
基于亲水相互作用液相色谱-三重四极杆质谱法研究白茶萎凋过程中代谢物的变化
2020-04-20 08:43:46陈翔田月月张丽霞
茶叶科学 2020年2期
陈翔,田月月,张丽霞
基于亲水相互作用液相色谱-三重四极杆质谱法研究白茶萎凋过程中代谢物的变化
陈翔,田月月,张丽霞*
山东农业大学园艺科学与工程学院,泰安市茶叶工程技术研究中心,山东 泰安 271018
采用亲水相互作用液相色谱-三重四极杆质谱法,对白茶鲜叶(F)和室内萎凋处理29 h(W)、53 h(P)的2个萎凋样进行了代谢物靶向检测,Quality control(QC)样本中共检测到111个代谢物,主要包括47种氨基酸及其衍生物、29种核酸类代谢物、12种维生素和辅酶、17种糖代谢物和6种甘油磷脂代谢物。与鲜叶F相比,萎凋叶W和P中显著增加的代谢物分别有39种和41种,其中共有代谢物35种;显著降低的代谢物分别有9种和11种,其中共有代谢物8种。显著变化代谢物分析结果表明,萎凋叶的细胞膜受损、DNA和RNA降解、氨基酸衍生物显著增加、糖代谢异常。根据代谢物含量的变化幅度和组成特异性,筛选了表征萎凋叶细胞膜生理状态和核酸、糖代谢的标志代谢物。此外,针对代谢物中氨基酸和核苷酸的组成特点,提出了研究氨基酸衍生物滋味特性的必要性和应用代谢谱分析建立萎凋工艺适度标准的可能性。
白茶;萎凋;靶向代谢谱
白茶是我国六大茶类之一,成茶具有“外形自然,满披白毫,毫香凸显,香气清鲜,滋味鲜醇,汤色杏黄”的品质特征[1]。传统白茶加工由萎凋、干燥工序组成,而萎凋是白茶品质形成的关键环节。
萎凋是指在一定环境条件下,鲜叶散失部分水分同时发生一系列以水解为主体的化学反应过程。萎凋是白茶、红茶和乌龙茶加工工序之一,其中以白茶的萎凋时间最长、萎凋失水减重最大。已有研究表明,白茶经萎凋,干物质含量下降4.2%~4.5%,多酚类物质下降36%,儿茶素总量下降49%,总糖下降50%以上[2],游离氨基酸含量显著增加[3]。由于受分析技术的制约,人们尚未对上述物质变化的相关代谢途径(糖、氨基酸、核酸等)及细胞膜结构特性变化进行研究,而目前兴起的代谢组学为开展这方面的研究提供了有效的手段和方法。
代谢组学(Metabolomics或Metabonomics)指应用核磁共振技术或色谱-质谱联用等分析技术,研究生物体或组织甚至单个细胞中核苷酸、氨基酸、糖类、脂类等全部小分子代谢物(分子量小于1 000)的成分及其动态变化的学科[4]。代谢组学根据所检测的代谢物范围分为靶向代谢组学(Targeted metabolomics)和非靶向代谢组学(Untargeted metabolomics)两类,对复杂生物样本广泛应用的是基于色谱-质谱联用技术的靶向代谢组学分析。目前,代谢组学在茶树生长发育[5]、茶叶品种分类与鉴别[6-9]、茶叶加工工艺优化[10-12]、茶叶品质控制[13-14]、茶叶原产地和年份鉴别[15]等领域加以应用[16]。Dai等[17]基于UHPLC-Q-TOF MS的非靶向代谢组学方法分析白茶萎凋过程中代谢物动态变化表明,氨基酸类物质酪氨酸、缬氨酸、亮氨酸、苯丙氨酸、赖氨酸、脯氨酸和异亮氨酸以及TF 3-没食子酸酯、TF没食子酸酯、TF二聚儿茶素、茶双没食子儿茶素A和茶双没食子儿茶素B的含量至少增加了5倍,而儿茶素、没食子儿茶素和没食子儿茶素没食子酸酯的含量降低了50%以上。
