
不同环境因子及破壳处理对锥栗种子萌发特性的影响
2020-04-17 08:58李吉涛陈宗游唐辉王满莲韦霄
广西植物 2020年2期
李吉涛 陈宗游 唐辉 王满莲 韦霄
摘要: 锥栗(Castanea henryi)是我国南方重要的木本粮食树种和特色果树资源,具有很高的食用及药用价值。为探明最适宜锥栗种子萌发的条件,该研究以当年采收的锥栗种子为材料,探讨了温度、光照、基质、基质含水量以及破壳处理等对锥栗种子萌发的影响,为锥栗种苗繁育提供理论基础。结果表明:(1)锥栗种子萌发的最适温度是25 ℃,其最终萌发率为92.67%,在其他温度条件下,锥栗种子的萌发率有所下降,但与25 ℃条件下的萌发率无显著差异。(2)黑暗条件可缩短锥栗种子发芽时间,明显提高种子的萌发率和发芽势,但黑暗条件下的种子胚根长度显著小于周期性光照条件下的胚根长度。(3)锥栗种子在泥炭土中的萌发率和发芽势均高于在河沙中的萌发率和发芽势,且泥炭土中种子胚根长度显著长于河沙中的种子胚根长度。(4)基质含水量对锥栗种子萌发率和发芽势无显著性影响。(5)种皮的机械阻碍是限制锥栗种子萌发的因素之一,破壳处理后,锥栗种子的发芽势和胚根长度与未破壳相比差异显著,实际生产中剪破种壳能加快种子的萌发进程。
关键词: 锥栗, 种子, 环境因子, 破壳处理, 萌发
中图分类号: Q945文献标识码: A
文章编号: 1000-3142(2020)02-0143-08
Abstract: Castanea henryi has very high edible and medicinal values, which is an important woody grain and a characteristic fruit tree resource in South China. In order to find out the best treatment strategy to improve the germination rate of C. henryi seeds, and provide scientific references for its exploration and utilization, effects of temperature, light, matrix, matrix moisture and shell-breaking treatment on seed germination of C. henryi were investigated. The results were as follows: (1) The suitable temperature for germination was 25 ℃, the final germination percentage was 92.67%, and the germination percentage under other temperature conditions decreased, but had no significant differences with that under 25 ℃. (2) The germination percentage