
融合表达乙醇酸氧化酶和过氧化氢酶转基因马铃薯的表型分析
2020-04-20 11:59:10
种子 2020年3期
(长江大学园艺园林学院,湖北 荆州 434025)
马铃薯(SolanumtuberosumL.),茄科一年生草本植物,是继水稻、小麦和玉米之后的第四大粮食作物,我国马铃薯种植面积居世界首位[1]。目前马铃薯的应用研究主要集中于粮食加工、抗病、脱毒、品质等方面,由于世界人口增长、全球财富增加和可耕地减少,马铃薯的生物工程改良显得越来越重要[2,3]。
光合作用是物质生产与产量形成的生理基础[4],而光呼吸是所有进行光合作用碳固定的细胞在光照下发生的一个生化过程,此过程消耗氧气,生成二氧化碳,是光合作用损耗能量的一个副反应[5,6]。预测表明,在未来大气中的CO2浓度达到最高水平时,仍可以减少植物光呼吸使光合速率上调12%~55%[7,8]。所以,通过构建光呼吸支路来提高光合效率和产量的研究逐渐引起重视[9,10]。目前,通过在植物叶绿体和过氧化物酶体内引入外源酶在拟南芥、水稻等植物中共建立了4条光呼吸支路,部分支路起到了使植物的光呼吸减少,光合作用和生物量产量增加等积极效果[11-14]。
乙醇酸氧化酶(Glycolate oxidase,缩写GLO)存在于过氧化氢酶体,是参与光呼吸过程中的关键酶。在光呼吸代谢支路中,其通常被叶绿体定位信号肽引入叶绿体表达,分解光呼吸的第一个产物乙醇酸,生成乙醛酸(Glyoxylate)与过氧化氢(H2O2)。过氧化氢酶(Catalase,缩写CAT)是分解H2O2避免其损害细胞的一种保护酶,Maier等[12]使用大肠杆菌的过氧化氢酶清除GLO分解乙醇酸产生的H2O2,在叶绿体中成功构建了光呼吸支路;Shen等[14]也在水稻叶绿体构建光呼吸支路时使用水稻的过氧化氢酶OsCAT2清除H2O2,获得了良好的效果。
本研究在马铃薯叶绿体中构建光呼吸支路,为了协调GLO和CAT的表达,将2个酶融合在1个表达盒中表达。融合表达的2个酶彼此空间距离近,由1个启动子启动,能实现最高效率的协调表达。通过分析2种表型的马铃薯转基因植株株高、有效叶面积、叶数、薯重等指标,有效评估了融合GLO、CAT分流乙醇酸,构建光呼吸支路的实际效益。
1 材料与方法
从水稻cDNA中扩增OsCAT2(AK 062174)和OsGLO3(AK 068638)编码序列,通过1个编码12个甘氨酸和3个丝氨酸(Gly 4 Ser)3接头的DNA序列(5′GGTGGTGGCGGTTCTGGTGGTGGCGGTTCTGGTGGTGGCGGTTCT),将2个基因的编码区融合在一起,并在编码的N端接入马铃薯rbcS叶绿体定位信号肽。将融合蛋白的表达盒接入到多基因转化载体pBIA 13(华南农业大学彭新湘研究员课题组赠送),通过烟草花叶病毒35 S启动子启动融合蛋白的表达。将重组后的质粒pBI-StTP-58 AA-CATGLO导入根癌农杆菌LBA 4404。
利用农杆菌介导法转化马铃薯(鄂马铃薯3号),以及通过DNA水平和蛋白水平表达分析,筛选得到4个转基因株系,分别为B 11、B 14、B 17、B 27。转基因植株在MS培养基上快速繁殖,当试管苗长至高2.5~3.0 cm,长1~2 cm新根时,移栽到泥炭土中种植。
同时在试管苗期,在MS培养基统一接种带芽茎段,每株系接种4瓶,每瓶接种5个茎段,重复4次,30 d后统计瓶苗的高度。选取第一代种植收获的大小均匀一致的薯块作为种薯,每个株系种6盆,每盆1株,随机排列。
在块茎膨大后期,统计株高及有效叶数,取相同部位的功能叶片若干,使用万深LA-S全能型植物图像分析仪测定其叶形和叶面积。收获时用称重法测定产量,按薯重的大小,将其划分为大、中、小薯3个标准:单株薯重大于20 g为大薯,10~20 g为中薯,小于10 g为小薯。