本文采用亲水相互作用液相色谱-三重四极杆质谱法,对白茶鲜叶(F)和萎凋处理29 h(W)、53 h(P)的2个萎凋叶进行氨基酸、核苷酸、维生素和辅酶、糖类和甘油磷脂类小分子代谢物进行靶向代谢谱分析,旨在了解白茶萎凋过程中代谢物的变化规律及其对白茶品质的影响,并为白茶萎凋工艺技术参数的确定提供分子水平的理论依据。
1 材料与方法
1.1 试验材料与仪器
材料:2018年6月中旬在泰安桃花峪茶园采摘福鼎大白茶树一芽二叶的新梢,茶树树龄为3年。进行室内厚摊萎凋处理,其室内环境条件为:温度23~33℃、平均温度28.3℃;空气湿度35%~64%、平均空气湿度48.6%。分别选取鲜叶和2个萎凋叶(处理29 h和53 h)各25 g于微波炉中,高火4 min,80℃烘至水分约为5%,粉碎机粉碎。准确称取0.1 g茶粉于离心管中,加入10 mL 100℃沸水浸提5 min,每隔1 min对离心管进行上下摇晃,然后置于高速离心机离心10 min,转速为8 000 r·min-1,4℃下取2 mL上清液经0.22 μm滤膜过滤,分别标记为F、W和P,–80℃保存,每个样品重复提取3次,均进行代谢组学分析。
仪器与试剂:格兰仕微波炉、FW100高速万能粉碎机(天津市泰斯特仪器有限公司);DHG-9140A型电热鼓风干燥箱(上海精宏实验设备有限公司);AB5500 QqQ质谱仪(美国AB SCIEX公司);Waters I-class超高效液相色谱仪(美国Waters公司);5430R低温高速离心机(德国艾本德股份公司);Fluka氨水(美国Sigma-aldrich公司);色谱柱(ACQUITY UPLC BEH Amide l.7 μm,2.1 mm×100 mm,美国Waters公司);乙腈(德国Merck公司)。
1.2 试验方法
1.2.1 样品制备方法
取试验样品在4℃缓慢解冻后,每份取等量100 μL,加入400 μL预冷甲醇-乙腈(甲醇∶乙腈=1∶1),涡旋混合,–20℃孵育1 h沉淀蛋白质,14 000,4℃条件下离心20 min,取上清液,真空干燥。质谱检测时加入100 μL乙腈-水溶液(乙腈∶水=1∶1)复溶,14 000,4℃离心15 min,取上清液进样分析。
1.2.2 色谱条件
样品采用Waters I-class超高效液相色谱仪进行分离。流动相:A为水溶液+25 mmol·L-1乙酸铵+25 mmol·L-1氨水(pH=9.75),B为乙腈。样品置于4℃自动进样器中,柱温40℃,流速为0.3 mL·min-1,进样量1 μL。液相梯度如下:0~1 min,B维持在95%;1~14 min,B从95%线性变化到65%;14~16 min,B从65%线性变化到40%;16~18 min,B维持在40%;18~18.1 min,B从40%线性变化到95%;18.1~23 min,B维持在95%。队列中每隔6个试验样本设置1个QC样本,共3个QC样本。
1.2.3 MRM质谱条件
采用AB5500 QqQ质谱仪进行质谱分析。ESI源条件如下:鞘气温度,350℃;干气温度,350℃;鞘气流量11 L·min-1;干气流量10 L·min-1;毛细管电压,正负模式中分别为4 000 V或–3 500 V;喷嘴电压,500 V;喷雾器压力,30 psi。采用MRM模式监测,每个MRM离子对的停留时间为3 ms,总的循环时间为1.263 s。
1.3 数据处理
采用MRM analyzer(R)对200个代谢物的MRM原始数据进行提取,得到各代谢物的峰面积,数据采集以及数据分析过程参照Cai等[18]研究结果。单变量分析方法包括变异倍数分析(Fold change analysis)、检验。
2 结果与分析
2.1 试验质量控制