and germination energy under the dark regime was significantly higher than that under alternating light regime, but the root length under alternating light regime was significantly longer than that under the dark regime. (3) Peat soil had the best effect on seed germination and seedling growth, the germination percentage and germination energy in peat soil were higher than that in sand, and the root length in peat soil was significantly longer than that in sand. (4) The matrix moistures had no significant effect on seed germination percentage and germination energy. (5) The mechanical barrier of seed shell was one of the limiting factors of seed germination. After shell-breaking treatment, the seed germination energy and root length were significantly different from those without shell-breaking. Shell-breaking treatment could accelerate seed germination in actual production.
Key words: Castanea henryi, seed, environmental factors, shell-breaking treatment, germination
錐栗(Castanea henryi)又名榛子、珍珠栗、旋栗或棒栗,为壳斗科(Fagaceae)栗属(Castanea)植物,是我国南方著名的干果和木本粮食树种之一,为我国特色落叶果树资源(刘国华和方精云,2001)。锥栗具有很高的食用价值和药用价值。锥栗果实富含淀粉、可溶性糖、不饱和脂肪酸、核黄素、Vb、Vc及人体所必需的Ca、Fe、Mg、Mn等微量元素(郑诚乐等,2003),其蛋白质和脂肪含量与小麦相当,比稻米高2~3倍,碳水化合物含量高于小麦(宋爱云等,2001)。锥栗所含的不饱和脂肪酸和多种维生素,对高血压、冠心病和动脉硬化等疾病有较好的预防和治疗作用,栗内薄皮可去皱、美容,栗壳有止反胃、消渴、止泻血的功能,栗花可治疗颈淋巴结核,栗毛球和树皮有去毒、消肿之功效,栗根有治偏肾气的作用(景芸等,2004;郑诚乐,2008)。锥栗的壳斗、树皮含有大量的鞣质,可提取拷胶。锥栗是落叶乔木,实生树可高达30 m,树干比板栗和茅栗直,材质坚实、耐水湿,是枕木、建筑、家具、造船的优质经济树种;其根系发达,能固结、改良土壤,防止水土流失,是提高森林覆盖率、有效改善生态环境的优良树种(王海为,2003)。
