量(40—100 cm)均呈显著负相关(P<0.05),苔藓结皮固定沙地CH4通量与0—100 cm土壤含水量及各土层温度均呈极显著负相关(P<0.01)。土壤水热因子对非生长季N2O通量的影响较小,除半固定沙地N2O通量与土壤温度存在显著正相关外(P<0.05),其他样地N2O通量与土壤水热因子关系均不显著(P>0.05)。
3.5 植被恢复对CH4、N2O通量的影响
在人工建植促进荒漠植被恢复过程中,植物生物量不断积累,从流动沙地到苔藓结皮固定沙地,根平均生物量由0.8 g/m2增至255.8 g/m2,地上生物量由2.1 g/m2增至568.2 g/m2,凋落物生物量由0增至69.9 g/m2,总生物量由2.9 g/m2增至893.7 g/m2;在土壤-植被协同作用下,荒漠土壤性状也逐渐改善,从流动沙地到苔藓结皮固定沙地,0—100 cm深度土壤平均有机碳含量由0.22 g/kg增至1.10 g/kg,全氮含量由0.02 g/kg增至0.07 g/kg,土壤容重由1.62 g/cm3降至1.43 g/cm3,土壤孔隙度由39.1%增至45.9%。
植物生物量和土壤性状不断积累和改善的过程能够显著影响荒漠土壤CH4和N2O通量的变化。如表4所示,荒漠土壤CH4通量与土壤有机碳含量和孔隙度呈显著负相关,与全氮含量呈极显著负相关,与土壤容重呈显著正相关,与地上及凋落物生物量呈显著负相关,与根及总生物量呈极显著负相关。荒漠土壤N2O通量与土壤有机碳和全氮含量呈显著正相关,与土壤容重呈显著负相关,与浅层土壤(0—40 cm)孔隙度呈显著正相关,与地上、根系及总生物量呈显著正相关,与凋落物生物量呈极显著正相关。
4 讨论
4.1 非生长季荒漠土壤CH4通量变化
CH4的产生和排放是一个复杂的生物化学过程,目前对非生长季土壤CH4“源/汇”的性质仍不明确。本研究表明,非生长季荒漠土壤总体表现为CH4的吸收汇 (-39.0 μg m-2h-1),这与胡宜刚等[11]对腾格里沙漠的研究结果一致。这是因为荒漠土壤属于好氧环境,在有氧条件下,土壤中的甲烷氧化菌可氧化CH4并将其当作唯一的碳源和能源。土壤中存在两类甲烷氧化菌:一类生长于高浓度CH4(>40 mL/m3) 环境中,属低CH4亲和力;第二类生长于接近大气CH4浓度的环境中,属高CH4亲和力[15]。不饱和的荒漠土壤以第二类甲烷氧化菌为主,其氧化CH4的临界浓度通常小于大气CH4浓度,因而这些甲烷氧化菌的生存依赖于对大气CH4的消耗[16]。一些学者对其他类型生态系统如青藏高原高寒草甸[17]、高纬度落叶松林[13]、温带刺槐林[18]以及农牧交错区草地土壤[19]的研究也认为非生长季土壤以吸收CH4为主,但Groffman等[20]研究发现非生长季的硬阔林土壤会出现少量CH4排放;吕金岭等[21]认为冻融期绿洲灰漠土CH4排放比重占全年排放量的6%—14%;孙向阳等[22]发现冬季北京低山区森林土壤CH4的吸收量几乎为0;陈哲等[23]发现在非生长季水稻田以排放CH4为主,而玉米田则以吸收CH4为主。这种生态系统间的差异可能与土壤CH4通量是产甲烷菌及甲烷氧化菌共同作用和传输的综合过程有关,不同气候带中植物群落和土壤类型有较大分异,导致两种菌群类别、数量和活性产生变化,从而改变了非生长季CH4通量格局。