采用QC样本对试验过程中系统的稳定性和重复性进行评价,试验结果见表1。结果表明,QC样本中检测到111个代谢物的相对标准偏差RSD<30%,说明这些物质在整个试验过程中稳定性和重复性较好。
2.2 靶向代谢物种类
将所检测到的111个代谢物依据代谢途径和分子结构特点,分为氨基酸及其衍生物、核酸代谢物、维生素和辅酶、糖代谢物和甘油磷脂代谢物5类。
2.2.1 氨基酸及其衍生物
共计47种,按分子结构特点及来源可分为蛋白质氨基酸、蛋白质氨基酸衍生物、非蛋白质氨基酸和寡肽4组(相对含量显著增加的共有代谢物用“*”标注,相对含量显著减少的共有代谢用“#”标注)。
(1)蛋白质氨基酸(14种):-酪氨酸Tyr*、-天冬酰胺Asn*、-亮氨酸Leu*、-缬氨酸Val*、L-丝氨酸Ser、-色氨酸Trp、-谷氨酰胺Gln、-精氨酸Arg、-脯氨酸Pro、-赖氨酸Lys、-苯丙氨酸Phe、-天冬氨酸Asp、-谷氨酸Glu#、-蛋氨酸Met#。
(2)蛋白质氨基酸衍生物(25种):①乙酰化(6种):-乙酰基--酪氨酸*、乙酰基--亮氨酸*、-乙酰基--苯丙氨酸*、-乙酰基-谷氨酰胺、6-乙酰基--赖氨酸、-乙酰基--丙氨酸;②甲基化(2种):二甲基甘氨酸*、肌氨酸(-甲基甘氨酸)*;③氨基酰化(1种):-氨基酰--天冬氨酸;④羟基化(1种):-4-羟基--脯氨酸;⑤脱氨衍生物(5种):-GABA*、苯乳酸*、对羟基苯甲酸*、咪唑丙烯酸、4-胍基丁酸#、⑥脱羧衍生物(3种):-乙酰尸胺*、色胺、-乙酰腐胺;⑦其他途径衍生物(7种):-2-氨基己二酸*、甲基胍*、-蛋氨酸亚砜;5-氨基戊酸、-瓜氨酸、-犬尿氨酸、5-氨基酮戊酸。
(3)非蛋白质氨基酸(4种):-腺苷高半胱氨酸*、-高丝氨酸、-高半胱氨酸、-哌啶酸(又称-高脯氨酸)。
(4)寡肽(4种):--谷氨酰--苯基丙氨酸、肌肽(--丙氨酰--组氨酸)、甘氨酰--亮氨酸、氧化谷胱甘肽。
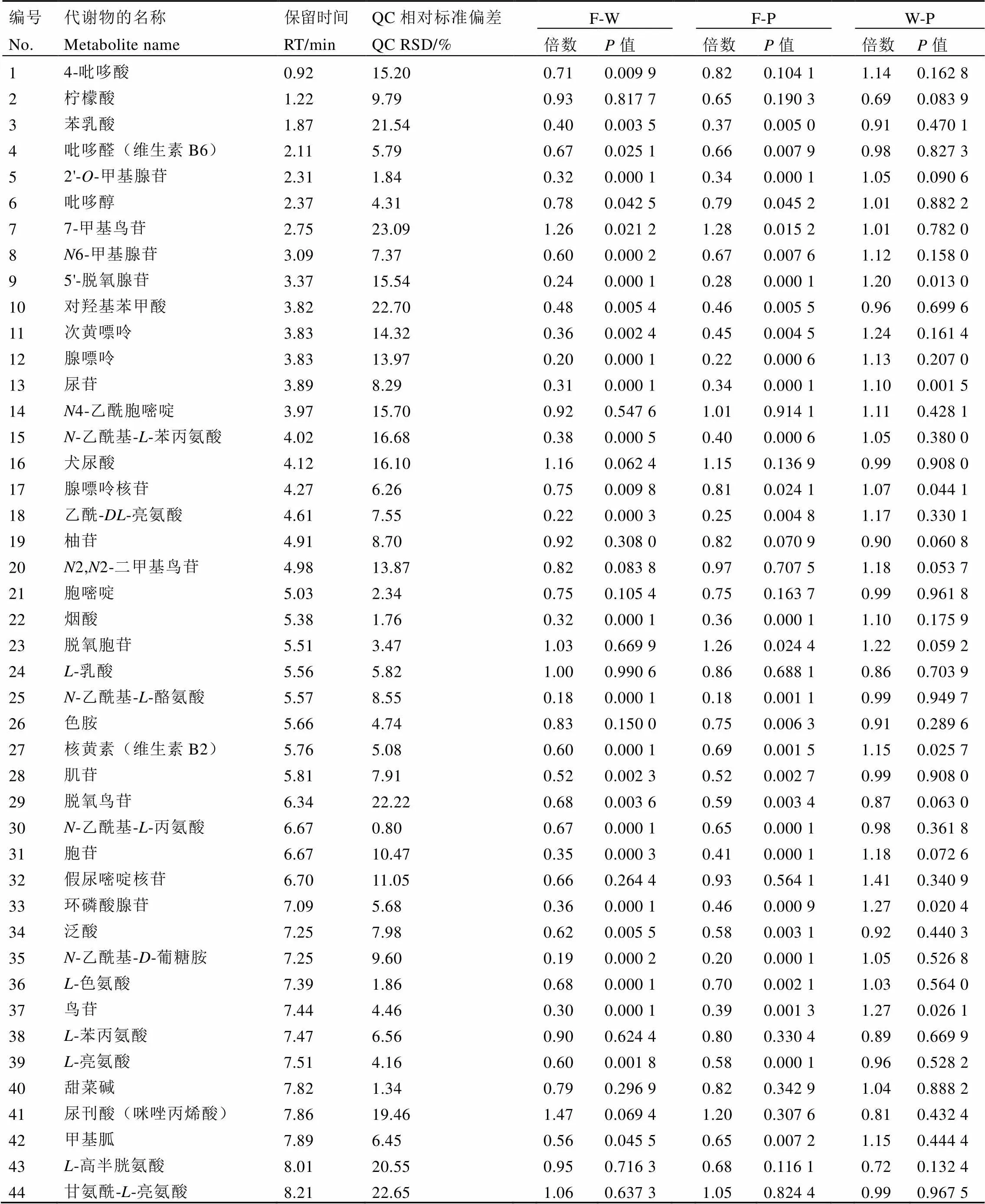
表1 检测出的代谢产物及其变化倍数
注:“倍数”是指鲜叶和萎凋叶中代谢物的含量比值。当倍数>1时,表明代谢物在萎凋叶中的含量降低,且值越大,下降幅度越大;当倍数<1时,表明代谢物在萎凋叶中含量升高,且值越小,则上升幅度越大
Note: ‘Fold’ refers to the ratio of metabolites in fresh leaves and withered leaves. When fold>1, it indicates that the content of metabolites in the withered leaves is decreased, and the larger the value, the greater the decrease. When fold<1, the content of metabolites in the withered leaves is increased, and the smaller the value, the greater the increase
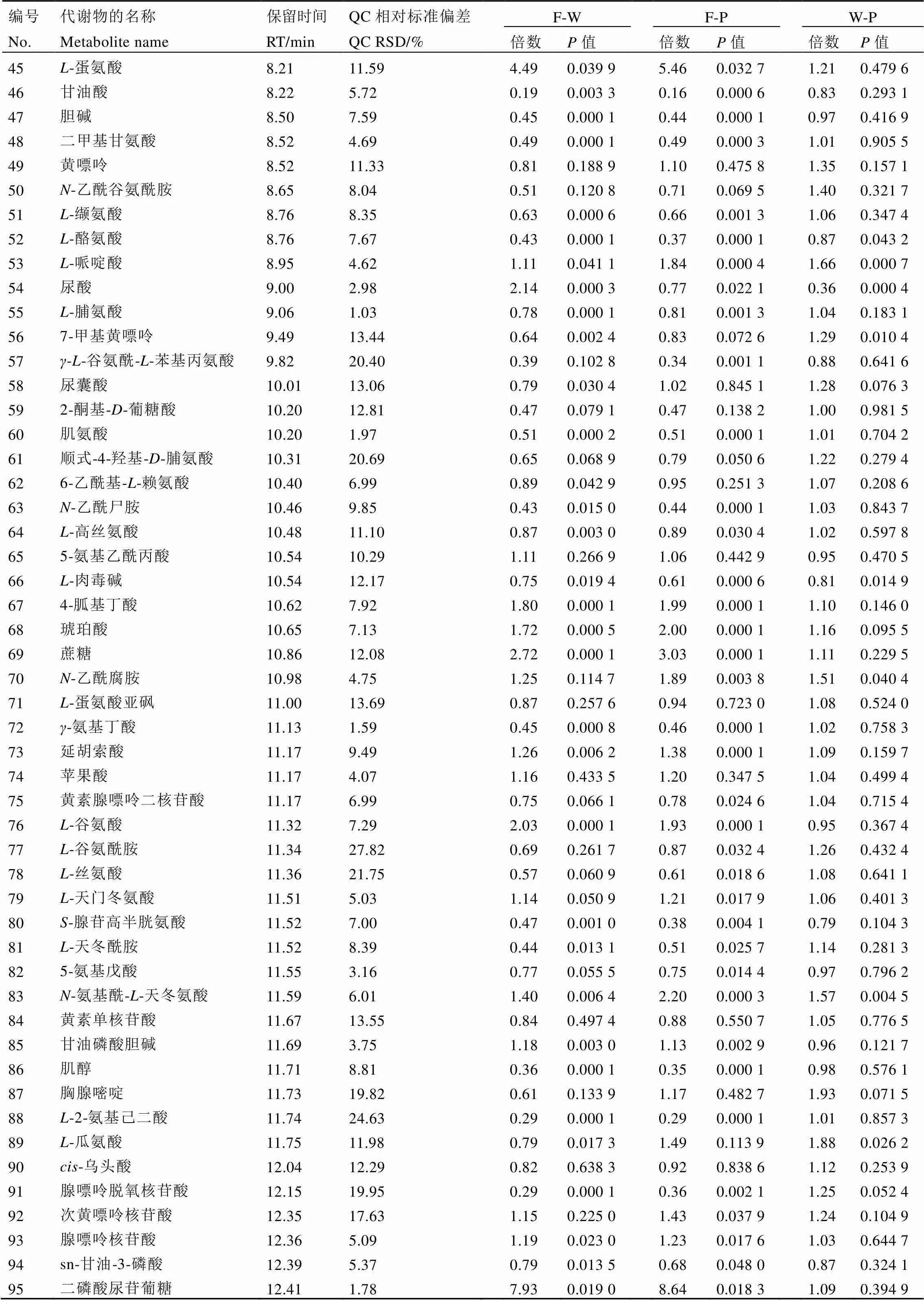
续表1
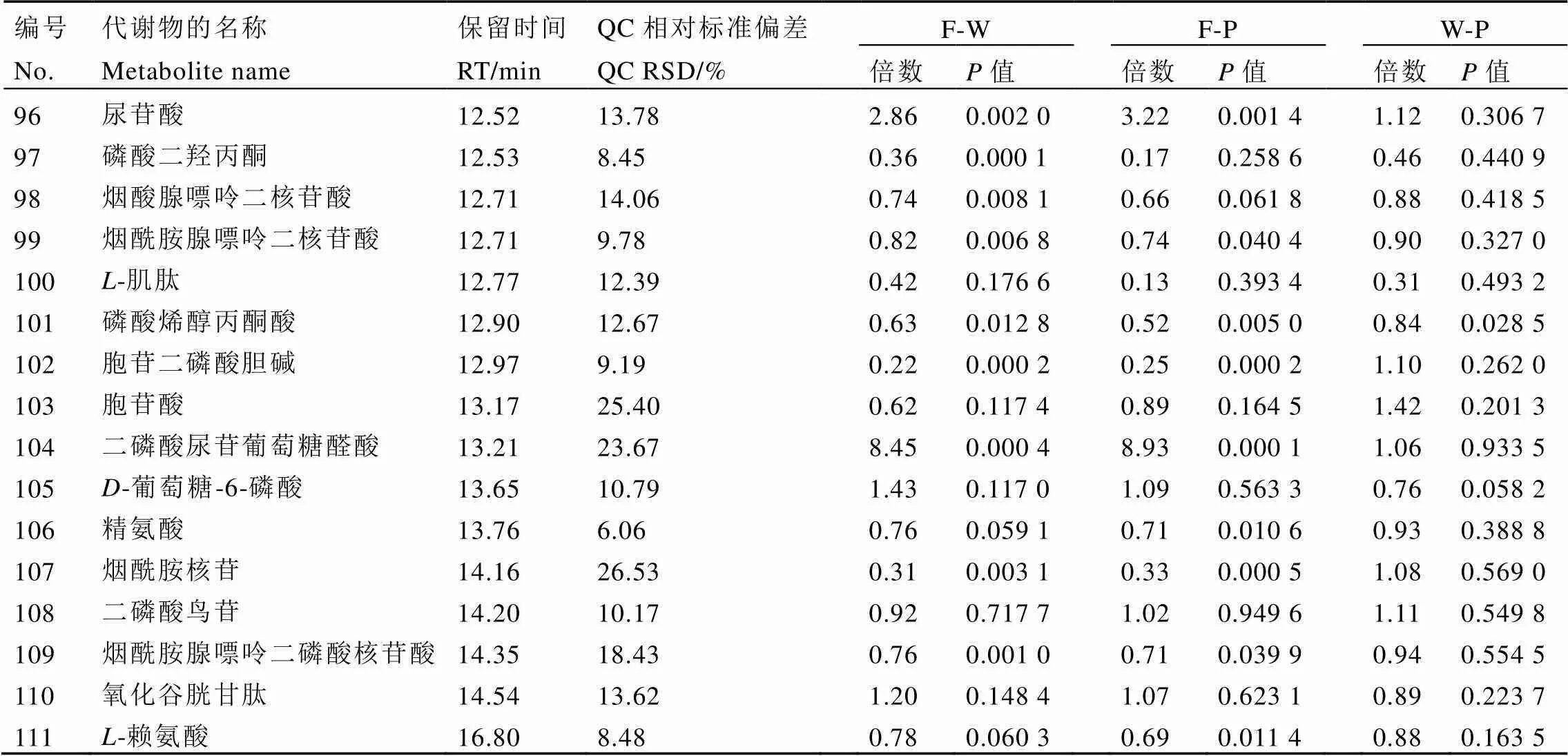
续表1
2.2.2 核酸代谢物
共计29种,按分子结构特点分为嘌呤、嘧啶两组。