种子是种子植物特有的延存器官,种子萌发与否对种子植物的生存和繁衍具有决定性作用(Rajjou et al., 2012),是它们完成种群扩散、占领新领地以及种群更新的极为关键的一步。种子萌发是植物对环境抵抗力最弱的阶段,其中会受到氧气、温度、阳光、水分等环境因子以及自身能力的综合影响,是植物为适应环境以保持自身的繁殖发展而形成的一种生物特性,具有重要的生态学意义。本文通过研究不同因素(温度、光照、基质类型、基质含水量、破壳处理等)对锥栗种子萌发的影响,探讨适合锥栗种子萌发的条件,以期为锥栗播种育苗及大量繁殖提供参考和理论依据。
1材料与方法
1.1 材料
锥栗种子于2016年10月采自广西灵川县海洋乡小平乐村山林的野生种群。种子采回经去杂清理后进行千粒重、种径测定,并置于温度为4 ℃、水分含量约4%的沙床进行沙藏保存,备用。
1.2 方法
种子千粒重是从干净种子中随机抽取1 000粒种子进行称重,重复三次。种径测定是随机抽取30粒种子进行纵径和横径的测定。种子萌发试验于2015年11月至2016年4月在室温条件和LRH-250-G光照培养箱(广东省医疗器械厂生产)条件下进行。
1.2.1 种子萌发特性测定实验前种子经清水搓洗,并以水选法选取饱满种子,用0.5% KMnO4浸泡消毒30 min,清水冲洗,滤干后备用。采用单因素试验,分别探讨不同温度、光照、基质、基质含水量及破壳处理五个试验因素对种子萌发的影响,萌发试验的所有处理均设三个重复,每重复50粒种子。种子置于培养皿中萌发,试验因素为基质的处理设河沙和泥炭土两种基质,其他试验因素的处理均用河沙作为培养基质;试验因素为基质含水量的处理设置5%、10%和20%三个不同的含水量梯度,其他試验因素处理的基质含水量约为10%,每隔一个星期根据培养皿失水情况补充水份,使各处理的基质保持相应的含水量。播种时,试验因素为光照处理的播种方式为坚果(种子)果球的一半露在基质外面,其他试验因素处理的播种方式为坚果(种子)的果球全部没入基质中。每30 d将萌发和霉烂的种子挑出培养皿,统计萌发的种子数(胚根伸出在5 mm以上即视为萌发)。每次统计时均对不同处理的所有新萌发种子的胚根进行长度和粗度的测定,试验结束时取其均值以了解不同的试验处理对胚根发育状况的影响。各试验处理设置如下:
(1)温度:设置15、20、25和30 ℃四个温度梯度,以室温作为对照。
(2)光照:室温条件下,设室内周期性光照(把培养皿置于靠近窗口的台面)和持续黑暗(0 lx, 24 h·d-1 ,用不透光的盖子罩住) 两个处理;25 ℃光照培养箱条件下,设置周期性光照(3 000 lx, 12 h·d-1)和持续黑暗(0 lx, 24 h·d-1,用不透光的盖子罩住)两个处理。
(3)培养基质:分别在光照培养箱(25 ℃、周期性光照3 000 lx, 12 h·d-1)和室内(室温、室内周期性光照)条件下,各设置河沙、泥炭土两种不同基质。