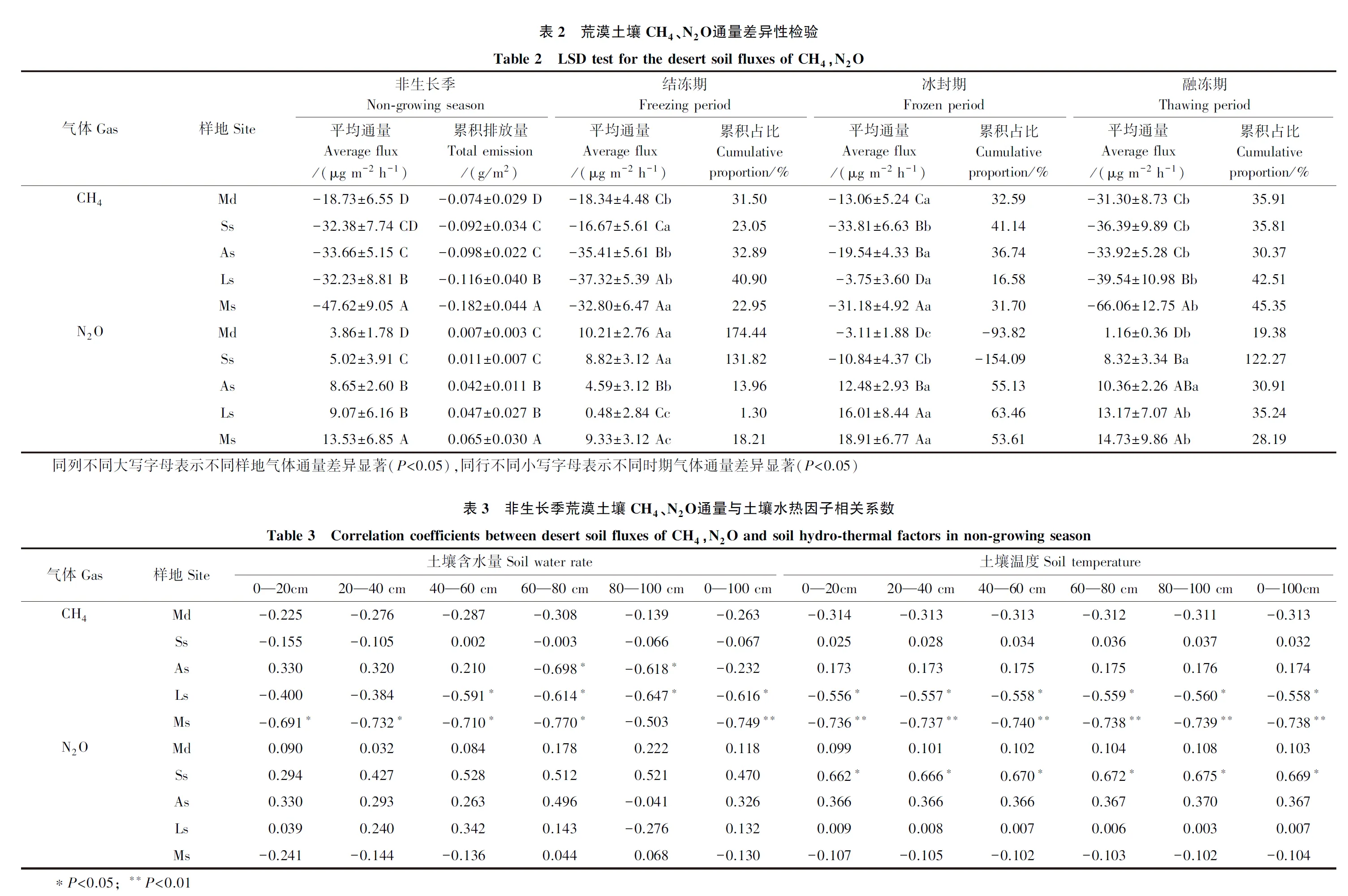
表4 植被恢复过程中荒漠土壤CH4、N2O通量与土壤-植被因子相关系数
Table 4 Correlation coefficients between desert soil fluxes of CH4,N2O and soil-vegetation factors in the process of vegetation reconstruction

气体Gas土壤因子Soil factors土层Soil layer/cm0—2020—40 40—60 60—80 80—100 0—100 植物因子Plant factorsCH4有机碳-0.975**-0.753-0.832-0.786-0.824-0.941*地上生物量 -0.949*全氮 -0.985**-0.903*-0.930*-0.999**-0.994**-0.987**根生物量 -0.993**容重 0.916*0.907*0.901*0.8020.900*0.930*凋落物生物量-0.933*孔隙度-0.909*-0.923*-0.851-0.812-0.846-0.912*总生物量 -0.964**N2O有机碳0.923*0.7970.951*0.8380.959**0.932*地上生物量0.948*全氮 0.905*0.897*0.997**0.929*0.907*0.934*根生物量 0.951*容重 -0.904*-0.871-0.906*-0.595-0.774-0.884*凋落物生物量0.967**孔隙度0.924*0.904*0.8000.6530.6470.835总生物量 0.955*
*P<0.05;**P<0.01
土壤水分是影响CH4吸收速率的一个重要因素,但在不同的土地利用方式或土壤含水量范围内,CH4吸收速率与土壤含水量的关系存在很大差异。在本试验中,附着生物结皮的固定沙地土壤含水量与CH4通量呈显著负相关性,这与Striegl等[24]研究结果一致。森林土壤CH4吸收率通常与土壤含水量呈负相关性,这是因为甲烷氧化菌消耗CH4的能力主要取决于土壤中气体扩散的速率,森林土壤含水量较高导致气体扩散受阻,甲烷氧化菌活性下降,CH4吸收率会随土壤含水量的增加而降低[25]。而荒漠土壤含水量较低且具有良好的气体交换通道,利于氧化菌对CH4的氧化,但过低的水分含量也会影响微生物活性,干旱胁迫明显限制了土壤对CH4的吸收。当沙地逐渐被固定,植被对沙质土壤能够起到一定的水源涵养作用,土壤含水量保持在一定范围后,甲烷氧化菌重新恢复活性,CH4吸收率随土壤含水量的上升而增大[26]。