(1)嘌呤代谢物(20种):①腺嘌呤类(8种):腺嘌呤*,2'--甲基腺苷*(存在于rRNA中)、5'-脱氧腺苷*、腺苷、N6-甲基腺苷,脱氧腺苷酸(dAMP)*、环磷酸腺苷酸(cAMP)*、腺苷酸(AMP),②鸟嘌呤类(5种):鸟苷*、脱氧鸟苷、7-甲基鸟苷、2,2-二甲基鸟苷、二磷酸鸟苷(GDP),③次黄嘌呤类(3种):次黄嘌呤*、肌苷*(次黄嘌呤核苷)、肌苷酸(IMP),④黄嘌呤类(2种):7-甲基黄嘌呤、黄嘌呤,⑤降解产物(2种):尿酸、尿囊酸。
(2)嘧啶代谢物(9种):①胞嘧啶类(5种):胞嘧啶、4-乙酰胞嘧啶、胞苷*、脱氧胞苷、胞苷酸(CMP),②胸腺嘧啶类(1种):胸腺嘧啶,③尿嘧啶类(2种):尿苷*、尿苷酸#(UMP),④其他(1种):假尿嘧啶核苷。
2.2.3 维生素和辅酶
共计12种,分别为:烟酰胺核苷酸*、烟酰胺腺嘌呤二磷酸核苷酸(NADP)、烟酰胺腺嘌呤二核苷酸(NAD)、烟酸*(VB3)、烟酸腺嘌呤二核苷酸(NAAD),核黄素(VB2)、黄素腺嘌呤二核苷酸(FAD)、黄素单核苷酸(FMN)、泛酸*(VB5)、吡哆醛、4-吡哆酸、吡哆醇。
2.2.4 糖代谢物
共计17种,主要涉及糖酵解、三羧酸循环、糖醛酸代谢途径的化合物和糖及其衍生物,据此分为4组。
(1)糖酵解途径代谢物(5种):-葡萄糖-6-磷酸、磷酸二羟丙酮、甘油-3-磷酸、磷酸烯醇丙酮酸(PEP)*、-乳酸。
(2)三羧酸循环代谢物(5种):柠檬酸、-乌头酸、琥珀酸#、延胡索酸、苹果酸。
(3)糖醛酸途径(3种):2-酮基--葡糖酸、二磷酸尿苷葡萄糖#(UDP-Glu)、二磷酸尿苷葡萄糖醛酸#(UDP-GluUA)。
(4)糖及其衍生物(4种):蔗糖#、肌醇*(半乳糖代谢产物)、-乙酰基-D-葡糖胺*、柚苷。
2.2.5 甘油磷脂代谢物
共计6种,包括甘油酸*、胆碱*、甜菜碱、-肉毒碱、胞苷二磷酸胆碱*(CDP-胆碱,细胞膜结构性磷脂生物合成过程中的一种必不可少的中间产物)和甘油磷酸胆碱。
上述所检测的生化成分主要涉及蛋白质、DNA、RNA相关的氨基酸、核苷酸类化合物及其衍生物,同时还包括与基础代谢相关的维生素和辅酶、糖代谢(糖酵解、TCA、葡糖醛酸)以及细胞膜成分相关的生化成分。
2.3 显著差异的代谢物筛选
显著差异代谢物的筛选标准为-value<0.05且Fold change>1.5或<0.67。一般来说,当筛选的候选代谢物合理且准确时,同组样能够通过聚类出现在同一簇(Cluster)中。同时,聚在同一簇内的代谢物具有相同的表达模式,可能在代谢过程中处于较为接近的反应步骤中。依据上述标准筛选出F和W显著差异物48种(图1)、F和P显著性差异物52种(图2)、W和P显著性差异物5种(图3)。
由图1可知,与鲜叶F相比,萎凋29 h的W样品中相对含量显著增加的代谢物有39种,增幅由大至小依次为:-乙酰基--酪氨酸、甘油酸、乙酰基-D-葡糖胺、腺嘌呤、乙酰--亮氨酸、CDP-胆碱、5'-脱氧腺苷、-2-氨基己二酸、dAMP、鸟苷、烟酰胺核苷酸、尿苷、2'--甲基腺苷、烟酸、胞苷、肌醇、cAMP、次黄嘌呤、磷酸二羟丙酮、-乙酰基--苯丙氨酸、苯乳酸、-乙酰尸胺、Tyr、Asn、GABA、胆碱、-腺苷高半胱氨酸、对羟基苯甲酸、二甲基甘氨酸、肌氨酸、肌苷、甲基胍、N6-甲基腺苷、核黄素、Leu、泛酸、PEP、Val、7-甲基黄嘌呤。其中,氨基酸代谢物16种(蛋白质氨基酸4种、氨基酸衍生物11种、非蛋白氨基酸1种)、核酸代谢物12种、维生素和辅酶4种、糖代谢4种、甘油磷脂代谢物3种。W中相对含量显著减少的代谢物有9种,降幅度由大至小依次为:UDP-GluUA、UDP-Glu、Met、UMP、蔗糖、尿酸、Glu、4-胍基丁酸、琥珀酸。其中,氨基酸类代谢物3种,核酸代谢物2种,糖代谢物4种。