(4)基质含水量:分别在光照培养箱(25 ℃、周期性光照3 000 lx, 12 h·d-1)和室内(室温、室内周期性光照)条件下,各设置5%、10%、20% 三个不同含水量梯度。
(5)破壳处理:分别在光照培养箱(25 ℃、周期性光照3 000 lx, 12 h·d-1)和室内(室温、室内周期性光照)条件下,各设置种子破壳(在种子尖端把壳剪开一个小孔)和未破壳两个处理。
1.2.2 数据统计与处理以萌发率和发芽势为指标,在规定时间(60 d)内计算发芽势,在种子萌发试验终期(规定日期内,本研究为150 d)计算最终发芽率。发芽率和发芽势计算公式如下:
萌发率(GP)=正常发芽种子粒数/参试种子总粒数×100%;
发芽势(GE)=规定时间(60 d)内发芽的种子数/供试种子数×100%。
采用SPSS 13.0 软件作单因素方差分析(one-way ANOVA),用Duncan法进行多重比较,方差不齐的采用Tamhanes T2 进行比较;种子形态的描述性统计值以平均值±标准误表示。同时,基于平均值和标准误对有关发芽率、发芽势等用软件Sigmaplot 10.0作图。
2结果与分析
2.1 锥栗种子形态特征
锥栗坚果呈圆锥形或卵形,横径为(0.65±0.01)cm,纵径为(0.83±0.02)cm,种皮为红褐色。成熟坚果千粒鲜重为(3 300±84.98) g。
2.1 不同温度对锥栗种子萌发的影响
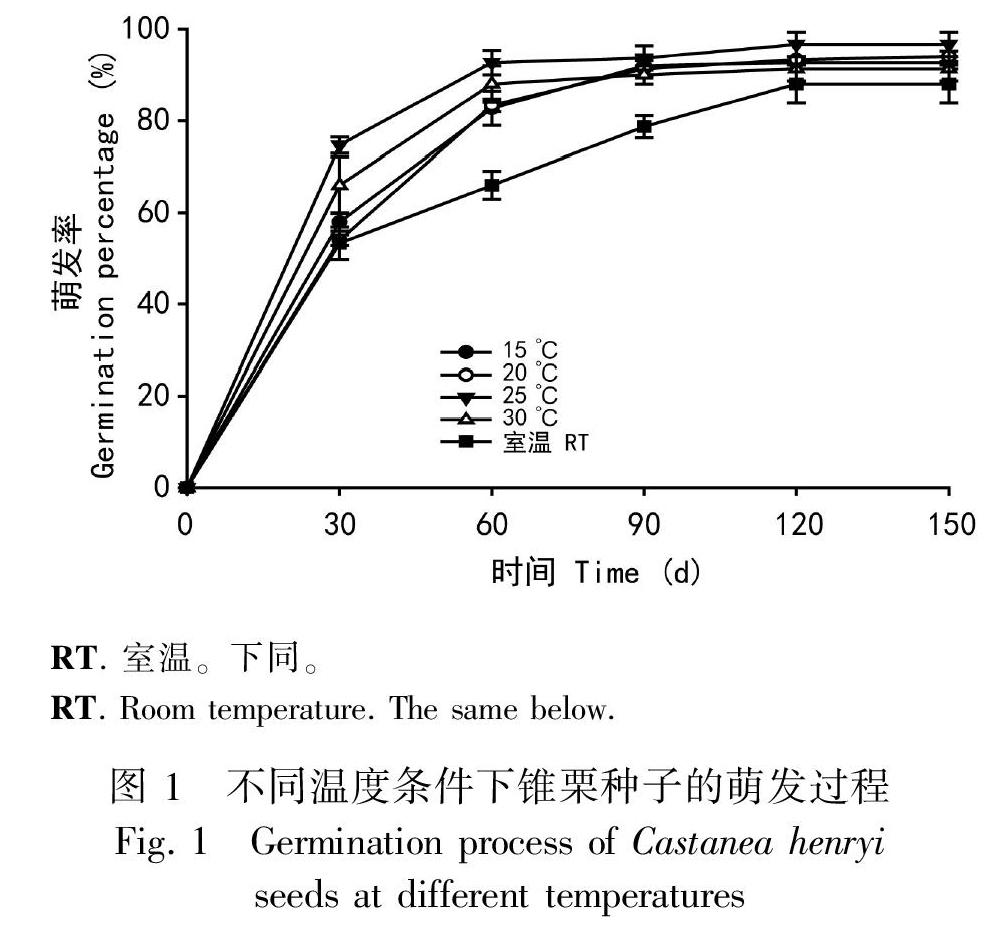
不同温度条件下,锥栗种子萌发过程存在差异。萌发前期(60 d内),温度对种子萌发率的影响较大,25 ℃的萌发率曲线斜率最大,萌发率增加量最大,30 ℃次之, 室温条件下最小,且25 ℃的萌发率分别高于20 ℃、15 ℃的萌发率。60 d后,室温条件下萌发率增加较多,其余几个温度萌发率逐渐趋于平稳。试验结束时,25 ℃萌发率最大,其次为15 ℃、20 ℃,30 ℃和室温的发芽率依次居后(图1)。对于发芽势,25 ℃和30 ℃的发芽势差异不显著,但它们显著高于15、20 ℃和室温的发芽势(图2:A)。五种温度条件中,25 ℃最适宜种子萌发,在播种后60 d内,随着温度的升高或降低,萌发率和发芽势都有所下降。不同温度条件下,RT. 室温。下同。
胚根粗度不存在显著性差异(P>0.05),但不同温度对胚根长度有影响(图2:B)。25 ℃的胚根长度显著长于其他温度的胚根长度(P<0.05),15 ℃、20 ℃、30 ℃的胚根长度两两之间差异不显著(P>0.05),但它们的胚根长度均显著长于室温的胚根长度(P <0.05)。
2.2 不同光照对锥栗种子萌发的影响
不同的光照条件,对锥栗种子萌发率有不同的影响。在25 ℃、黑暗条件下,种子萌发率在播种60 d后达到峰值,为92.67%,在25 ℃、周期性光照条件下萌发率上升较为平缓,最终萌发率不高,为79.33%;室温下,在种子萌发的早中期(120 d之前),黑暗下萌发率远高于周期性光照条件下的萌发率,直到试验末期(120 d)后,两者的累积萌发率才基本持平(图3)。相同的温度条件下,黑暗条件下的种子发芽势显著高于周期性光照条件下的种子发芽势(P<0.05)(图4:A)。从总体来看,黑暗条件下,种子的萌发率和发芽势明显高于光照下,黑暗更利于种子萌发。不同光照条件下,胚根粗度差异不显著(P>0.05),但胚根长度差异显著(图4:B)。在相同温度条件下,黑暗条件下的种子胚根长度均显著小于周期性光照条件下的种子胚根长度(P<0.05)。