土壤温度也是影响CH4吸收速率的另一个重要因素。试验结果显示,在温度较低的非生长季,地衣结皮和苔藓结皮固定沙地CH4通量与土壤温度呈显著负相关性,这是因为土壤温度通过改变产甲烷菌和甲烷氧化菌群落结构而间接影响CH4通量。产甲烷菌对外界温度变化十分敏感,其最适生存温度为30—40℃,在非生长季温度降低会显著削弱产生CH4的能力[27]。而甲烷氧化菌对温度的敏感性弱于产甲烷菌,Sitaula等[28]发现在平均温度低于1℃的高纬度森林土壤中仍有大量甲烷氧化菌存在,而且荒漠土壤中的甲烷氧化菌为高CH4亲和力菌群,氧化CH4时所需活化能较低,所以低温时仍能大量吸收CH4。李俊等[16]发现土壤温度在-5—10℃时对甲烷氧化菌的影响较为明显,而在10—20℃时则无法左右其活性。本研究中非生长季荒漠土壤平均温度为4.7℃,因此甲烷氧化菌正处于受土壤温度显著影响的范围。
不同植被覆盖与土地利用方式变化对土壤CH4吸收速率有较大影响。本研究中,通过人工建植促进荒漠植被恢复,使流动沙地逐渐演变为固定沙地,在此过程中,CH4通量发生显著变化。从流动沙地到苔藓结皮固定沙地,非生长季CH4平均吸收量由18.7增至47.6 μg m-2h-1。植被恢复对CH4通量的影响主要表现为三个方面:首先,甲烷氧化菌在团粒结构较好的壤土中可保护自己免受干扰,而在保水保肥性差的沙土中则易失去其高活力的生态位,导致CH4吸收率下降[29]。植被恢复是加速沙地土壤演变和细粒化的过程,固定沙地土壤容重减小,孔隙度和黏粒含量的增加可为甲烷氧化菌提供良好的生存环境,促进CH4吸收。其次,植被恢复是土壤养分富集的过程,植被-土壤系统的协同和反馈作用能够显著增加荒漠土壤底物浓度,活性有机碳是土壤微生物生长的速效基质,可有效促进微生物种群扩大和活性提高[30]。最后,植物发达的通气组织对土壤-大气之间的气体交换起着重要的传导作用,根系气孔氧气的下行渗漏促进CH4在根际土壤发生氧化,同时根系在大量吸收养分的同时可将甲烷氧化菌等土壤微生物从有害底物中释放,提高其氧化CH4的能力[31]。因此,在荒漠生态系统中,植被恢复可显著增加土壤对CH4的吸收能力。
4.2 非生长季荒漠土壤N2O通量变化
非生长季不同生态系统中土壤N2O的“源/汇”属性同样存在高度的不确定性。本研究中荒漠土壤为N2O的排放源 (7.9 μg m-2h-1),而胡宜刚[11]等认为腾格里沙漠在非生长季为N2O的弱汇 (-0.9 μg m-2h-1);其他生态系统如高寒草甸[17]和硬阔林[13]在非生长季排放N2O,北方农牧交错区草地冻融季N2O排放可占全年排放的70%[19],而温带半干旱草原则表现为吸收N2O[32],东北典型农田生态系统在覆雪期会出现N2O弱吸收[23]。土壤N2O是在土壤微生物的参与下,通过硝化作用(自养硝化和异养硝化)、反硝化作用(生物反硝化和化学反硝化)、硝态氮异化还原等作用下产生的[33],不同生态系统中硝化或反硝化菌群的变化可能是导致“源/汇”分异的重要原因。