2 结果与分析
2.1 马铃薯GLO-CAT转基因植株出现2种表型
在试管苗阶段,转基因株系B 14的试管苗高高于野生型5%,B 11降低0.4%;而株系B 17,B 27相对于野生型显著降低,分别降低64%、39%(图1 A);对块茎繁殖的第二代植株进行观察分析,转基因株系B 14的株高高于野生型3%,B 11降低9%;而株系B 17、B 27相对于野生型显著降低,分别降低52%、38%(图1 B,图2 B)。
同时,转基因株系B 17和B 27出现叶片扭曲变厚、叶色变深、小叶相连等现象;B 11、B 14叶形、叶色和小叶数无明显差异(图2 A)。株系B 11、B 14的叶面积与野生型相比分别增加29%和15%,而B 17、B 27分别减少25%和15%(图1 C)。相对于野生型,株系B 11的有效叶数增加3%,B 14增加27%,而B 17和B 27显著减少,分别减少31%和50%(图1 D)。
转基因株系B 14的株高、叶面积、有效叶数与野生型相比均有所提高,为第1种表型;而B 27和B 17株高,叶面积、有效叶数均有所降低,且出现叶片畸形的现象,为第2种表型。
2.2 GLO-CAT转基因植株薯重的分布及形态改变
研究发现,转基因株系B 11、B 14和B 17收获的块茎外部形态与野生型无明显差异,而B 27的块茎则出现畸形,表现为多个或大或小的块茎发育在一起(图3 A)。
对块茎繁殖的第二代植株块茎进行薯重分析,转基因株系B 14的单株薯重相对于野生型显著上升,增加23%,B 11降低19%;而B 17和B 27的单株薯重显著下降,分别降低77%和76%(图3 B)。大于20 g的块茎比例(大薯率),转基因株系B 14为47%,野生型为20%,B 11与野生型接近,而B 27和B 17低于野生型,分别为13%和0%;10~20 g的块茎比例(中薯率),转基因株系B 11为24%,野生型为13%,其余株系略低于野生型;小于10 g的块茎比例(小薯率),野生型为67%,株系B 11、B 14低于野生型,分别为57%和43%,而B 27、B 17高于野生型,分别为77%和89%(图3 C)。

注:A为叶片;B为单株。图2 块茎繁殖的第二代植株的表型
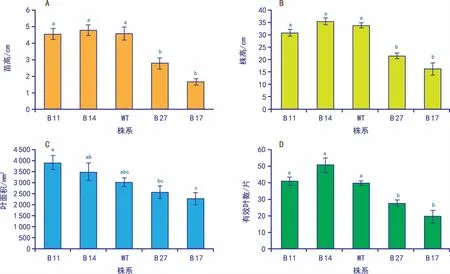
注:A为试管苗高度分析(误差线为标准误,小写字母表示0.05水平差异显著性。下同);B为块茎繁殖的第二代植株高度分析;C为块茎繁殖的第二代植株叶面积分析;D为块茎繁殖的第二代植株有效叶数分析。图1 GLO-CAT转基因植株的形态指标
转基因株系B 14的单株薯重、大薯率与野生型相比有增加的趋势;而转基因植株B 17和B 27薯重明显降低,小薯偏多。
3 讨 论
3.1 GLO-CAT融合表达对马铃薯代谢支路建立有积极的意义
目前,通过构建光呼吸支路来提高作物的生物量和产量,是极其有效的途径之一。Kebeish等[11]在拟南芥叶绿体中引入大肠杆菌乙醇酸分解代谢途径,成功减少了光呼吸代谢物通过过氧化物酶体和线粒体的通量,使其转基因植物生长得更快,产生更多的根茎生物量;Shen等[14]通过在水稻叶绿体中引入3种水稻自源酶,将乙醇酸完全氧化成CO2,从而使转基因水稻植株的光合效率、生物量产量和氮素含量显著提高。在本研究中,转基因株系B 14的叶形、叶色和小叶数与野生型无明显差异,但叶面积、有效叶数、株高,单株薯重均明显提高,与上述研究的结论相一致。说明在马铃薯叶绿体中引入GLO-CAT融合蛋白构建光呼吸代谢支路可行,通过分流乙醇酸、降低光呼吸,有效的增加了生物量和产量。