2.3 不同基质对锥栗种子萌发的影响
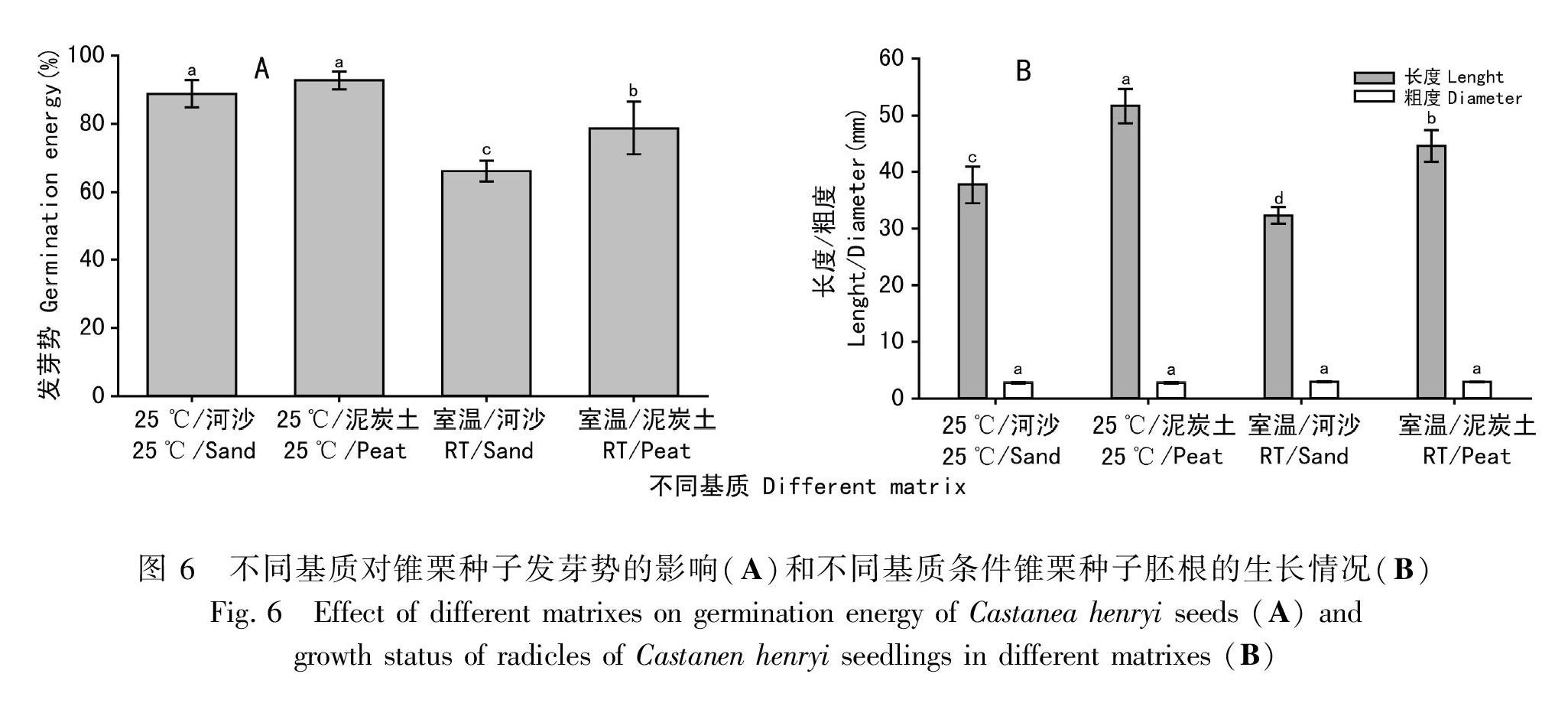
河沙和泥炭土对锥栗种子发芽的快慢和萌发率有不同影响(图5)。25 ℃、周期性光照条件下,图 3不同光照条件下锥栗种子的萌发过程60 d内,河沙、泥炭土中种子的萌发率均快速升高, 前者略高于后者。随着时间的推移,萌发率缓慢上升,在试验中后期(90 d后)达到顶峰,后者略高于前者。室温、周期性光照条件下,泥炭土中种子的萌发率高于同一阶段河沙中的萌发率(图5),25 ℃、周期性光照条件下,河沙中的种子发芽势略高于泥炭土,但它们之间并不存在显著性差异(P>0.05),而室温、周期性光照条件下,河沙中的种子发芽势显著低于泥炭土中的种子发芽势(P<0.05)(图6:A)。综合分析,泥炭土更适于种子萌发,可能由于泥炭土土质疏松,透气性好,且肥力较好。基质对胚根粗度的影响不显著(P>0.05),seeds in different matrixes而对胚根长度却有显著影响 (P<0.05)(图6: B)。在相同的温度条件下,沙河中的种子胚根长度显著小于泥炭土中的种子胚根长度(P<0.05)。
2.4 基质含水量对锥栗种子萌发的影响
不同基质含水量对种子萌发有一定的影响,在25 ℃、周期性光照条件下,含水量为5%、10%、20%基质中种子最终萌发率分别为84.44%、86.67%、81.11%,而在室温、周期性光照条件下相同含水量基质中种子的最终萌发率为88.89%、90%、84.44%(圖7)。对于发芽势,在相同温度、光照条件下,均以含水量为10%的基质中为最大,含水量20%的基质次之,5%基质中的最小,不同含水量基质中种子发芽势之间无显著性差异(P>0.05)(图8:A)。综上可知,锥栗种子萌发对基质图seeds in different matrix moistures含水量要求不严格,适应能力较强。基质含水量对胚根粗度不具有显著性影响(P>0.05);在25 ℃、周期性光照条件下,不同含水量基质中胚根长度无显著性差异(P>0.05);而在室温、周期性光照条件下,不同含水量基质中胚根长度却有显著性差异(P<0.05);在相同温度、光照条件下,均以含水量为20%基质中的胚根长度最大,含水量5%的次之,含水量10%的最小(图8:B)。
2.5 破壳处理对锥栗种子萌发的影响
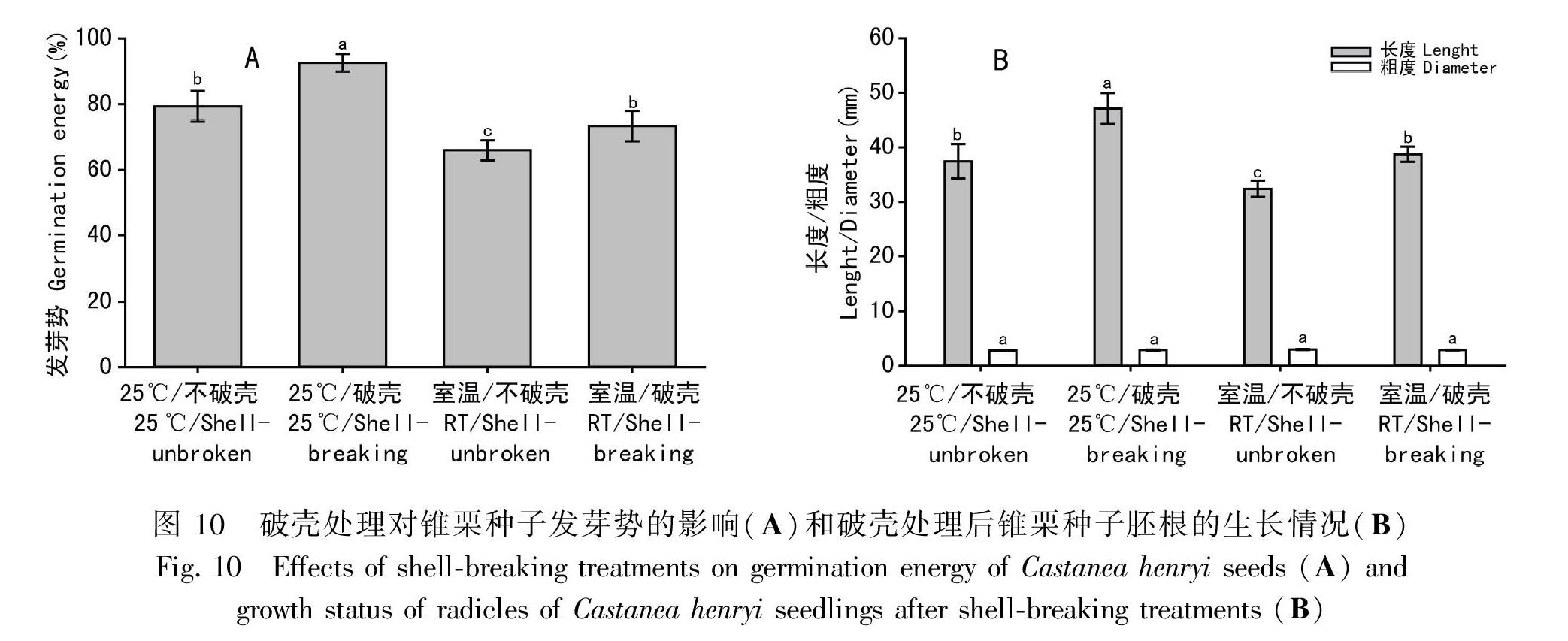
在25 ℃、周期性光照条件下,未破壳处理的种子在播种30 d后才开始萌发,晚于破壳种子,90 d后二者的累积萌发率相同,但发芽势显著低于破壳种子(P<0.05)。在室温、周期性光照条件下,破壳处理的种子的萌发率高于未破壳种子,破壳种子发芽势显著高于未破壳种子(P<0.05)(图9,图10:A)。由此可知,在相同温度、光照条件下,破壳与否可影响种子的萌发效果,打破外壳的束缚,使种子更容易收水分和氧气,进而有利于种子萌发。破壳与否对于胚根粗度不产生显著性影响(P>0.05),但在相同条件下,破壳种子的胚根长度显著大于未破壳种子(P<0.05)(图10:B)。