本研究表明,虽然土壤水热因子不是影响N2O通量的主控因素,但N2O仍表现出较强的季节性变化。很多研究认为冬季土壤N2O的排放量增大,本研究也证明了以上结论。这种冷季高排放现象出现的原因可能表现为以下四个方面:第一,非生长季冻融交替对参与N2O产生的硝化和反硝化细菌数量影响较小,土壤反硝化酶在低温状态下仍能保持较高活性,所以在非生长季土壤依然可以大量排放N2O[34];第二,非生长季地表自然覆盖物主要为凋落物和积雪,较厚的覆盖层能够隔离表层土壤与冷空气的直接接触,降低土壤冻结强度及冻结深度,形成温暖和厌氧的土壤环境有利于反硝化过程中N2O的产生[20];第三,冻融循环可以改变土壤养分格局,破坏团聚体稳定性,使部分细根或微生物细胞分解,增强土壤碳、氮矿化能力,有效氮含量升高会提升反硝化菌群活性,促进反硝化作用产生N2O[35];第四,禁锢-释放理论。有研究表明,当地表被积雪覆盖或土壤冻结时,土壤-大气之间的气体交换十分微弱,反硝化作用产生的N2O将被暂时封存于土壤孔隙中,当温度升高土壤开始融冻后,被禁锢的N2O将呈脉冲式排放[36]。Teepe和Ludwig[37]也认为冻融循环后沙质土壤中N2O气体排放没有增加,而壤土中N2O排放量明显增加。由此可知,不同质地土壤在冻融循环作用下具有不同的N2O排放特征,禁锢-释放机理并不适用于荒漠生态系统。
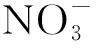
5 结论
非生长季荒漠土壤是CH4的吸收汇,是N2O的排放源。在植被恢复促进沙漠化逆转过程中,CH4和N2O通量变化明显,从流动沙地到苔藓结皮固定沙地,荒漠土壤对N2O的排放和CH4的吸收量逐渐增大。根据环境温度变化将非生长季分为结冻期、冰封期和融冻期,不同时期荒漠土壤CH4、N2O通量差异显著,研究区土壤在融冻期CH4吸收贡献率最大,在结冻期N2O排放贡献率最大。环境因子对CH4、N2O通量的影响有所不同,土壤含水量能够显著影响3类固定沙地土壤对CH4的吸收,土壤温度是地衣结皮和苔藓结皮固定沙地CH4通量的重要影响因子。除半固定沙地土壤温度与N2O通量存在显著正相关关系外,其他样地中土壤水热因子不是影响N2O通量的主控因素。植物生物量的积累和土壤理化性质的改善能够显著影响荒漠土壤CH4、N2O通量,表明人工建植促进植被恢复会显著改变荒漠生态系统的温室气体排放格局。
猜你喜欢
新疆大学学报(自然科学版)(中英文)(2022年3期)2022-06-04 13:45:40
军事文摘(2021年18期)2021-12-02 01:27:58
草业学报(2021年6期)2021-06-28 11:01:36
内蒙古林业(2021年6期)2021-06-26 10:42:32
幼儿智力世界(2021年4期)2021-05-17 17:10:29
水土保持通报(2020年6期)2020-03-10 01:46:52
科普童话·百科探秘(2018年7期)2018-09-13 11:12:08
新疆大学学报(自然科学版)(中英文)(2018年4期)2018-02-10 00:09:15
辽宁林业科技(2017年4期)2017-06-22 10:23:36
浙江大学学报(工学版)(2016年2期)2016-06-05 09:20:50