注:A为块茎形态;B为块茎单株鲜重;C为薯重分布(大薯≥20 g,20 g>中薯>10 g,小薯≥10 g)。图3 块茎繁殖的第二代植株块茎形态及薯重
研究也发现了一种负的表型,转基因株系B 27和B 17出现叶片变短、小叶数变少、叶色变深、叶片弯曲和薯重降低、小薯多等现象,这并不是光呼吸支路带来的优势。有研究指出,马铃薯的品质和产量性状是受复杂的微效多基因控制的数量性状[15]。张先文[16]在水稻的转基因研究中,综合考虑了有效光合面积、株高等的变化。所以在马铃薯光呼吸代谢支路建立后,也应该考虑其他限制因子的变化。同时,GLO和CAT的适度表达,也是构建光呼吸支路的重要因素。如果GLO活性过高,CAT活性太低,不能将GLO分解乙醇酸产生的H2O2迅速的分解,将会影响其他代谢通路,抵消在叶绿体中分解释放CO2产生的积极作用。本研究采用了融合蛋白的表达方式,有利于GLO和CAT的协调表达。本研究中,B 27和B 17这2个转基因株系的叶片出现畸形的现象,可能的原因是融合蛋白的表达量过高,超过了CAT清理限度,叶绿体中的H2O2未能及时清除所致。
3.2 GLO-CAT融合表达对马铃薯生物量和产量的影响
光合作用主要发生在叶片中,通常叶片被称为狭义的源,生长发育中的果实是光合产物积累的主要库,生产中可以通过协调源库之间的关系,达到源库平衡以实现作物的高产[17]。张绍荣等[18]证明,马铃薯产量随光合速率与单株叶面积之积的增大而增加,两者呈正相关。李文婷分析指出,马铃薯叶片积累同化物越早、越多、时间越长,源的活性就越大,产量也越高[19]。康朵兰等也提出,马铃薯的干物质含量、块茎产量、块茎数量和单株叶面积与光合速率呈正相关关系[20]。Nolke等[21]将大肠杆菌糖基脱氢酶(GlcDH)3个亚基(D、E、F)组成的重组蛋白(DEFp)导入马铃薯叶绿体,降低了光呼吸并导致茎叶生物量显著增加,以此增加了光合速率和叶面积,使块茎产量提高了2.3倍。说明了光呼吸代谢支路所造成的光合作用变化会直接影响作物产量。本研究中,转基因株系B 14的株高、叶面积、有效叶数高于野生型,致使单株薯重、大薯率明显提高,表明马铃薯因块茎碳汇能力强而具有更大的转基因优势。而B 27、B 17产量降低,可能由于单株的叶面积和有效叶数太少所致。
4 结 论
本研究通过融合表达乙醇酸氧化酶和过氧化氢酶,使转基因株系B 14的单株薯重显著高于野生型,株高、有效叶数和叶面积均有所提高,说明融合蛋白成功分流了乙醇酸,降低了光呼吸、提高了光合作用,生物量的增加可以表现在块茎中。同时研究发现,由于叶片形状、有效光合面积、有效叶数、株高等因子的变化,转基因株系B 27和B 17出现块茎畸形和薯重降低的现象。这一现象,在光呼吸代谢改造中不应忽视。
猜你喜欢
浙江农业科学(2024年4期)2024-05-07 04:41:08
中国烟草科学(2022年2期)2022-05-27 10:05:35
亚热带植物科学(2022年1期)2022-05-17 12:39:32
植物营养与肥料学报(2021年10期)2021-12-05 04:26:46
四川农业科技(2019年5期)2019-07-01 09:46:46
农家顾问(2016年10期)2016-11-21 08:07:24
广西林业科学(2016年1期)2016-03-20 05:33:01
浙江柑橘(2016年1期)2016-03-11 20:12:31
食品科学(2013年6期)2013-03-11 18:20:13
中国烟草学报(2012年4期)2012-04-09 07:11:42