3讨论与结论
种子萌发需要充足的水分,适宜的温度,充足的氧气和适当的光照,但因生境和种类的不同,影响种子萌发的主要因子也有所不同。种子萌发过程受到温度、光照、基质、基质含水量等环境因子及播种前破壳处理的综合影响。
温度是影响种子萌发的关键因素之一,适宜的温度能够提高种子萌发率和促进幼苗生长(Ojeifo et al., 2007; 张小娇等,2014)。不同物种种子的萌发温度也不同,如海韭菜(Triglochin maritima)(Khan & Ungar, 1999)、坡柳(Dodonaea viscosa)(霍冬芳等,2016)等在高温下种子的萌发率较高,而低温能提高梭梭(Haloxylon ammodendron)(黄振英等,2001b)等植物种子的萌发率。本研究结果表明,锥栗种子萌发具有较好的环境适应性,萌发温差范围较大,且最适宜萌发温度为25 ℃,温度过高或过低对种子萌发均有不同程度的影响。这与白沙蒿(Artemisia sphaerocephala)种子类似(黄振英等,2001a)。
不同物种的种子萌发所需要的光照条件有所不同(杨期和等,2001)。锥栗种子在光照和黑暗条件下均能萌发,不管是在恒温条件下,还是在室温条件下,黑暗条件都更有利于锥栗种子萌发,符合杨期和等(2005)提出的锥栗种子萌发不需要光的结论,可能是因为锥栗种子比较大,所储藏的物质和能量较多,能满足种子萌发和幼苗生长所需的物质,这与Pamukov & Schneider(1978)的研究结果相似。
不同基质含有的营养物质、微生物种类及数量不同,其松紧程度也不同,从而影响种子的萌发、幼苗的生长(Puerta-Pinero et al., 2006)。在相对适宜的光温条件下,泥炭土中种子的萌发率高于河沙,这可能是由于泥炭土质地较轻、透气性好,氧气供给充足,减少种子霉变,有利于种子萌发,而河沙粒间孔隙小、通透性差、通气透水不良、质地重,因此种子容易发霉。由此可知,泥炭土更适于锥栗种子萌发。
水是种子萌发必备的条件之一,是控制種子萌发的最重要因素。种子萌发过程中需要从基质中吸收水分,从基质中进入种子的水分能溶解一些贮藏物质,使之成为种胚能够利用的养料,基质含水量的不同也会影响植物种子的萌发(林涛等,2008)。锥栗在含水量为5%~20%的基质中均能萌发,发芽率也无显著性差异,表明锥栗种子对基质含水量要求不严格,适应范围较广。
锥栗的外壳韧硬坚实且外层没有结合缝,透气性差、透水性差,导致萌发过程中吸水困难,影响种子萌发。人为打破种子外壳的束缚,减小种皮的阻碍作用,让种子直接处于容易吸收水分的状态下,使种子原生质吸收到足够水分而促使其所含的物质迅速活跃起来,从而促进种子发芽。破壳处理能提高锥栗种子的萌发率和发芽势,这与大别山五针松(Pinus dabeshanensis)种子类似(韩建伟等,2014)。
综上所述,锥栗种子最适萌发温度是25 ℃,黑暗条件更有利于种子萌发,种子萌发对基质含水量要求不严格,适宜以泥炭土等质地较轻、透气性好的基质作为萌发基质,在实际生产中,剪破种壳能提高种子的萌发率。