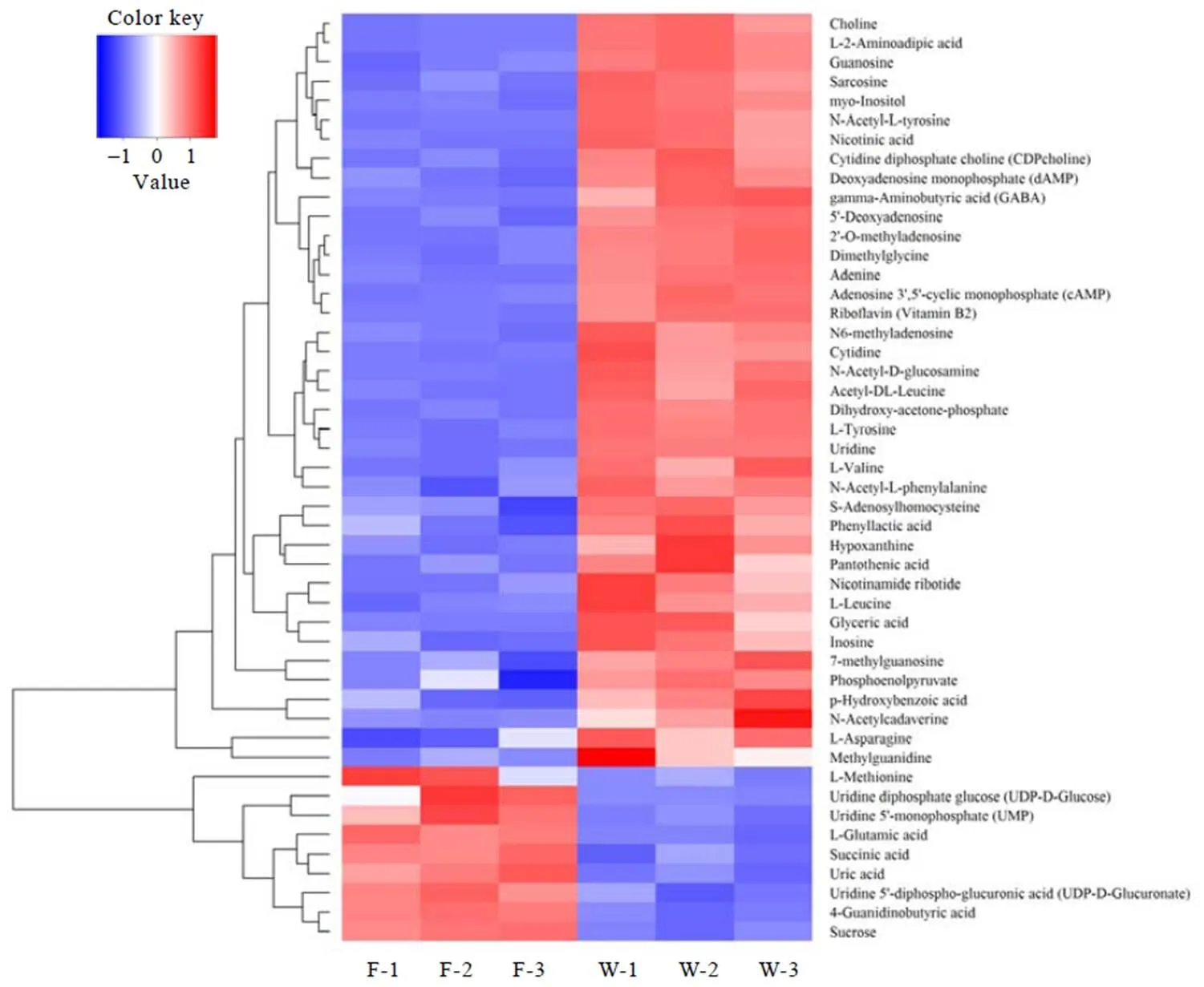
图1 F和W代谢物层次聚类结果
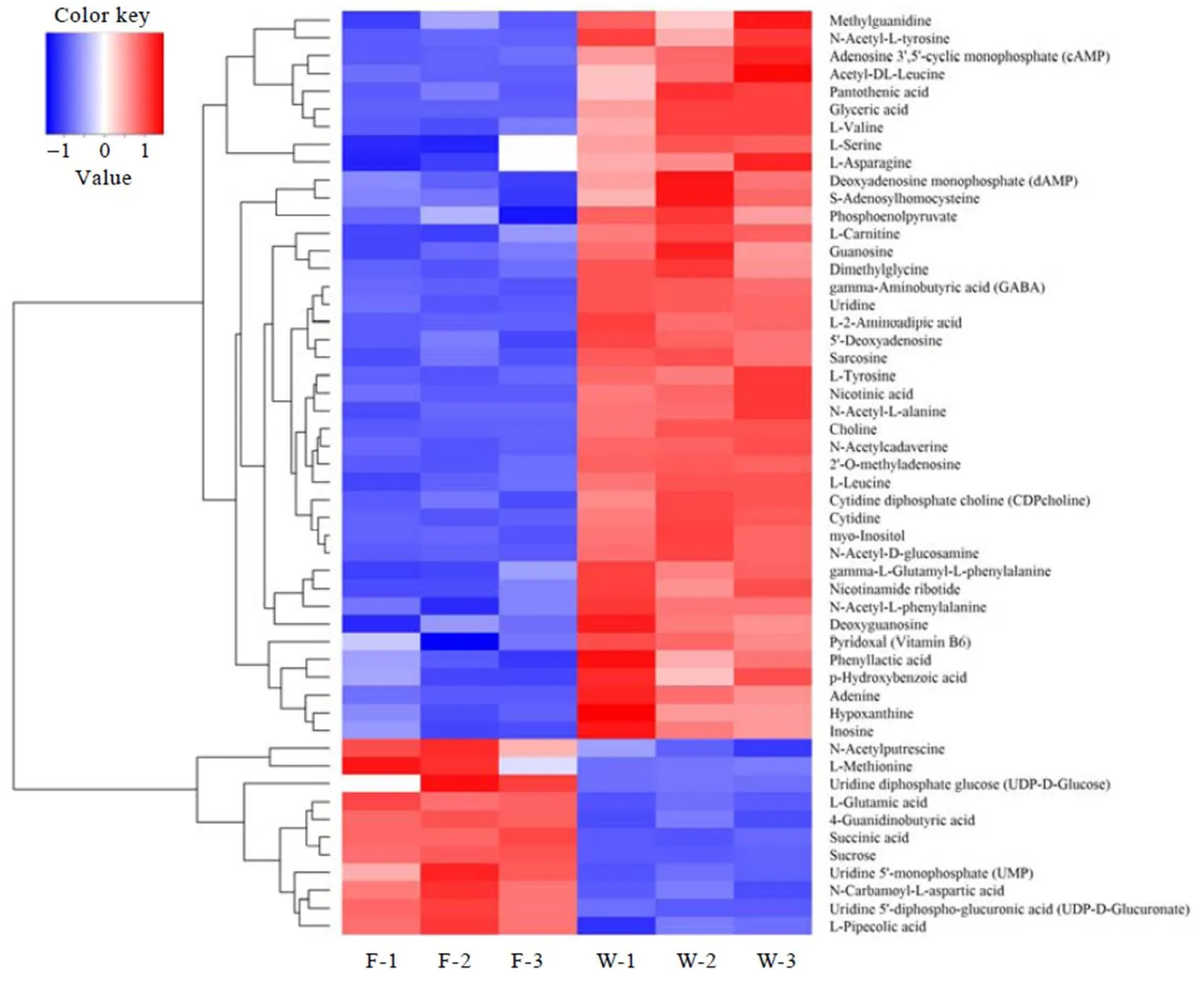
图2 F和P代谢物层次聚类结果
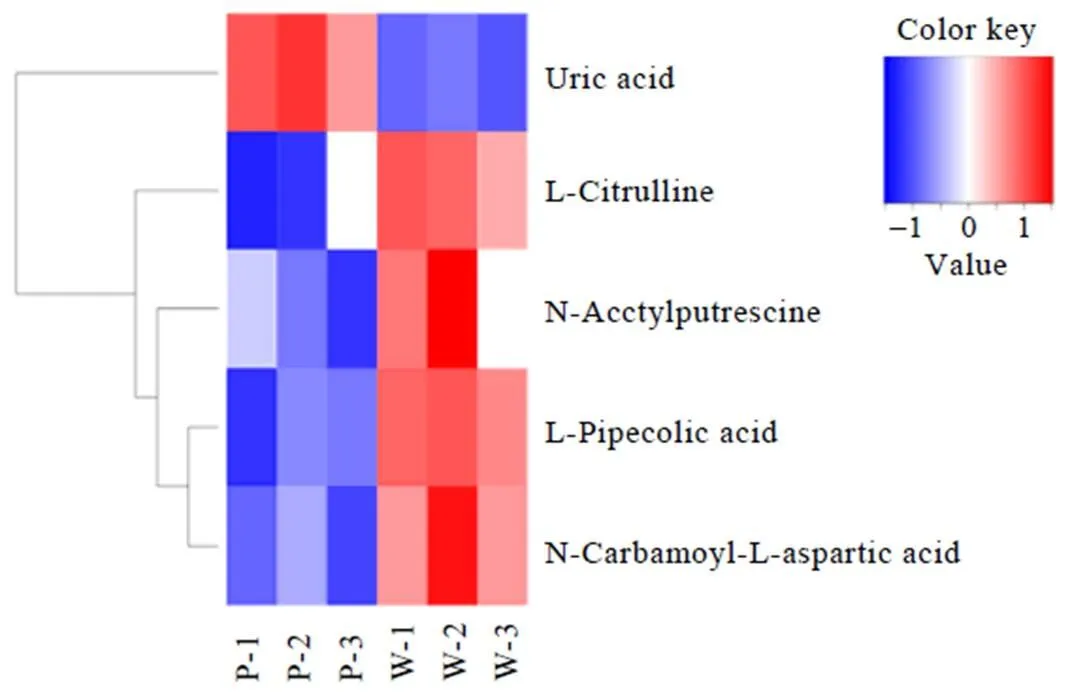
图3 P和W代谢物层次聚类结果
由图2可知,与鲜叶F相比,萎凋53 h的P中相对含量显著增加了41种代谢物,增幅由大至小依次为:甘油酸、-乙酰基--酪氨酸、乙酰基--葡糖胺、腺嘌呤、CDP-胆碱、乙酰--亮氨酸、5'-脱氧腺苷、-2-氨基己二酸、烟酰胺核苷酸、2'--甲基腺苷、--谷氨酰--苯丙氨、尿苷、肌醇、烟酸、dAMP、苯乳酸、Tyr、-腺苷高半胱氨酸、鸟苷、-乙酰基--苯丙氨酸、胞苷、胆碱、N-乙酰尸胺、次黄嘌呤、GABA、对羟基苯甲酸、cAMP、二甲基甘氨酸、Asn、肌氨酸、肌苷、PEP、泛酸、Leu、脱氧鸟苷、-肉毒碱、Ser、甲基胍、-乙酰基--丙氨酸、VB6、Val。其中,氨基酸类代谢物20种(蛋白质氨基酸5种、氨基酸衍生物13种、非蛋白氨基酸1种、寡肽1种)、核酸代谢物11种、维生素和辅酶4种、糖代谢物3种、甘油磷脂代谢物3种。萎凋叶P相对含量显著减少的代谢物有11种,降幅由大至小依次为:UDP-GluUA、UDP-Glu、Met、UMP、蔗糖、-氨基酰--天冬氨酸、琥珀酸、4-胍基丁酸、Glu、-乙酰腐胺、-哌啶酸,其中,氨基酸类代谢物6种,核苷酸1种,糖代物4种。
萎凋叶W、P与鲜叶F相比,相对含量显著增加的共有代谢物有35种,差异代谢物10种,其中,W中相对含量显著增加的差异代谢物有磷酸二羟丙酮、6-甲基腺苷、核黄素和7-甲基黄嘌呤4种;P中相对含量显著增加的6种差异代谢物分别为--谷氨酰--苯丙氨酸、脱氧鸟苷、-肉毒碱、Ser、-乙酰基--丙氨酸和VB6。相对含量显著减少的共有代谢物8个,差异代谢物4个,W中相对含量显著减少的差异代谢物为尿酸;P中相对含量显著减少的3种差异代谢物分别是-氨基酰--天冬氨酸、-乙酰腐胺和-哌啶酸。
由图3可知,与W相比,萎凋叶P中相对含量显著增加的代谢物仅有尿酸(0.36)1种,相对含量显著减少的代谢物有4种,主要为氨基酸及其衍生物,减少幅度由大到小依次为:瓜氨酸(1.88)、-哌啶酸(1.66)、-氨基酰--天冬氨酸(1.57)和-乙酰腐胺(1.51)。
3 讨论
3.1 白茶萎凋代谢特点及标志物筛选
本试验基于已有的HILIC-QqQ MS MRM技术为分析平台,该方法可以快速对32个KEGG通路中近200个代谢物靶向定量分析,但是未能对茶叶中主成分多酚类、生物碱类、茶氨酸进行测定,下一步需要补充数据库相关的样本量。
本研究结果表明,经过29 h和53 h萎凋处理的萎凋叶(W和P)与鲜叶相比,在所检测的111种代谢物中,含量显著增加的分别有39种和41种,显著降低的仅9种和11种。这些代谢物分别涉及氨基酸、核酸、维生素和辅酶、糖和甘油磷脂相关代谢途径,可以从多个维度反映白茶萎凋过程中细胞生理状态和代谢特点,主要有以下表现:
第一,细胞膜受损,甘油磷脂合成代谢受阻。甘油磷脂是生物体细胞膜结构的重要组成成分,同时也参与细胞膜对蛋白质的识别和信号传导。植物体内的甘油磷脂在磷脂酶D催化下发生磷酸与取代基团(如胆碱、肌醇、丝氨酸、甘油等)之间酯键的水解,从而释放出取代基团。在本研究所检测的代谢物中,与甘油磷脂相关的取代基化合物有胆碱、肌醇以及取代基丝氨酸、甘油的降解或氧化产物甘油酸,这3种代谢物在萎凋叶W和P中均显著增加,且上升幅度为:甘油酸>肌醇>胆碱。由此表明,细胞膜主要结构成分的甘油磷脂分子发生降解,从而导致细胞膜结构的损伤。甘油酸、肌醇和胆碱的含量变化可作为细胞膜生理状态的代谢标志物。此外,CDP-胆碱含量显著增加,甘油磷酸胆碱含量无显著变化,由此表明,甘油磷酸胆碱合成代谢受阻,从而导致中间代谢物累积。
第二,RNA、DNA降解。与鲜叶相比,萎凋叶P、W中嘌呤类共有代谢物8种,分别为腺嘌呤、5'-脱氧腺苷、2'--甲基腺苷、dAMP、cAMP、鸟苷、次黄嘌呤、肌苷;嘧啶类共有代谢物2种,分别为胞苷和尿苷,且这些共有代谢物均显著增加,其中以腺嘌呤类代谢物含量增加的种类较多,幅度较大;萎凋叶中含量显著下降的代谢物仅有UMP 1种。这可能与UMP存在于RNA之中,其降解速度较DNA快有关。此外,依据DNA、RNA分子组成特点和含量变化情况,可筛选出5'-脱氧腺苷、dAMP作为DNA降解的标志物,尿苷作为总RNA的降解标志物,2'--甲基腺苷作为rRNA降解的标志物,肌苷作为tRNA降解的标志物。
第三,氨基酸及其衍生物组成特点。(1)氨基酸衍生物种类多(25种),占氨基酸代谢物种类的53.19%,主要衍生化途径包括乙酰化、甲基化、氨基酰化、羟基化以及脱氨、脱羧等。其中,6种蛋白质氨基酸存在乙酰化形式,而甘氨酸则以二甲基甘氨酸、肌氨酸2种甲基化形式存在,且在萎凋叶中显著增加。(2)芳香族、碱性氨基酸及其衍生物增幅较大。如酪氨酸及其衍生物:-乙酰基--酪氨酸、对羟基苯甲酸;-苯丙氨酸的衍生物:-乙酰基--苯丙氨酸、苯乳酸;-赖氨酸的衍生物:-2-氨基己二酸、-乙酰尸胺;-精氨酸的衍生物:甲基胍;(3)含量显著降低的氨基酸仅谷氨酸和蛋氨酸2种。
第四,糖代谢异常。糖酵解途径中的PEP显著增加,表明中间代谢物累积;TCA途径中的琥珀酸显著减少,表明TCA途径受阻;糖醛酸途径UDPG-Glu、UDPG-GluUA显著减少,多糖合成受阻。此外,作为光合产物的蔗糖显著减少,不能为糖代谢提供必要的底物。
3.2 代谢谱分析可为白茶萎凋工艺参数确定提供依据
目前白茶加工萎凋工序的适度标准主要以失重率、感官指标并结合萎凋时间进行判断,虽然简单易行,但缺少从分子水平上进行判定的依据。
本研究应用代谢谱分析,不仅可以通过甘油磷脂及其降解物的含量变化了解细胞膜结构的生理状态,而且可以了解萎凋叶中与滋味相关的主要代谢途径以及代谢物的变化情况,从而可在分子水平上为白茶萎凋适度提供科学依据。本研究虽然选取了萎凋处理时间差异大的2个萎凋样W、P,但两者的代谢谱差异很小,在111个所检测的代谢物中仅有5个存在显著差异,说明29 h至53 h之间的萎凋处理对茶叶品质形成影响较小。究其原因,可能与泰安空气湿度低(25%~46%,平均36.6%),萎凋叶失水过快,处理29 h和53 h的2个萎凋样的含水量低且两者之间差异小密切相关。经测定,W的含水量为11.7%,P的含水量为8.1%,显著低于生物体进行正常生理代谢反应的水分范围。以上结果表明,通过对不同萎凋时间处理的茶样进行代谢谱差异显著性分析,可以了解萎凋叶代谢物的变化进程,从而有利于从分子水平判定萎凋工艺技术参数的合理性。
3.3 白茶萎凋代谢物对滋味的影响