参考文献:
HAN JW, ZHANG ZY, WANG EM, et al., 2001. Pinus dabeshanensis seed characteristics and the promotion of seed germination [J]. Chin Agric Sci Bull, 30(1): 5-10. [韩建伟, 张智勇, 王恩茂, 等, 2014. 大别山五针松种子特性及促进种子萌发的研究 [J]. 中国农学通报, 30(1):5-10.]
HUANG ZY, GUTTERMA Y, HU ZH, et al., 2001a. Seed germination in Artemisia sphaerocephala II. The influence of environmental factors [J]. Chin J Plant Ecol, 25(2): 240-246. [黄振英, GUTTERMA Y, 胡正海, 等, 2001a. 白沙蒿种子萌发特性的研究Ⅱ. 环境因素的影响 [J]. 植物生态学报, 25(2):240-246.]
HUANG ZY, ZHANG SX, GUTTERMA Y, et al., 2001b. Influence of light, temperature and salinity on the seed germination of Haloxylon ammodendron [J]. Acta Phytophysiol Sin, 27(3): 275-280. [黄振英, 张新时, Gutterma Y, 等, 2001b. 光照、温度和盐分对梭梭种子萌发的影响 [J]. 植物生理学报, 27(3):275-280.]
HUO DF, HUANG BQ, SU WH, et al., 2016. Effects of high temperature treatment on seed germination of semi-Savanna species: Dodonaea viscosa (L.) Jacq [J]. Seed, 36(10): 10-18. [霍冬芳, 黄博强, 苏文华, 等, 2016. 萌发前高温处理对坡柳种子萌发率的影响 [J]. 种子, 36(10):10-18.]
JING Y, LIANG YC, YANG H, 2004. Comparison of fruit nutrition components of Castanea henryi clones [J]. J Zhejiang For Coll, 21(2): 176-179. [景芸, 梁一池, 杨华, 2004. 不同锥栗无性系果实营养成分的比较分析 [J]. 浙江林学院学报, 21(2):176-179.]
KHAN MA, UNGAR IA, 1999. Effect of salinity on seed germination of Triglochin maritima under various temperature regimes [J]. Great Basin Nat, 59(2): 144-150.