在茶叶品质化学及形成机理研究中,目前普遍关注的是茶多酚及其氧化产物,其次为游离氨基酸,并提出了评判茶叶滋味物质协调性的“酚/氨比值”,明确了游离氨基酸是茶汤鲜度的贡献者[1]。在本项研究中,除检测到14种蛋白质氨基酸外,还检测到25种氨基酸衍生物,包括乙酰化、甲基化、氨基酰化以及脱氨、脱羧等多种途径产生的代谢物,其中有11种代谢物在萎凋叶中显著增加。这些代谢物对白茶滋味有何影响?目前尚未见报道,有待进一步研究。
在食品工业领域,呈味核苷酸被称为“超级味精”,虽然它们自身的鲜味微弱,且增加浓度对其鲜味提升作用不明显,但它们对鲜味及甜味、肉味和醇厚感有增效作用,对酸味、苦味、腥味和焦味等不良风味有消除或抑制作用[19]。呈味核苷酸的主要种类有肌苷酸、鸟苷酸、胞苷酸、尿苷酸和黄苷酸[20]。在白茶萎凋叶中虽然检测到29种核酸类代谢物,但与鲜叶相比显著增加的主要为核苷代谢物,如肌苷、鸟苷、胞苷和尿苷,而呈味的尿苷酸显著减少,其他呈味核苷酸类无显著变化。由此表明,现有白茶萎凋工艺还有待进一步优化。核酸经磷酸二酯酶水解为核苷酸,再由磷酸单酯酶水解为核苷和磷酸。为了使白茶中的鲜味核苷酸含量增加,尚需开展白茶萎凋过程中呈味核苷酸变化规律的研究。同时,该项研究还将为白茶萎凋时长判定和适度标准制订提供生化依据,因而具有一定的理论价值和实际指导作用。
[1] 施兆鹏, 黄建安. 茶叶审评与检验[M]. 4版. 北京: 中国农业出版社, 2010. Shi Z P, Huang J A. Tea evaluation and inspection [M]. 4th ed, Beijing: China Agricultural Press, 2010.
[2] 顾谦, 陆锦时, 叶宝存. 茶叶化学[M]. 北京: 中国科学技术出版社, 2002. Gu Q, Lu J S, Ye B C. Tea chemistry [M]. Beijing: Science and Technology of China Press, 2002.
[3] 杨伟丽, 肖文军, 邓克尼. 加工工艺对不同茶类主要生化成分的影响[J]. 湖南农业大学学报(自然科学版), 2001, 27(5): 384-386. Yang W L, Xiao W J, Deng N K. Effects of processing technology of different teas on the main biochemistry components [J]. Journal of Hunan Agricultural University, 2001, 27(5): 384-386.
[4] Fiehn O, Kopka J, Trethewey R N, et al. Identification of uncommon plant metabolites based on calculation of elemental compositions using gas chromatography and quadrupole mass spectrometry [J]. Analytical Chemistry, 2000, 72(15): 3573-3580.
[5] 王凯. 基于稳定性同位素15N示踪和代谢谱分析技术的茶叶氮代谢研究[D]. 合肥: 安徽农业大学, 2012. Wang K.15N stable isotope tracer-based spectral analysis techniques and metabolism of nitrogen metabolism of tea plant [D]. Hefei: Anhui Agricultural University, 2012.