LIN T, TIAN YL, WANG Y, et al., 2008. Effects of moisture contents of medium, temperature conditionon seeds germination of Caragana korshinskii and other four psammophytes [J]. J Inn Mongol Agric Univ, 29(4): 30-33. [林涛, 田有亮, 王燕, 等, 2008. 基质含水量、温度条件对柠条等5种沙生植物种子萌发的影响 [J]. 内蒙古农业大学学报, 29(4):30-33.]
LIU GH, FANG JY, 2001. Spatial patterns of chestnut (Castanea millissima) and its species geographical distribution in China [J]. Acta Ecol Sin, 21(1): 164-170. [刘国华, 方精云, 2001. 我国栗属物种地理分布及其空间特征分析 [J]. 生态学报, 21(1):164-170.]
OJEIFO IM, EMUH FN, IGBENIJEOGOR B, 2007. Effect of different temperature regimes on seed germination and growth parameters of Dacryodes edulis [J]. Agric J, 2(1): 16-19.
PAMUKOV K, SCHNEIDER MJ, 1978. Light inhibition of Nigella germination: The dependence of a high irradiance reaction on 720-nm irradiance [J]. Bot Gaz, 139(1): 56-59.
PUERTA-PINERO C, GOMEZ JM, ZAMORA R, 2006. Species specific effects on topsoil development affect Quercus ilex seedling performance [J]. Acta Oecol, 29(1): 65-71.
RAJJOU L, DUVAL M, GALLARDO K, et al., 2012. Seed germination and vigor [J]. Ann Rev Plant Biol, 63: 507-533.
SONG AY, CHEN Q, LEI RP, et al., 2001. Analysis of content of nut nutrient elements in different cultivars of Henry chestnut Castanea henryi [J]. Econ For Res, 19(4): 5-7. [宋愛云, 陈钦, 雷日平, 等, 2001. 锥栗栽培品种果实营养成分差异的分析 [J]. 经济林研究, 19(4):5-7.]
WANG HW, 2003. Study on cultivars and community of Castanea henyi [D]. Fuzhou: Fujian Agricultural Forestry University: 6. [王海为, 2003. 锥栗的品种与群落学研究 [D]. 福州:福建农林大学:6.]
YANG QH, YANG W, LI XR, 2001. Affecting factors of tropical plant seed germination [J]. Seed, 5: 45-48. [杨期和, 杨威, 李秀荣, 2001. 热带植物种子萌发影响因素初探 [J]. 种子, 5:45-48.]
YANG QH, YE WH, ZHANG Y, et al., 2005. Eco-physiological characteristics of germination and storage of Castanopsis chinensis seeds [J]. J Beijing For Univ, 27(1): 92-95. [杨期和, 叶万辉, 张云, 等, 2005. 锥栗种子萌发和贮藏特性的初步研究 [J]. 北京林业大学学报, 27(1):92-95.]
ZHANG XQ, QI J, CAO WX, 2014. Effects of salt, temperature and their interaction on seed germination and seedling growth of Elymus nutans [J]. Chin J Grassl, 36(1): 24-30. [张小娇, 祁娟, 曹文侠, 2014. 盐分、温度及其互作对垂穗披碱草种子萌发及幼苗生长的影响 [J]. 中国草地学报, 36(1):24-30.]
ZHENG CL,2008. Studies on several aspects on Chinquapin (Castanea henryi Rehd. & Wils.)s biology [D]. Fuzhou: Fujian Agricultural Forestry University: 1-3. [郑诚乐, 2008. 锥栗若干生物学问题研究 [D]. 福州: 福建农林大学:1-3.]
ZHENG CL, WU SH, SHE WQ, et al., 2003. Analyses of nutrient ingredients and quanlity evalution in nuts of Chinquapin (Castanea henryi) [J]. J Fujian Coll For, 23(4): 293-296. [郑诚乐, 吴少华, 佘文琴, 等, 2003. 锥栗果实营养成分分析与品质的模糊评判 [J]. 福建林学院学报, 23(4):293-296.]
(责任编辑 蒋巧媛)
猜你喜欢
儿童时代·幸福宝宝(2019年9期)2019-10-28
莫愁·家教与成才(2017年7期)2017-07-11
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
中国市场(2016年36期)2016-10-19
河北渔业(2015年10期)2015-10-15
湖北农业科学(2015年17期)2015-10-09