[6] 张磊, 曾仲大, 叶国注, 等. 基于气相色谱-质谱联用与液相色谱-质谱联用的非靶向代谢组学用于3类茶叶中化学成分分析[J]. 色谱, 2014, 32(8): 804-816. Zhang L,Zeng Z D, Ye G Z, et al. Non-targeted metabolomics study for the analysis of chemical compositions in three types of tea by using gas chromatography-mass spectrometry and liquid chromatography-mass spectrometry [J]. Chinese Journal of Chromatography, 2014, 32(8): 804-816.
[7] Zhang L, Ning L, Yan Y C, et al. Development of the fingerprints of crude Pu-erh tea and ripened Pu-erh tea by high performance liquid chromatography [J]. Journal of Chinese Pharmaceutical Sciences, 2011(20): 352-359.
[8] 李万春. 气质联用在不同茶叶品质鉴定中的应用[D]. 南京: 南京理工大学, 2012. Li W C. Application of GC-MS in different tea quality identification [D]. Nanjing: Nanjing University of Science and Technology, 2012.
[9] Van d H J J J, Akermi M, Ünlü F Y, et al. Structural annotation and elucidation of conjugated phenolic compounds in black, green, and white tea extracts [J]. Journal of Agricultural and Food Chemistry, 2012, 60(36): 8841-8850.
[10] 王秀梅. 祁门红茶加工过程中代谢谱分析及其品质形成机理研究[D]. 合肥: 安徽农业大学, 2012. Wang X M. Analysis of metabolism profile and its quality formation mechanism in the process of keemun black tea [D]. Hefei: Anhui Agricultural University, 2012.
[11] Lee J E, Lee B J, Chung J O, et al.1H NMR-based metabolomic characterization during green tea () fermentation [J]. Food Research International, 2011(44): 597-604.
[12] Tan J, Dai W, Lu M, et al. Study of the dynamic changes in the non-volatile chemical constituents of black tea during fermentation processing by a non-targeted metabolomics approach [J]. Food Research International, 2016, 79: 106-113.
[13] Dai W, Qi D, Yang T, et al. Non-targeted analysis using ultra-performance liquid chromatography-quadrupole-time of flight mass spectrometry uncovers the effects of harvest season on the metabolites and taste quality in tea (L.) [J]. Journal of Agricultural & Food Chemistry, 2015, 63(44): 9869-9878.
[14] 汤莎莎, 芦晨阳, 周君, 等. 基于电子鼻和HS-SPME-GC -MS技术解析乌牛早茶的挥发性风味物质[J]. 食品工业科技, 2018, 39(14): 223-230. Tang S S, Lu C Y, Zhou J, et al. Volatile flavor compounds of different Wu Niuzao leaves and different heating temperatures based on electronic nose and GC-MS [J]. Science and Technology of Food Industry. 2018, 39(14): 223-230.
[15] Pongsuwan W, Bamba T, Yonetani T, et al. Quality prediction of Japanese green tea using pyrolyzer coupled GC/MS based metabolic fingerprinting [J]. J Agric Food Chem, 2008, 56(3): 744-750.
[16] 戴宇樵, 吕才有. 代谢组学技术在茶学中的应用研究进展[J]. 江苏农业科学, 2019, 47(2): 24-28. Dai Y Q, Lv C Y. Research progress of application of metabolomics technology in tea science [J]. Jiangsu Agricultural Sciences, 2019, 47(2): 24-28.
[17] Dai W D, Xie D C, Lu M L, et al. Characterization of white tea metabolome: Comparison against green and black tea by a nontargeted metabolomics approach [J]. Food Research International, 2017, 96: 40-45.
[18] Cai Y P, Weng K, Guo Y, et al. An integrated targeted metabolomic platform for high-throughput metabolite profiling and automated data processing [J]. Metabolomics, 2015, 11(6): 1575-1586.
[19] 崔桂友. 呈味核苷酸及其在食品调味中的应用[J]. 中国调味品, 2001, 16(10): 25-29.Cui G Y. Favor nucleotides and its application in food seasoning [J]. Chinese Condiment, 2001, 16(10): 25-29.
[20] 沈浥. 调味品中呈味核苷酸的研究进展和我国标准化现状[J]. 中国调味品, 2017, 42(1): 177-180. Shen Y. Research progress of the flavor nucleotides in condiments and standardization status in China [J]. China Condiment, 2017, 42(1): 177-180.
The Changes of Metabolites during the Withering Process of White Tea Based on HILIC LC-QqQ MS Method
CHEN Xiang, TIAN Yueyue, ZHANG Lixia*
College of Horticulture and Engineering, Shandong Agricultural University, Tea Engineering and Technological Research Center, Tai'an 271018, China
In this study, the metabolic profiles of fresh leaves (F), two indoor withered samples for 29 h (W) and 53 h (P) respectively were investigated by HILIC-QqQ MS based targeted metabolomics. Totally 111 metabolites were detected in the QC sample, including 47 amino acids and their derivatives, 29 nucleic acid metabolites, 12 vitamins and coenzymes, 17 sugar metabolites and 6 glycerophospholipid metabolites. Compared with the fresh leaves (F), 39 and 41 metabolites were significantly increased in withered samples W and P, with 35 overlapped metabolites. On the other hand, 9 and 11 metabolites were dramatically reduced, with 8 overlapped metabolites. The metabolite changes suggested that the cell membranes were damaged, DNA and RNA were degraded, amino acids derivatives were significantly increased and glucose metabolism was abnormal. Furthermore, according to the variations of metabolite contents and compositions, the marker metabolites of the withered leaves were identified which could reflect the physiological states of cell membrane, nucleic acid and glucose metabolism. Eventually, based on the characteristic changes in amino acids and nucleotides, the necessities of characterizing the flavor properties of amino acids and their derivatives as well as exploring the possibility of using metabolic profiling as white teawithering standardswere proposed.
white tea, withering, targeted metabolic profiling

S571.1;O657.63
A
1000-369X(2020)02-238-12
2019-08-30
2019-12-23
山东省高校“双一流”建设团队(SYL2007YY03)
陈翔,男,硕士研究生,主要从事茶叶加工与品质化学方面的研究,592063686@qq.com。*通信作者:lxzhang@sdau.edu.cn
投稿平台:http://cykk.cbpt.cnki.net
猜你喜欢
陕西师范大学学报(自然科学版)(2022年5期)2022-11-09 10:42:10
世界科学技术-中医药现代化(2022年3期)2022-08-22 00:33:26
肝博士(2022年3期)2022-06-30 02:48:28
云南化工(2020年4期)2020-05-19 09:15:24
科海故事博览·中旬刊(2020年3期)2020-03-15 05:49:04
Journal of Sport and Health Science(2019年6期)2019-11-26 07:30:53
安徽化工(2018年5期)2018-10-23 03:22:34
中学生数理化·高二版(2016年3期)2016-12-26 09:36:58
合成化学(2015年10期)2016-01-17 08:56:26
华东师范大学学报(自然科学版)(2014年1期)2014-04-16 02:54:58
